| Скачать .docx | Скачать .pdf |
Реферат: Доисторический человек кембриджский путеводитель


|
|
Scaned and Created by BOBER
David Lambert
and the Diagram Group
Denis Kennedy
Annabel Else
David Harding
Arthur Lockwood
Richard Hummerstone
Graham Rosewarne, and
Joe Bonello, Ray Burrows, Richard Czapnik,
Brian Hewson, Philip Patenall
CAMBRIDGE UNIVERSITY PRESS
Cambridge
London New York New Rochelle Melbourne Sydney
ПЕРЕВОД С АНГЛИЙСКОГО
В. 3. МАХЛИНА
ПОД РЕДАКЦИЕЙ
А. Н. ОЛЕЙНИКОВА
ББК 26.323
Л 21
УДК 569.9-03.20
Л 21 Доисторический человек: Кембриджский путеводитель:
Пер. с англ.— Л.: Недра, 1991.— 256 с: ил.—
Пер. изд.: Англия, 1987.
ISBN 5—247—01726—9 (рус.)
ISBN 0—521—33364—4 (англ.)
ISBN 0—521—33644—9 (англ.)
В превосходно иллюстрированной книге общедоступно развернуто родословное древо
человека и его ближайших родичей: от предков приматов до высших обезьян и первых людей.
Показано, как в процессе эволюции возникли и трансформировались основные органы и физиоло-
гические системы человеческого организма. Приведены современные палеоантропологические
и археологические методы исследования и анализа доисторических антропоидов по их иско-
паемым остаткам. Иллюстрации органически связаны с текстом и являются основным содержа-
нием книги.
Для широкого круга читателей — от старшеклассников до ученых, музейных работников,
популяризаторов науки.
л 1904000000-313 52 91 ББК 26.323
043(01)—91
ISBN 5—247—01726—9 (рус.)
ISBN 0—521—33364—4 (англ.)
ISBN 0—521—33644—9 (англ.)
© Diagram
Visual Information
Ltd, 1987
© Перевод на русский язык.
В. 3. Махлин, 1991
© От редактора перевода.
А. Н. Олейников, 1991
This book was originally published in the English language by Diagram Visual Information Ltd of
195 Kentish Town Road London NW5 8SY, England.
Глава 1
18 Уникальное животное-
20 Древнейшие предки человека
22 Свидетельства еравншельной анатомии
24 Коеги и мышцы
26 Череп, челюсти и зубы
28 Конечности
30 Кожа
32 Нервы и оршны чувств
34 Мозг
36 Т ри системы жи а к-обест течения
38 Размножение
 40 Эмбриональное развитие человека
40 Эмбриональное развитие человека
42 Изъяны нашею орншизма
44 Биомолекулярные связи
46 Человек-примат
Глава 2
50 Как появились приматы
52 Полуобезьяны
54 Мир, в котором они жили
56 Первые приматы
58 /Утаписы
60 Лемуры
62 До.тгопягы
Глава 3
66 Об антропои тах
68 Животный мир и климат миоцена
70 Обезьяны Нового Света
72 Антропоиды Старого Света
74 Обезьяны Старою Света (1)
76 Обезьяны Старою Света (2)
78 Нтминоиты
80 Дриопитек и труте
82 Древние пошины
Глава 4
Человекообразные
обезьяны и человек
86 Крупные человекообразные обезьяны
88 Люди, обезьяны и молекулы
90 Сравнительная анатомия человека
 и человекообразных обезьян
и человекообразных обезьян
92 Человек — родич обезьяны
94 Образ жизни человекообразных
Глава 5
«Человекообезьяны»
и ранние люди
98 Становление человека
100 Человеческая триба
102 Австралопитек из Афара
104 Австралопитек африканский
106 Австралопитек могучий
108 Австралопитек Бойса
110 Человек умелый
112 Ранние следы материальной культуры
114 Первые стоянки
116 Животный мир древней Африки
Глава 6
120 Человек прямоходящий
122 Меняющийся мир
124 Прямоходящий человек в Африке
126 Прямоходящий человек в Европе
128 Прямоходящий человек в Азии
130 Рубила и чопперы
132 Охота
 134 Жилище и очаг
134 Жилище и очаг
Глава 7
138 Направления эволюции человека
140 Оледенения
142 О неандертальском человеке
144 Европейские неандертальцы
146 Неандертальцы за пределами Европы
148 Орудия неандертальцев
150 Охота
152 Петеры, шалаши и одежда
 154 Погребения, ритуалы, искусство
154 Погребения, ритуалы, искусство
Глава 8
158 О раннем современном человеке
160 Новые приемы обработки камня
и кости
162 Культура кроманьонцев
164 Способы охоты
166 Охотничья добыча
168 Жилище и одежда
170 Искусство и ритуалы
172 Кроманьонские погребения
174 Мезолит
 176 Орудия эпохи мезолита
176 Орудия эпохи мезолита
![]()
Глава 9
Современный человек
осваивает Землю
![]() 180 Происхождение современного человека
180 Происхождение современного человека
182 Современный человек в Африке
184
186
188 Человек и животный мир Америки
190 Первые австралийцы
 192 Климат, цвет кожи и телосложение
192 Климат, цвет кожи и телосложение
Глава 10
196 Потепление климата
198 Возникновение земледелия
200 Скотоводство
202 Орудия эпохи неолита
204 Дома, могилы и святилища
206 От камня к металлу
208 Появление городов
210 Орудия эпохи бронзы
212 Культура Северной Америки
214 Центральная и Южная Америка
216 Питание и телосложение
218 Рост численности населения
 220 Будущее человечества
220 Будущее человечества
Глава 11
224 Образование ископаемых остатков
226 Поиски ископаемых остатков
228 Научные раскопки
230 Реконструкция ископаемых остатков
232 Кости и предметы материальной
культуры
 234 Ключи к познанию окружающей среды
234 Ключи к познанию окружающей среды
236 Относительная датировка
238 Абсолютная хронометрия
240 Трудности, связанные с изучением
ископаемых остатков
242 В поисках древнего человека (1)
244 В поисках древнего человека (2)
246 Музейные экспозиции (1)
249 Музейные экспозиции (2)
252 Музейные экспозиции (3)
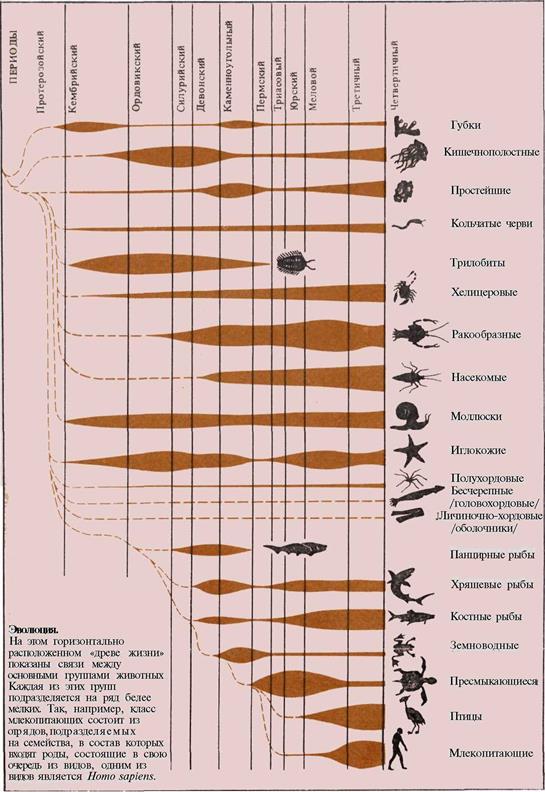
На протяжении последних десятилетий
наука о происхождении и развитии человека
обогатилась многими открытиями. Она взя-
ла на вооружение точные методы биологии,
физики и химии, тесно сплелась с геологи-
ческими дисциплинами, освоила ряд совер-
шенных технических приемов и вышла на
новые теоретические рубежи.
Книга Давида Ламберта «Доисториче-
ский человек», посвященная этой интерес-
ной области знания, была впервые опубли-
кована Издательством Кембриджского уни-
верситета в 1987 году.
Издательство Кембриджского универ-
ситета — одно из старейших в Западной
Европе. Надпись на его фирменной марке
гласит, что право «издавать и продавать
книги любого содержания» было пожало-
вано Университету королем Генрихом VIII
в 1534 году. Начиная с 1584 года Универси-
тет постоянно издает разнообразную лите-
ратуру, среди которой видное место зани-
мают естественнонаучные работы. Кроме
Кембриджа издательство имеет отделения
в Лондоне, Нью-Йорке, Нью-Рошелле,
Мельбурне и Сиднее. А издание, с которого
сделан перевод, было напечатано в Турине.
По замыслу издателей, книга Давида
Ламберта о доисторическом человеке приз-
вана дополнить написанный этим же авто-
ром и выпущенный двумя годами раньше
научно-популярный обзор «Доисторическая
жизнь» (Кембридж, 1985), повествующий
о происхождении и эволюции органического
мира — от простейших клеточных структур
до человека. Перевод книги «Доисториче-
ская жизнь» также будет издан Ленинград-
ским отделением издательства «Недра».
Емкий и занимательный рассказ Д. Лам-
берта о становлении человечества сопро-
вождается большим количеством превосход-
ных иллюстраций, выполненных коллекти-
вом художественной группы «Диаграмма».
Язык книги легок для восприятия, что
делает текст ее вполне доступным даже для
неподготовленного читателя. Вместе с тем
изложение материала ведется без каких-
либо упрощений, при полном соблюдении
научной строгости содержания. Автор не
избегает вопросов, не решенных наукой
окончательно, и в ряде случаев излагает
конкурирующие взгляды различных уче-
ных. В книге немало указаний на материалы,
полученные в нашей стране. Однако преоб-
ладающая часть сведений касается более
известных автору данных по зарубежным
территориям.
Ученые различных стран нередко следуют
традициям издавна сложившихся нацио-
нальных научных школ. . В определенной
мере такой подход можно проследить, читая
книгу Д. Ламберта. Так, например, в нашей
стране третичный период геологической ис-
тории, о котором идет речь в нескольких гла-
вах, уже давно подразделен на два периода:
палеогеновый, включающий палеоцен, эоцен
и олигоцен, и неогеновый, охватывающий
миоцен и плиоцен. А докембрийские отложе-
ния подразделяются на две акротемы: ар-
хейскую (в которую входят азойские и архе-
озойские образования) и протерозойскую.
Внимательный читатель, вероятно, обнару-
жит и некоторые другие подробности, не
вполне совпадающие с трактовкой фактов,
известной ему из отечественной литературы.
Однако следует помнить, что различные
аргументированные точки зрения имеют
равное право на существование. И посколь-
ку излагаемый автором материал всегда
сопровождается ссылками на научные ис-
точники, откуда подчерпнуты соответствую-
щие сведения, мы полностью сохранили из-
лагаемые автором концепции, дающие воз-
можность познакомиться с современными
воззрениями английских исследователей.
Существуют международные правила,
согласно которым в биологической и палеон-
тологической литературе названия живот-
ных и растений в обязательном порядке
пишутся по-латыни. Это позволяет избежать
ошибок, нередко возникающих вследствие
того, что на разных языках одно и то же
животное (или растение) может называться
по-разному или, что еще хуже, одно и то же
название может использоваться для обозна-
чения разных организмов. Для жителей
стран, пользующихся латинским шрифтом,
вкрапление в текст латинских терминов не
составляет затруднений при чтении. Сохра-
нив в переводе использованные автором
латинские наименования биологических
объектов имеет смысл напомнить несложные
правила их чтения. В отличие от английско-
го, немецкого и французского языков, боль-
шинство латинских букв и их сочетаний чи-
| таются так |
же, как |
и пишутся. |
|
| Латинские |
Следует |
Латинские |
Следует |
| буквы |
читать |
буквы |
читать |
| а |
a |
n |
н |
| b |
6 |
o |
о |
| с |
ц |
p |
п |
| d |
д |
q |
кв |
| е |
э |
r |
р |
| f |
ф |
s |
с |
| g |
г |
t |
т |
| h |
звук, по- |
u |
у |
| добный |
|||
| украин- |
|||
| скому г |
|||
| i |
и |
v |
в |
| j |
й |
w |
в |
| k |
к |
x |
кс |
| 1 |
ль |
y |
иили ы |
| m |
м |
z |
з |
| Кроме того, в тексте встретится несколь |
|||
| сочетаний букв, которые читаются так: |
|||
| ja, уа — я |
ju, уи — |
ю ph — |
ф |
| je, уе — е |
| ае — э |
| rh |
| р т |
Буква «s» между двумя гласными читает-
ся как «з».
Без труда запомнив эти основные прави-
ла, можно совершить первый шаг к свобод-
ному чтению биологической и палеонтоло-
гической литературы.
Александр Олейников
Эта книга — сжатое изложение истории происхождения человечества.
Популярная форма подачи материала сочетается в ней со строгой научной
терминологией. Многочисленные реалистические изображения, реконструк-
ции палеобиологических объектов, диаграммы и родословные древа снаб-
жены надлежащими пояснениями. В итоге удалось создать полезное посо-
бие, доступное каждому — от любознательного школьника до начинающего
ученого.
В книге 11 глав. Каждой из них предпослано краткое введение, после
которого следуют разделы, посвященные конкретным вопросам, отраженным
в их заглавиях.
Глава 1. Что такое человек? Здесь рассматриваются основные признаки,
которые отличают нас от остальных живых существ. Показано, как развива-
лись в ходе эволюции органы и физиологические системы человеческого
организма.
Глава 2. Примитивные приматы. В этой главе идет речь о происхождении
приматов — группы млекопитающих, в которую входит и человек. Описы-
ваются также низшие приматы, вымершие и современные полуобезьяны.
Глава 3. Эволюция антропоидов. Здесь прослежены основные тенденции
эволюции высших приматов — группы, к которой относятся человекооб-
разные обезьяны и человек. Глава заканчивается рассказом о доисториче-
ских человекообразных обезьянах, которые, вероятно, были предками совре-
менных обезьян и человека.
Глава 4. Человекообразные обезьяны и человек. Рассматриваются мно-
гочисленные признаки сходства, по которым можно судить о тесном родстве
между человеком и современными человекообразными обезьянами.
|
|

Глава 5. «Человекообезьяны» и ранние люди. Освещены основные про
цессы эволюции, в результате которых доисторические человекообразные
обезьяны превратились в первобытного человека. Дано описание австрало-
питеков и их возможного потомка Homo habilis (человек умелый) — первого
известного представителя нашего рода.
Глава 6. Человек прямоходящий. Охарактеризованы внешний облик
и образ жизни широко распространенного в прошлом вида человека — Homo
erectus (человек прямоходящий), который, очевидно, был нашим предком.
Глава 7. Неандертальский человек. В этой главе идет речь о древней-
ших представителях вида Homo sapiens (человек разумный); к этому виду
относится современный человек.
Глава 8. Современный человек в Европе. Здесь рассказано о древней-
ших европейских представителях наиболее совершенного подвида современ-
ного человека — Homo sapiens sapiens.
Глава 9. Современный человек осваивает Землю. Как явствует из загла-
вия, речь идет о расселении современного человека по земному шару.
Глава 10. После ледникового периода. В этой главе рассказано, как
преобразилась жизнь человека в ходе культурной эволюции за последние
10 тысяч лет.
Глава 11. Познание истории человечества. Речь идет о труде ученых,
исследующих наше доисторическое прошлое, о достижениях крупнейших
палеонтологов и археологов, о музеях, где представлены интересные экспо-
зиции по истории человечества.
В заключение приведен перечень книг для самостоятельного чтения.
В книге «Доисторический человек» внесены незначительные изменения
в классификацию приматов, предложенную в 1979 году Ф. Салаем и Е. Дел-
|
|
соном; принят проведенный в 1982 году учеными Кембриджского универ-
ситета пересмотр датировки ряда подразделений шкалы геологического вре-
мени; отмечены спорные вопросы, относительно которых у специалистов
имеются различные точки зрения.
Автор несет полную ответственность за все сведения, изложенные в этой
книге, и благодарит всех специалистов, содействовавших подготовке настоя-
щего издания.
Выдержать в этой книге соответствие требованиям совре-
менных воззрений автору помогли консультанты:
Доктор Л. Айелло [Dr Leslie Aielio, University College,
University of London];
Доктор П. Эндрюс [Dr Peter Andrews, British Museum
(Natural History), London];
Доктор П. Л. Древетт [Dr Peter L. Drewett, Institute of
Archaeology, University of London];
Доктор X. Штрингер [Dr Christopher Stringer, British
Museum (Natural History), London];
Профессор Б. Вуд [Professor Bernard Wood, The Univer-
sity of Liverpool, England].
При подготовке книги к печати были использованы мно-
гие справочные издания, большая часть которых перечислена
ниже. Выражаю благодарность авторам и издателям этих
работ и приношу извинения за возможную неполноту при-
веденного списка.
Aielio L. Discovering the Origins of Mankind. Longman,
1982.
Archer M. and Clayton G. (editors). Vertebrate Zoogeo-
graphy and Evolution in Australasia. Hesperian Press. Austra-
lia, 1984.
Bordaz J. Tools of the Old and New Stone Age. David and
Charles, 1971.
Brace C. E. and Montagu A. Human Evolution. Macmil-
lan, 1977.
Clapham F. M. (editor). The Rise of Man. Sampson Low,
1976.
Cole S. The Neolithic Revolution. British Museum (Na-
tural History), 1970.
Coles J. M. and Higgs E. S. The Archaeology of Early Man.
Faber and Faber, 1969.
Day M. H. Fossil Man. Hamlyn, 1969.
Gribbin J. and Chef as J. The Monkey Puzzle. Bodley Head,
1982.
Jurmain R., Nelson H., Kurashina H. and Turnbaugh W. A.
Understanding Physical Anthropology and Archeology. West
Publishing Co., 1981.
Kennedy G. E. Paleoanthropology. McGraw Hill, 1980.
Leakey M. Olduvai Gorge. Collins, 1979.
Leakey R. E. The Making of Mankind. Michael Joseph,
1981.
Lewin R. Human Evolution. Blackwell Scientific Publi-
cations, 1984.
McEvedy C. and Jones R. Atlas of World Population Histo-
ry. Allen Lane, 1978.
Napier P. Monkeys and Apes. Hamlyn, 1970.
Oakley K. P. Man the Tool-Maker. British Museum (Natu-
ral History), 1972.
Palmer S. Mesolithic Cultures of Britain. Dolphin Press,
1977.
Phillips P. The Prehistory of Europe. Penguin Books, 1981.
Poirier F. E. Fossil Evidence. C. V. Mosby Co., 1981.
Reader J. Missing Links. Collins, 1981.
Romer A. S. Man and the Vertbrates. Penguin Books, 1954.
Romer A. S. The Vertebrate Body. W. B. Saunders Co.,
1970.
Semenov S. A. Prehistoric Technology. Adams and Dart,
1964.
Shackley M. Neanderthal Man. Duckworth, 1980.
Szalay F. S. and Delson, E. Evolutionary History of the
Primates. Academic Press, 1979.
Tomkins S. The Origins of Mankind. Cambridge Univer-
sity Press, 1984.
Weiner J. S. Man's Natural History. Weidenfeld and Nicol-
son, 1971.
Wood B. The Evolution of Early Man. Peter Lowe, 1976.
Кроме того, ряд сведений почерпнут из серии публикаций
The Emergence of Man (Time-Life Books, 1972—1973), из
Британской энциклопедии и журнала National Geographic.
 В этой главе дается общий
В этой главе дается общий
обзор физиологических и
психических особенностей,
делающих вид
Homo sapiens
(гомо сапиенс
— человек ра-
зумный) уникальным.
Однако, являясь результа-
том эволюционного про-
цесса, наше тело унаследо-
вало ряд систем от других
организмов. Сравнение с
различными современными и
вымершими позвоночными
позволяет выявить основные
изменения, которые привели
к формированию специфи-
ческих особенностей в строе-
нии костей, легких, системы
кровообращения и других ор-
ганах человека.
В конце главы определяется
место человека в царстве
животных как члена отряда
приматов класса млекопи-
тающих.
На рисунке видны 46 хромосом
в клетке человеческого тела
во время ее деления (увеличено
в 5600 раз) Гены человеческих
хромосом являются носителями
той наследственной информации,
которая делает наш вид
отличным от всех прочих.
(По фотоснимку из журнала
«Сайентифик Америкен»).
|
|

![]() 2 Зак. 5116
2 Зак. 5116
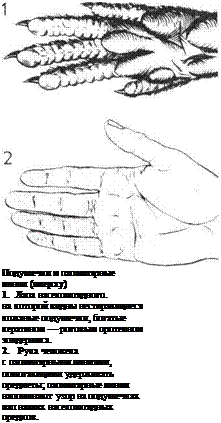
Особенности человеческого
организма.
На этих четырех рисунках
изображены характерные
особенности, отличающие нас
от прочих живых существ.
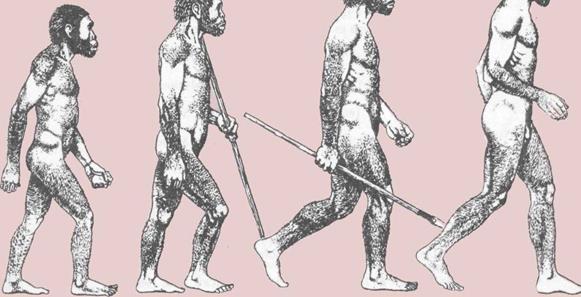
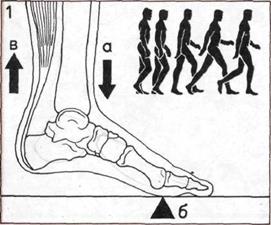
1. Ходьба на двух ногах.
Вес тела переносится с пятки
через наружный край ступни
на подушечки и большой палец.
Нога и ступня действуют
подобно рычагу.
а — Нагрузка передается
через большеберцовую кость.
б — Точка опоры приходится
на носок.
в — Усилие создается
ахиллесовым сухожилием,
которое, сокращая мышцы икры,
поднимает пятку.
2. Разнообразие функций рук.
а — Силовой захват.
б — Точный захват.
в — Ладони, образующие чашу.
Виду Homo sapiens давали самые различные опре-
деления: разумное, политическое, пользующееся
орудиями, общественное и обладающее самосоз-
нанием существо. Вооруженные высокоразвитой тех-
никой и организованные в крупные социальные груп-
пы, мы в настоящее время используем растения и
животных, преобразуем поверхность Земли для удов-
летворения наших потребностей в пище, топливе,
жилье и транспорте. Для защиты от себе подобных
мы сейчас обладаем также оружием, способным пол-
ностью стереть с лица нашей планеты большинство
видов живых организмов.
И тем не менее, человек разумный (Homo sa-
piens) — это самое могучее из всех живых существ—
состоит из тех же химических элементов: углерода,
водорода, кислорода и азота, что и любые прочие
млекопитающие. Мы отличаемся лишь тем, как эти
элементы сгруппированы в химические соединения,
клетки, ткани и системы органов, которые образуют
организм, обладающий всесторонним сочетанием
разнообразных способностей, не встречающихся
более ни у одного другого живого существа.
Скачущая лошадь обгонит бегущего человека.
У орла более острое зрение, чем у нас. Нашим зубам
и ногтям не сравниться с клыками и когтями леопар-
да. И все же мы обладаем четырьмя решающими осо-
бенностями, сочетание которых присуще только на-
шему виду; это прямостоящий скелет, подвижных
руки, способные манипулировать предметами, трех-
мерное цветовое зрение и уникальный по своей слож-
ности мозг. Совокупность этих четырех свойств дает
людям преимущества, превращающие их в хозяев
планеты.
|
|

Прямостоящий скелет позволяет нам передвигать-
ся, в отличие от других животных, на двух ногах, пе-
ренося вес с пятки на пальцы ноги, что превращает
каждый шаг в упражнение по балансированию, тре-
бующее мгновенной координации работы мускулов
спины, бедер и ног. Мы можем не только ходить, но
также бегать, прыгать, плавать, нырять и взбираться
на скалы и деревья. Бегуны на длинные дистанции
своей выносливостью превосходят оленя.
Не испытывая потребности опираться на передние
конечности, мы пользуемся гибкими и чуткими паль-
цами рук, чтобы исследовать поверхность пред-
метов на ощупь, а также сжимать их с необходимой
силой и точностью. Пользуясь по своему желанию тем
или иным орудием попеременно, мы можем значи-
тельно эффективнее воздействовать на окружающую
нас среду, чем любые другие млекопитающие, перед-
ние конечности которых снабжены копытами или
вооружены когтями.
Смотрящие вперед и чувствительные к цвету глаза
позволяют нам отчетливо фокусировать изображе-
ния, точно определять расстояние и различать не
только цвет, но также форму и яркость освещения
предметов. Эти способности имеются у немногих
других млекопитающих. Мы можем следить за пере-
мещением предметов, не поворачивая головы, при
помощи одного лишь движения глаз. А то, что мы
стоим прямо, возвышаясь над уровнем земли, позво-
ляет нам видеть значительно дальше, чем видят дру-
гие наземные млекопитающие таких же размеров.
И наконец, мозг, очень крупный относительно раз-
меров тела, обладает великолепными способностями
к обучению, логическому мышлению, управлению
речью и к точной координации зрения и движений
рук.
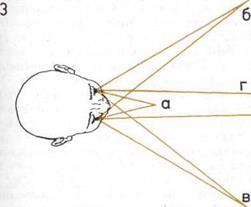
3. Бинокулярное
(стереоскопическое) зрение.
Оба глаза могут быть
сфокусированы на объектах,
расположенных в различных
направлениях и удаленных
на разные расстояния:
а — Близко.
б — Слева.
в — Справа.
г — Далеко.
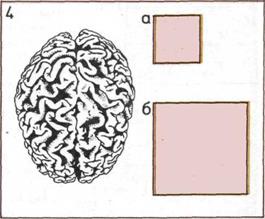
4. Большой мозг.
Если рассматривать
человеческий мозг сверху,
видно, что он изборожден
глубокими извилинами. Площадь
их поверхности очень велика,
от этого и зависят в значительной
степени умственные
способности человека.
а — Видимая площадь
поверхности мозга.
б — Площадь поверхности
мозга с «расправленными»
извилинами (2090 см2
).
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
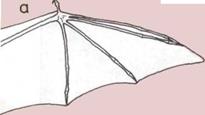
Аналогичные структуры.
а — Крыло летучей мыши,
позвоночного Это крыло состоит
из кожаных перепонок,
натянутых между костями
передних конечностей,
пальцами и задними
конечностями.
б — Крылья бабочки,
беспозвоночного. Каждое
крыло — это перепонка,
растянутая на жестких
трубчатых прожилках и
покрытая чешуйками,
которые перекрывают друг
друга.
Гомологичные структуры.
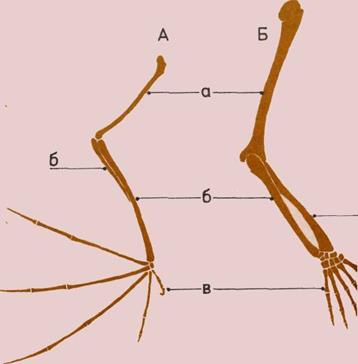
 А — Кости крыла летучей мыши.
А — Кости крыла летучей мыши.
Б— Кости руки и кисти человека.
Крыло и рука могут казаться
непохожими, по они имеют общие
структурные особенности,
унаследованные от общего
позвоночного предка.
а — Плечевая кость.
б — Лучевая и локтевая кости.
в — Пять пальцев.
В сходстве строения тела человека и некоторых
современных и вымерших животных лежит ключ к
решению вопроса об их общих предках: чем больше
сходных признаков, тем ближе родство. Но анатомы
проводят тщательное различие между гомологич-
ными органами тела, сходство которых вызвано об-
вдим происхождением (например, рука человека и
крыло птицы), и аналогичными органами, которые
хотя и кажутся похожими, однако не связаны един-
ством происхождения (например, крылья мух
и птиц).
Современные примитивные морские организмы,
очень похожие на живших 700 миллионов лет назад,
свидетельствуют о значительной древности основных
характерных черт нашего тела: его внутреннего стро-
ения и внешнего облика, его передней и задней сто-
рон, его правого и .левого боков.
![]()
 У морской актинии только два слоя клеток: один -
У морской актинии только два слоя клеток: один -
наружный - служит ей зашитой и передает инфор-
мацию о внешнем мире, а другой выстилает ее внут-
реннюю полость и обеспечивает питание и размно-
жение. Через единственное отверстие поступает
пища, которую переваривают клетки, выстилающие
внутренний мешочек, и через то же отверстие выде-
ляются отходы жизнедеятельности тела и отлага-
ются яйца. У червей этот мешочек превращается в
трубку, имеющую рот с одного конца и задний про-
ход с другого, что. в основном соответствует стро-
ению и нашей системы пищеварения.
Такое расположение органов автоматически при-
водило к тому, что у тела можно было различить
переднюю и заднюю части. Специальные клетки,
сгруппированные в виде мышц, помогали организ-
мам двигаться вперед в поисках пищи для рта. Вок-
руг рта появлялись пучки щупалец или зубы, слу-
жащие для захвата пищи. Клетки, которые специа-
лизировались в поиске пищи по запаху или внеш-
нему виду, также сосредотачивались в передней ча-
сти тела, где формировалась голова с мозгом, коор-
динирующим сигналы нервной системы, сложность
которой все возрастала.
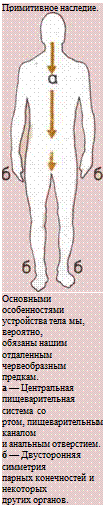 Тем временем тело ранних червеобразных предков
Тем временем тело ранних червеобразных предков
человека приобрело двустороннюю симметрию: каж-
дая сторона представляла собой зеркальное отраже-
ние другой. Это облегчало продвижение вперед, пово-
рот в ту или иную сторону и сохранение правильного
курса движения. Парность наших конечностей, глаз,
ушей и ноздрей является следствием такого строе-
ния тела.
|
|
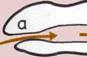
Передняя и задняя части тела.
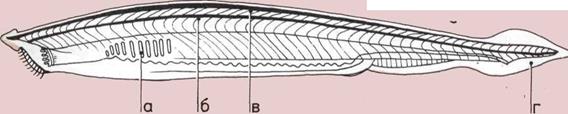 На этом продольном разрезе
На этом продольном разрезе
червя показано направленное
прохождение пищи,
обусловленное развитием трех
функциональных структур.
а — Рот.
б — Пищеварительный канал.
в — Анальное отверстие.
Amphioxus, современный
ланцетник.
У него нет ни головы,
ни челюстей, ни позвонков,
но подобные существа, вероятно,
были предками всех
позвоночных — от рыб до
человека.
Как и у рыб, у ланцетника
есть:
а — Жабры.
б — Нервный тяж.
в — Homox орда
(предшественник позвоночника).
г — Хвостовой плавник.
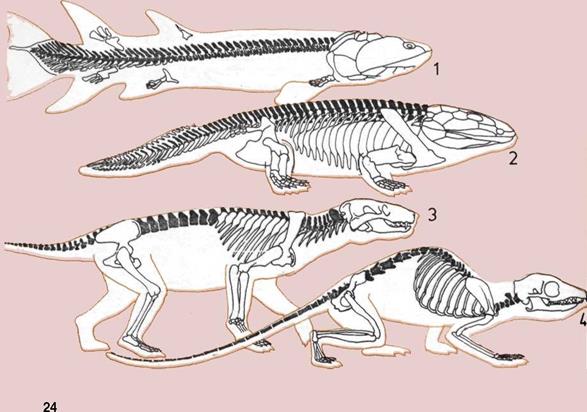
Развитие позвоночника.
На шести рисунках
прослеживается эволюция
позвоночника от горизонтального
стержня до вертикального
столба.
1.
Прямой позвоночник рыбы
эустеноптерон
(Eusthenopieron) дал выигрыш
в силе ее плавательным мышцам.
Эта рыба жила 375 миллионов
лет назад.
2.
Сильный изогнутый
позвоночник поддерживал
ранних ползающих
земноводных, таких как
ихтиостега (Ichthyostega),
при их передвижении по суше.
3.
Сложные изменения
позвоночного столба и ребер
улучшили способность
к передвижению по суше
древнего пресмыкающегося
тринаксодона (Thrinaxodon),
похожего на млекопитающих,
предком которых он, возможно,
являлся.
Человеческое тело состоит в основном из костей
и мышц, развившихся из среднего слоя клеток, ко-
торый впервые образовался между двумя другими
слоями у некоторых ранних беспозвоночных. У мно-
гих из них появилась наружная раковина или скелет
для защиты и опоры, а у позвоночных развился внут-
ренний костяной скелет. Скопления кальция и фос-
фора служили двойной цели: как защита жизненно
важных органов и как место прикрепления мышц,
что создавало точку опоры для конечностей или плав-
ников. Краб вынужден по мере роста сбрасывать
время от времени свой панцирь, лишаясь защиты
до тех пор, пока не затвердеет новая броня. Внут-
ренний же скелет растет одновременно со всем те-
лом.
Основной частью скелета бесчелюстных рыб и всех
позднейших позвоночных является спинной хребет
(позвоночник), состоящий из соединенных друг с
другом позвонков. Они укрывают легко уязвимый
спинной мозг. Ребра, расположенные через опреде-
ленные промежутки по обеим сторонам спинного
хребта, защищают мягкие внутренние органы и слу-
|
|
жат местом прикрепления находящихся между ними
мышечных волокон. С помощью мышц рыба пово-
рачивает свое тело, отталкивается хвостом от воды
и продвигается вперед.
У рыб почти прямой позвоночник и сравнительно
слабые позвонки. Позвоночные, передвигающиеся
по суше, нуждаются в более сильной внутренней
опоре. Ранние земноводные, такие как Ichtyostega,
имели прочный, изогнутый в виде плавной дуги поз-
воночник, к которому снизу было подвешено тело;
большая грудная клетка поддерживала тело, когда
оно лежало на земле. Кроме того, ребра, вероятно,
раздвигались и сдвигались, помогая работе легких.
У некоторых наземных позвоночных спинной хре-
бет претерпел сложные изменения. По мере того как
конечности становились органами передвижения,
хвост уменьшался и превращался просто в балан-
сир или метелку, отгоняющую мух. У человекообраз-
ных обезьян и человека хвост исчез совсем, ребра
в области шеи и поясницы постепенно сошли на нет.
У пресмыкающихся, бывших предками млекопита-
ющих, уменьшился размер шейных ребер, что позво-
лило им более свободно двигать головой, а шейные
позвонки выпрямились и подняли голову вверх. У не-
которых млекопитающих в результате дальнейшего
изменения двух первых шейных позвонков голова
стала еще более подвижной. У человека обозначился
второй изгиб позвоночника в нижней части спины,
который приподнял грудь и подал ее несколько на-
зад. Грудь и голова оказались выше бедер, что обес-
печило телу хорошее равновесие при стоянии на двух
ногах.
|
|
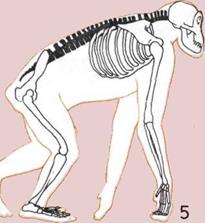
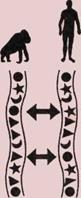
4.
Гибкий позвоночник ранних
насекомоядных млекопитающих,
похожих на эту современную
полуобезьяну тупайя, позволял
им карабкаться по деревьям.
5.
У почти прямоходящих
шимпанзе позвоночник более
прямой, чем у их четвероногих
обезьяноподобных предков,
имевших дугообразно изогнутый
хребет.
6.
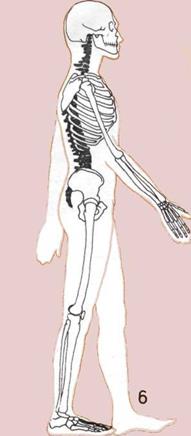 Прочный и подвижный
Прочный и подвижный
позвоночный столб человека
имеет изгибы, позволяющие
удерживать голову и тело над
центром тяжести.
Наши череп, челюсти и зубы имеют разное и слож-
ное происхождение.
У ранних позвоночных, несколько напоминавших
ныне живущих бесчелюстных рыб — миног, в сущно-
сти, не было ни черепа, ни челюстей. В их кожном
покрове находились хрящевые или костные пластин-
ки, которые прикрывали голову и шею, защищали
мозг, глаза и внутреннее ухо и придавали прочность
жаберным щелям, располагавшимся по бокам го-
ловы.
 У некоторых рыб часть жаберных пластинок исчез-
У некоторых рыб часть жаберных пластинок исчез-
ла, а одна пара увеличилась и образовала подвижные
челюсти, из бугорков которых со временем развились
зубы -- острые покрытые эмалью отростки, укреп-
ленные (как и сейчас у акул) в коже. Со временем
некоторые защитные пластинки, находившиеся в ко-
же, слились воедино с черепной коробкой, прикрыв-
шей большую часть головы, и, соединившись с че-
люстями, превратились в прочный костяной череп.
При этом в нижней челюсти образовались дополни-
тельные кости.
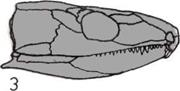
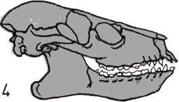
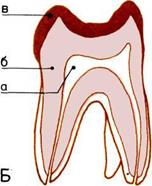
Развитие черепа (слева).
На пяти черепах (четыре из
которых принадлежат
ископаемым животным)
показана эволюционная
тенденция к уменьшению числа
костей.
1. Кистеперая рыба.
2. Примитивное земноводное.
3.
Примитивное
пресмыкающееся.
4.
Ископаемый лемур
(млекопитающее).
5.
 Современный человек.
Современный человек.
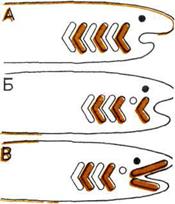
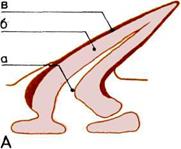
Развитие челюстей.
На трех рисунках показано,
как жаберные перегородки рыбы
превращаются в челюсти.
А — Бесчелюстная рыба.
Костные жаберные перегородки
(окрашены) перемежаются
с жабрами.
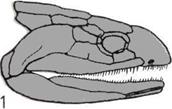
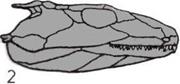 Б — Первая пара жабр,
Б — Первая пара жабр,
уменьшаясь, превращается
в брызгальце — крохотное
отверстие для всасывания
очищенной от ила воды.
В — Первая пара жаберных
перегородок смещается,
образуя подвижные челюсти.
|
|||||
|
|||||
|
|||||
|
|||||
|
|
Развитие конечностей.
На шести рисунках
прослеживаются изменения
костей таза и грудной клетки
(бедер и плеч), которые дали
конечностям выигрыш
в прочности и силе.
1.
Таз рыбы Eusthenopteron
не был прикреплен к спинному
хребту, а плечевой пояс этой
рипистидии был соединен с почти
неподвижным черепом.
2.
Тазовый пояс, соединяющий
задние конечности
с позвоночником, и плечевой
пояс, отделенный от черепа,
давали больший выигрыш
в силе и большую свободу
движений ранним четвероногим
животным, подобным этому
земноводному Ich thyostega.
3.
Дальнейшие изменения
сделали конечности этого
звероподобного
пресмыкающегося —
тринаксодона — еще более
сильными и подвижными.
4.
У полуобезьяны тупайя
длинный узкий таз и подвижное
плечевое соединение, что
позволяет ей поднимать
передние лапы и лазать
по деревьям.
Руки и ноги представляют собой отростки тела,
историю которых можно проследить по ископаемым
остаткам, начиная с массивных плавников рипидис-
тий (Rhipidistia) — кистеперых рыб, которые, воз-
можно, являются предками всех наземных позво-
ночных.
Эти плавники служили главным образом в каче-
стве балансиров, но были достаточно сильными, что
позволяло их обладателям, которые дышали воз-
духом, переползать короткие расстояния по суше.
Передние плавники (предшественники рук) были
присоединены к плечевому поясу, прикрепленному
в свою очередь к черепу. Задние плавники (пред-
шественники бедер и ног) прикреплялись к костям
таза внутри тела.
У ранних земноводных из плавников развились
членистые конечности. Колени и локти выдавались
вбок по сторонам тела. Задние конечности были
прикреплены к тазовому поясу, усиленному связкой
со спинным хребтом, что облегчало продвижение впе-
ред. Одновременно с этим плечевой пояс отсоеди-
|
|

пился от черепа, и теперь голова и передние конеч-
ности получили возможность двигаться независи-
мо друг от друга. Почти каждая из костей конечно-
стей этих древних доисторических животных имеет
свой аналог в скелете человека. Изменения связаны
в основном не с числом костей, а с соотношением их
размеров.
У высокоразвитых пресмыкающихся, похожих на
млекопитающих и являющихся далекими потомка-
ми земноводных, развились конечности с локтями и
коленями, находящимися с нижней стороны тела,
что позволило делать более широкие шаги и быстрее
передвигаться по суше. Каждая из конечностей за-
канчивалась пятью вытянутыми вперед когтистыми
пальцами; прежний массивный плечевой пояс стал
легче, что увеличило подвижность передних ко-
нечностей.
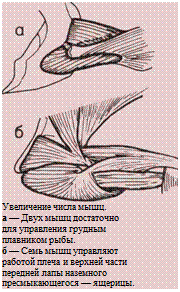
 Такое же строение тела сохранилось и у ранних
Такое же строение тела сохранилось и у ранних
млекопитающих. Дальнейшее развитие тела, позво-
лившее приматам в конце концов перейти к ходьбе
на двух ногах и освободившее их руки, будет рас-
смотрено в последующих главах.
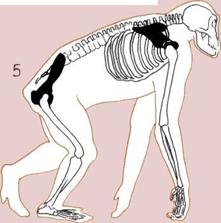
5.
Таз и плечевой пояс
человекообразной обезьяны
позволяют ей ходить и лазать,
используя все четыре конечности,
а также висеть на ветках,
держась за них руками.
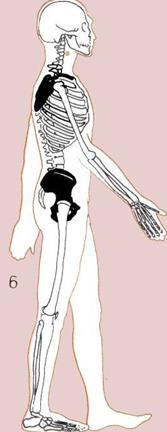
6. Короткий широкий таз
 и широкие подвижные лопатки
и широкие подвижные лопатки
человека позволяют ему
ходить на двух ногах
и свободно вращать руками.
|
|
Наша кожа, как и нервная система, вероятно, об-
разовалась из наружного слоя двуслойных тел наших
беспозвоночных предков. У нас, как и у рыб, имею-
щих тех же предков, кожа защищает мягкие, влаж-
ные внутренности от опасностей внешнего мира. Но
остатки костяного панциря ранних рыб у нас либо
исчезли, либо погрузились в тело, образовав клю-
чицы и значительную часть черепа.
 Наша кожа водонепроницаема, как и у рептилий,
Наша кожа водонепроницаема, как и у рептилий,
но от роговой чешуи, которой были покрыты тела
наших пресмыкающихся предков, у нас остались
только ногти — уплощенные щитки на пальцах
рук и ног, развившиеся из когтей. Способствую-
щие цепкости гребешки на пальцах рук и ног (папил-
лярные линии) остались у нас от жестких подуше-
чек, помогавших нашим предкам ходить и лазать.
Строение кожи.
Поперечные разрезы кожи
рыбы, пресмыкающегося
и человека показывают
сходство их строения: толстая
волокнистая соединительная
ткань — дерма — располагается
под более тонким поверхностным
слоем — эпидермисом. Наличие
этих двух слоев предполагает
общее происхождение кожи
всех позвоночных.
1.
Разрез кожи рыбы.
а — Эпидермис.
б — Чешуя.
в — Дерма.
2.
Разрез кожи
пресмыкающегося.
а — Роговая чешуя.
6 — Эпидермис.
в — Дерма.
3.
Разрез кожи человека.
а — Эпидермис.
б — Дерма.
в — Волосы.
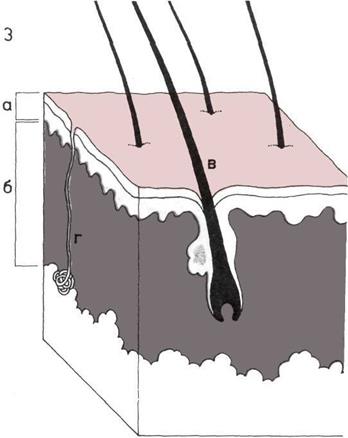 г — Канал потовой железы.
г — Канал потовой железы.
Волосы, покрывающие тело человека, вероятно,
развились из чувствительных выростов на коже, ко-
торые впервые наблюдаются у теплокровных те-
риодонтов — пресмыкающихся, явившихся непосред-
ственными предками млекопитающих. Менее яс-
ным представляется происхождение кожных желез;
вероятно, они развились у ранних млекопитающих
из сальных желез, выделения которых поддержи -
вают в здоровом состоянии кожу и волосы. Между
прочим, у некоторых людей имеются добавочные
грудные сосцы, остатки тех рядов сосцов, которые,
очевидно, имелись па нижней стороне тела ранних
млекопитающих.
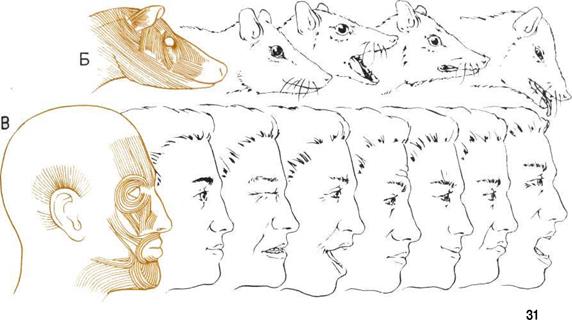
 Мускулы под кожей лица, развившиеся у наших
Мускулы под кожей лица, развившиеся у наших
предков-приматов, позволяют нам пользоваться ми-
микой - - языком выражений лица. Мимика отсут-
ствует у таких низших позвоночных, как рыбы,
которые, подобно маскам, сохраняют непроницае-
мое выражение.
|
|

| Развитие мимики. A.
Невыразительный взгляд А Б. Мускулатура и выражение B.
Высокоразвитая мускулатура |
|
|
Развитие нервной системы.
Два беспозвоночных
(изображены с сильным
увеличением) представляют
основные стадии эволюции
нервной системы.
А — Гидра (Hydra),
пресноводная родственница
медуз и актиний (морских
анемонов), имеет нервную сеть
охватывающую все тело
систему нервных клеток.
Б — Планария (Planaria),
плоский червь, имеет
двустороннюю симметрию
и центральную нервную
систему.
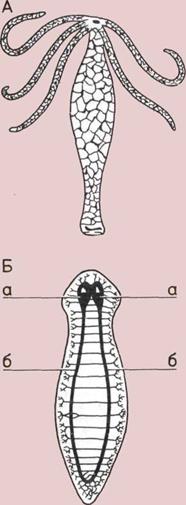 а — Мозг, расположенный
а — Мозг, расположенный
в головной части.
б — Нервные тяжи
(предшественники нашего
спинного мозга).
Все животные общаются и управляют своим телом
с помощью нервных клеток. Наша сложная нервная
система восходит к неупорядоченной нервной сети
низших существ, подобных медузам. У рыб появи-
лась внутренне взаимосвязанная нервная структура,
спинной мозг которой управляет рефлекторными
действиями, пересылает в головной мозг сигналы от
нервных окончаний, специализирующихся на вкусо-
вых, обонятельных и зрительных ощущениях. Нерв-
ная система также поддерживает равновесие и пе-
редает приказы головного мозга отдельным группам
мышц.
У нас, как и у ранних позвоночных, основные
органы чувств сосредоточены в голове — бывшем
переднем конце тела. У человека, как и у рыб, боль-
шинство этих органов получают ощущения от ве-
ществ, растворенных в воде, или от вибраций, про-
ходящих по жидкости. Так, вкусовые сосочки языка
человека различают вещества, растворенные в воде.
А носовые мембраны должны быть влажными, чтобы
чувствовать запахи. Поскольку носовые мембраны
связаны и со ртом, запахи являются, на самом деле,
сочетанием вкусовых и обонятельных ощущений.
Как и у рыб, жидкость, заключенная в закрытых
трубках внутреннего уха человека, обеспечивает ему
ощущение равновесия. Но изменения в ходе эво-
люции -- сначала у земноводных, затем у пресмы-
кающихся и наконец у млекопитающих — преобра-
зовали строение некоторых частей прежних рыбьих
жабр, снабдив уши наружными отверстиями и прев-
ратив их в эффективные органы слуха, которые вос-
принимают звуковые колебания, передаваемые по
воздуху. Внутри ушей звуки преобразуются в коле-
бания жидкости, которая омывает нервные окон-
чания в улитке - свернутой трубке внутреннего
уха.
Глаза возникли даже раньше, чем уши, образо-
вавшись из чувствительных к свету пятен, имевших-
ся у примитивных беспозвоночных. Уже у первый рыб
быгли парные глаза, очень похожие на наши и устро-
енные подобно фотокамере. Но слезные железы,
которые смачивают глаза, подверженные воздей-
ствию воздуха, появились только у земноводный,
а веки образовались еще позднее — у пресмыкаю-
щихся. Острое бинокулярное цветовое зрение, кото-
рым владеет человек, возникло у ведущих дневной
образ жизни предков человекообразный обезьян.
|
|
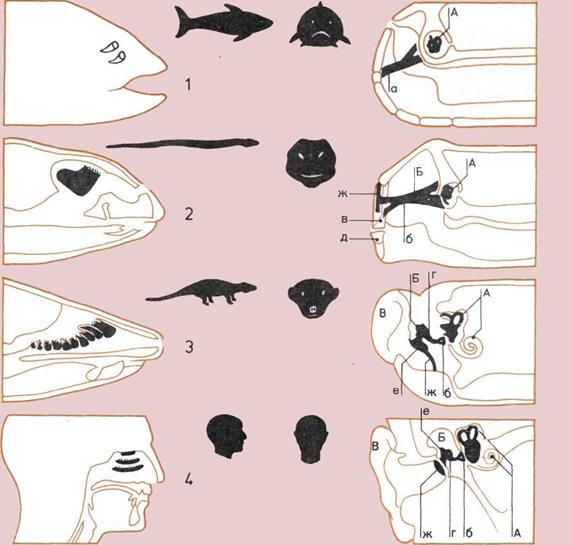
Чувство обоняния.
На продольном схематическом
разрезе четырех голов показано
развитие органов обоняния.
1.
Рыба; обонятельные
мембраны в полостях,
отделенных ото рта.
2.
Пресмыкающееся;
обонятельные мембраны
в носовой полости, связанной
со ртом.
3.
Типичное млекопитающее;
органы обоняния увеличились,
чю позволяет остро ощущать
запахи.
4.
Человек; органы обоняния
уменьшились, что
свидетельствует о большей
зависимости человека от зрения,
нежели от чувства обоняния.
Развитие органа слуха.
Поперечный разрез тех же
четырех голов показывает,
как перемещение косточек
помогло усиливать звуки,
приносимые колебаниями
воздуха, и передавать их во
внутреннее ухо.
А — Внутреннее ухо.
Б — Среднее ухо.
В — Наружное ухо.
а, б — Подъязычная кость (а)
превращается в стремечко
(слуховую кость) (б).
в, г — Квадратная челюстная
кость (в) превращается
в наковальню (г).
д, е — Артикулярная кость (д)
превращается в молоточек (е).
ж — Барабанная перепонка,
передающая звуковые волны
в среднее ухо.
![]() 3 Зак. 5116.
3 Зак. 5116.
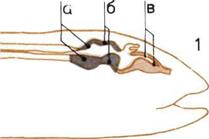
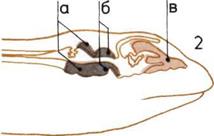
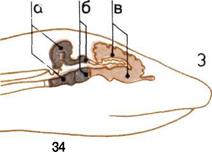
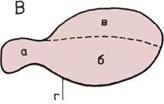
Развитие мозга.
По продольным разрезам
четырех голов прослеживаются
последовательные изменения
пропорций трех основных
отделов мозга.
1. Мозг рыбы.
2. Мозг пресмыкающегося.
3.
Мозг примитивного
млекопитающего
4. Мозг человека.
В каждом н из них мы видим:
а — Задний, или ромбовидный,
мозг (включая его ответвление -
мозжечок).
б — Средний мозг.
в — Передний (большой) мозг
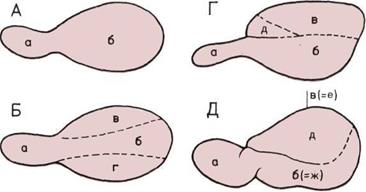
Слои мозга (вверху на с. 35).
Согласно одной из теорий, наш
мозг состоит из трех слоев,
которые отражают стадии его
эволюции.
а — Стадия пресмыкающихся;
задний и средний мозг —
рудимент прошлых эпох.
Человеческий мозг, который изобрел столько раз
нообразных вещей — от кремневых топоров камеи
ного века до атомных бомб, ведет начало от про
стого вздутия спинного мозга в его головной часто
около основных органов чувств. Головной мозг в
качестве координирующего центра нервной систе-
мы появился уже у таких примитивных животных
как дожившие до настоящего времени плоские чер-
ви. Но эти существа, как и большинство беспоз-
воночных, руководствовались главным образом
инстинктом; разумное поведение стало возможным
только с появлением более крупного и сложного
мозга.
Уже у ранних рыб мозг, как и у нас, состоял и
трех основных частей: задней, средней (мезэн
|
|
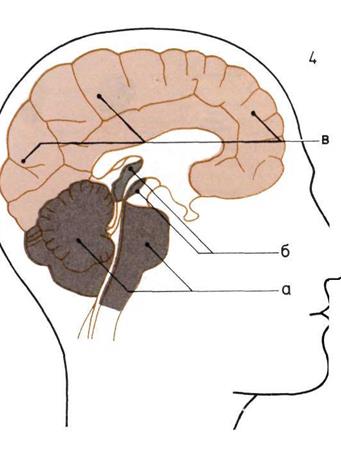
цефалон) и передней (диэнцефалон). Но функции
мозга рыб очень ограниченны. Его задняя часть
обеспечивает равновесие, средняя отвечает за зрение,
а передняя ведает главным образом обонянием.
У пресмыкающихся сравнительно сильнее развиты
задняя и средняя части мозга, что способствует
улучшению слуха, зрения и координации деятель-
ности органов чувств; все эти эволюционные изме-
нения помогают приспособиться к наземному образу
жизни.
 У ранних млекопитающих мозг становится зна-
У ранних млекопитающих мозг становится зна-
чительно более крупным и сложным, чем у пресмы-
кающихся. В задней части мозга появляется крупный
мозжечок, похожий по своей форме на кочешок ка-
пусты; он осуществляет координацию сложных дви-
жений. Функция координации деятельности органов
чувств передалась в переднюю часть, из которой
развился крупный, обладающий многочисленными
извилинами отдел головного мозга, управляющий
процессами запоминания и обучения. Даже у мле-
копитающих с маленьким мозгом, например у ежа,
передняя часть мозга преобладает над средней.
У более высокоразвитых млекопитающих, например
у обезьян, передние доли мозга становятся относи-
тельно еще более крупными (этот процесс рассматри-
вается в нескольких следующих главах). Огромный
головной мозг человека — местопребывание разу-
ма -- перерос и затмил своими размерами осталь-
ные части мозга, значительно преобразовав при
этом сам череп.
|
|
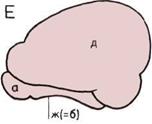
Развитие коры головного мозга
(слева).
На схематическом изображении
(вид сбоку) прослеживаются
тенденции развития коры
левого полушария. Большая
часть серого вещества, богатого
нервными клетками, собирается
на поверхности мозга, образуя
его кору, или паллиум
(«плащ»).
А — Примитивная форма мозга.
Б — Земноводное.
В — Примитивное
пресмыкающееся.
Г — Высокоразвитое
пресмыкающееся.
Д — Примитивное
млекопитающее.
Е — Высокоразвитое
млекопитающее.
Изображены следующие
органы.
а — Обонятельная луковица.
б — Палеоиаллиум (старая
плашевидная часть головного
мозга).
в — Архипаллиум
(примитивная плетевидная
часть).
 г — Базальные нервные центры.
г — Базальные нервные центры.
д — Неопаллиум (новая
плащевидная часть, богатая
ассоциативными центрами).
е — Гиппокамп, или аммониев
рог (прежний архипаллиум).
ж — Обонятельная доля мозга
(прежний палеопаллиум, или
«обонятельный мозг»).
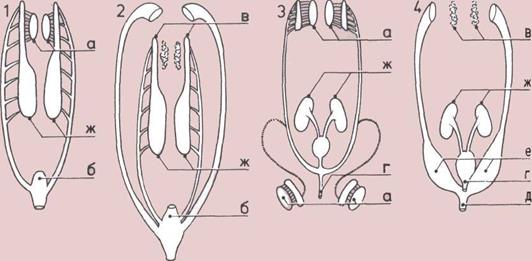
Как возникли легкие (вверху).
На продольных разрезах видно
общее происхождение легких
у наземных позвоночных
и плавательного пузыря у рыб.
1.
Примитивное легкое
двоякодышащей рыбы развилось
из горлового кармана (мешка,
камеры).
2.
Модификация легкого
двоякодышащей рыбы.
3.
Наполненный воздухом
плавательный пузырь,
придающий плавучесть
костистым рыбам.
4.
Легкое наземного
позвоночного с имеющей много
складок внутренней стенкой,
которая позволяет дышать
эффективнее, чем более простые
легкие рыбы.
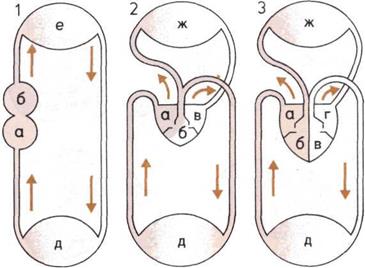
Развитие кровообращения
(справа).
1.
Кровообращение рыбы;
двухкамерное сердце нагнетает
венозную кровь из тела в жабры,
свежая кровь поступает из
жабр в тело.
2.
Кровообращение одного
из пресмыкающихся;
трехкамерное сердце нагнетает
смешанную кровь в легкие и тело.
3.
Кровообращение
млекопитающих;
четырехкамерное сердце не дает
смешиваться венозной
и артериальной крови.
а — г Камеры сердца.
д — Тело.
 е — Капилляры жабр.
е — Капилляры жабр.
ж — Легкие.
Легкие, система кровообращения и почки осущест-
вляют жизненно важные функции. Легкие снабжа-
ют тело кислородом, необходимым для «сжигания»
пищи и обеспечения жизненных процессов энергией,
а также удаляют из организма отработанный газ —
двуокись углерода. Кровь поставляет клеткам тре-
бующиеся им питательные вещества и кислород.
Почки очищают кровь и удаляют вредные отходы
жизнедеятельности тела.
Легкие по своему происхождению, вероятно, вос-
ходят к влажным мешочкам в горле кистеперых рыб,
в которых они накапливали атмосферный воздух,
необходимый для жизни в болотных лужах с низ-
ким содержанием кислорода. Легочные мешки зем-
новодных и даже пресмыкающихся еще и сейчас
имеют участки поверхности, приспособленные для
поглощения кислорода. Легкие млекопитающих
крупнее по размерам и сложнее по строению. У зем-
новодных воздух накачивается мышцами горла, а
у млекопитающих своего рода вакуумный насос при-
водится в действие мышцами ребер и диафрагмой.
 У двоякодышащих рыб нос, выполнявший перво-
У двоякодышащих рыб нос, выполнявший перво-
начально лишь функции обоняния, превратился
в орган, который позволяет всасывать воздух в лег-
кие и предотвращает попадание в них воды. У прес-
мыкающихся надо ртом появилось костное нёбо,
частично отделяющее пищу от вдыхаемого воздуха.
У млекопитающих к костному нёбу добавилось мяг-
кое нёбо, позволяющее не прерывать дыхание во
время еды, что жизненно необходимо для теплокров-
ных животных.
Наша система кровоснабжения унаследовала
очень архаичные черты: так, соленый вкус крови
свидетельствует о том, что нашими предками явля-
ются простые одноклеточные организмы, приспо-
собленные к обитанию в морской среде. Однако
система кровообращения подверглась коренному
преобразованию. Древняя система сердце — жабры,
имевшаяся у предков современных рыб, преобразова-
лась в систему, состоящую из сердца и кровеносных
сосудов, по которым венозная кровь подается в лег-
кие, а свежая, обогащенная кислородом кровь по-
ступает из легких в клетки по всему телу. У наших
холоднокровных земноводных и пресмыкающихся
предков легкие развивались быстрее, чем система
кровообращения. Только у птиц и млекопитающих
имеется четырехкамерное, так сказать «двустволь-
ное», сердце, в котором свежая (артериальная) кровь
полностью отделена от венозной. Такая эффектив-
ная система кровообращения делает нас теплокров-
ными и позволяет нашему телу поддерживать рав-
ный темп жизнедеятельности — вне зависимости от
времени суток и температуры окружающей среды.
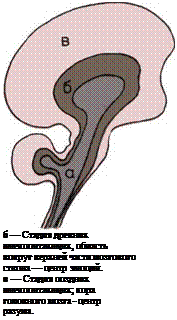
Почки, по-видимому, произошли от имевшихся
у беспозвоночных крошечных изолированных тру-
бочек, по которым отбросы жизнедеятельности орга-
низма откачивались из полостей между органами.
У рыб эти насосы сгруппировались в виде двух
длинных полосок, расположенных внутри тела; к ним
добавились очищающие кровь фильтры — почки,
и отбросы организма стали удаляться через спе-
циальное выводное отверстие — клоаку. У пресмы-
кающихся почки образовали два плотных скопления,
сосредоточенных в дальней задней части тела. У че-
ловека и прочих млекопитающих каждый из ка-
нальцев почек приобрел длинный завиток, выводя-
щий часть воды обратно в тело, что препятствует
его высыханию.
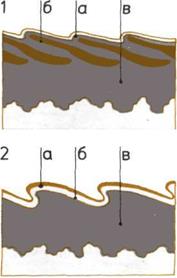
Уменьшение числа артерий
(внизу).
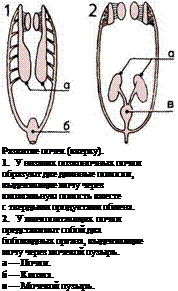 На пяти рисунках показано
На пяти рисунках показано
уменьшение числа артерий,
сопровождавшее переход
от жабр к легким.
А — Примитивная рыба.
Б — Рыба, дышащая легкими.
В — Земноводное.
Г — Пресмыкающееся.
Д — Млекопитающее.
1—6 — Дуги аорты
(первоначально связанные
с жабрами).
а — Камеры сердца.
б — Легкие
|
|
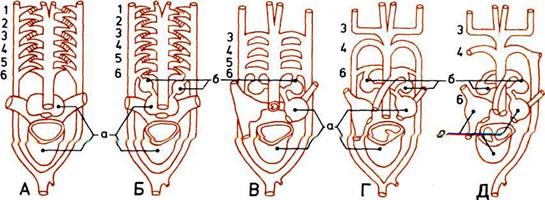
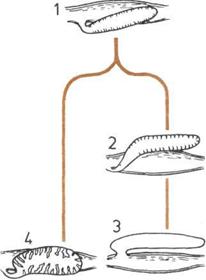
Развитие половых органов.
Изменение мужских и женских
половых органов способствовало
переходу к внутреннему
оплодотворению.
1.
Самец низшего позвоночного
животного; сперма из яичек
выходит через клоаку.
2.
Самка низшего позвоночного;
яйцеклетки из яичников выходят
через клоаку.
3.
 Самец млекопитающего;
Самец млекопитающего;
сперма из яичек (обычно
опушенных) проходит через
мочеиспускательный канал
полового члена (не показан
на рисунке), а затем попадает
во влагалище.
Способ, посредством которого оплодотворяется
человеческая яйцеклетка, и ее последующий рост
в материнском теле совершенно непохожи на под-
верженную рискованным случайностям манеру раз-
множения наших далеких предков. Подобно их совре-
менным родственникам, эти древние морские бес-
позвоночные, по всей вероятности, беспорядочно
метали в море огромные количества мужских и жен-
ских клеток. У позвоночных же развились более
целенаправленные и эффективные способы выделе-
ния половых клеток, гарантирующие оплодотворение
яиц спермой.
У примитивных позвоночных половые железы
испускают сперму и яйца в полости тела, откуда они
выделяются через поры, расположенные поблизости
от отверстий, выводящих из тела отбросы. Но у рыб
развились особые трубки для выведения наружу
продуктов деятельности половых желез.
 Земноводные, подобно рыбам, мечут свою не
Земноводные, подобно рыбам, мечут свою не
имеющую твердой скорлупы икру в воду, где она
оплодотворяется вне тела самки. У пресмыкающихся
способ размножения приспосабливается к наземному
образу жизни. Самцы вводят сперму непосредствен-
но внутрь тела самок, где влага предохраняет спер-
матозоиды от гибели. Оплодотворение яйцеклетки
внутри тела позволяет самкам пресмыкающихся
откладывать меньше яиц, чем, скажем, лягушка или
а — Яички.
б — Клоака.
в — Яичники.
г — Мочеиспускательный канал.
д— Влагалище.
е — Матка.
ж — Почки.
|
|
жаба, чтобы обеспечить достаточную для сохране-
ния данного вида численность потомства.
Яйца пресмыкающихся приспособлены для раз-
вития в наземных условиях. Они покрыты скорлу-
пой, окружены пленкой — мембраной и содержат
вместилище жидкости, что предохраняет зародыш
от повреждений и высыхания. В яйцах имеются
также значительный запас пищи в виде желтка,
своеобразные «легкие» и коллектор для отходов жиз-
недеятельности.
Человек и прочие млекопитающие унаследовали
от пресмыкающихся основные особенности строе-
ния этого так называемого амниотического яйца,
но в то время как большинство пресмыкающихся
откладывают яйца и оставляют их в прогретом солн-
цем грунте до тех пор, пока вылупятся детеныши,
яйца плацентарных млекопитающих развиваются
в тепле и безопасности в материнской утробе. Здесь
временный орган — плацента — снабжает еще не
родившегося детеныша большим количеством пита-
тельных веществ, чем то, которое могло поместиться
внутри яйца, ограниченного скорлупой; через пла-
центу же осуществляется и выделение из зародыша
продуктов распада. Это новшество в способе размно-
жения оказалось столь успешным, что наш вид про-
должает процветать, несмотря на длительную бере-
менность, заканчивающуюся обычно рождением
лишь одного беспомощного малыша.
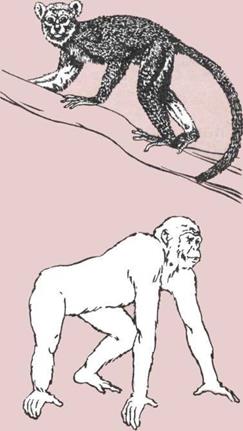
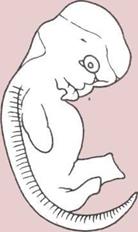
Эмбрионы и эволюция.
На трех разрезах
прослеживаются изменения в
защите и питании зародыша,
развивающегося в теле матери.
А — Эмбрион земноводного.
Б — Эмбрион пресмыкающегося.
В — Эмбрион человека
(млекопитающего).
а — Эмбрион.
б — Желток (запас пиши).
в— Белок (запас пищи).
г — Оболочка (защита).
д — Водяная оболочка, или
амнион (амортизирующий
мешочек).
![]() е — Хорион (воздушное
е — Хорион (воздушное
пространство, через которое
поступает кислород и удаляются
газовые отбросы).
ж — Аллантоис, мочевой
мешок (место, где собираются
отходы метаболизма).
з — Плацента (временный
орган с хорноновой или
аллантоидной структурой);
проходящая через плаценту
кровь матери снабжает эмбрион
питательными веществами
Рудиментарные органы.
![]()
![]() Эти три органа, возникшие
Эти три органа, возникшие
в ходе эволюции, сохраняются
у человека, хотя и не выполняют
сейчас никаких полезных
функций.
а — Аппендикс.
б — Ушные мышцы.
в — Копчик (рудиментарная
хвостовая кость).
|
|
|
|
Эмбриональное
развитие человека
Сходство человеческого эмбриона с эмбрионами
других животных является убедительным доводом в
пользу нашего родства с другими формами жизни.
У каждого животного, от червя до человека, рост
тела начинается со всего лишь двух слоев деля-
щиеся клеток — внешнего (эктодермы) и внутрен-
него (эндодермы), которые соответствуют двум
слоям клеток, образующим тело таких примитивных
беспозвоночных, как медуза. Из эктодермы разви-
ваются кожа и нервная ткань; из эндодермы — внут-
реннее покрытие пищеварительного тракта и свя-
занные с ним органы, такие как поджелудочная же-
леза и печень, а также жаберные или легочные ткани
и некоторые другие органы. Но у всех позвоноч-
ных из третьего среднего слоя — мезодермы —
развивается большинство частей тела, включая кос-
ти, мышцы, артерии и вены, органы половой и моче-
вой систем и хрящевой стержень (хорда), являю-
щийся предшественником позвоночника.
На ранней стадии жизни эмбриона клетки мезо-
дермы по обоим бокам хорды образуют сегменты
(сомиты), являющиеся двусторонними зачатками
сегментированного тела позвоночных. На этой ста-
дии человеческий зародыш напоминает зародыш
рыбы: имеет подобный рыбьему мозг, «жаберные
мешки» на шее, зачатки конечностей, похожие на
плавники; разделенные на сегменты мышцы вдоль
туловища; хвост; сердце и почки, как у рыбы, и не-
сколько больших парных кровеносных сосудов, по-
добных тем, которые у рыб соединяют сердце и
жабры.
Однако вскоре первоначальные органы, делающие
зародыш человека похожим на рыбий, подвергают-
ся изменениям в направлении уподобления его мле-
копитающему. Жаберные перегородки не превра-
щаются в жабры, а образуют лицо, челюсти, гор-
тань, язык, уши и эндокринные органы, располо-
женные в шее. Из потенциального рыбьего плава-
тельного пузыря развиваются легкие. Большинство
парных кровеносных сосудов, соединяющих сердце
и жабры, уменьшаются в размерах и исчезают.
Артерии конечностей перестраиваются и начинают
снабжать кровью ноги и руки. В сердце образуются
перегородки; почки приобретают форму, характер-
ную для млекопитающих. Туловище утрачивает
часть мыши — появляется талия. Через некоторое
время о хвосте зародыша напоминают только срос-
шиеся кости в нижней части позвоночника.
За период между четвертой и шестой неделями
развития человеческий эмбрион превращается из
рыбоподобного организма в организм, неотличимый
от зародыша обезьяны. В двухмесячном возрасте
зародыш уже, без всякого сомнения, является кро-
шечным человеческим существом.
|
|
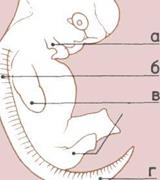
Зародыш человека (вверху).
Органы человеческого
зародыша, аналогичные
органам зародышей других
позвоночных.
а — Жаберные карманы.
б — Сомиты (сегменты тела).
в — Зачатки конечностей.
г — Хвост.
Доказательства эволюции
(слева).
Три столбца рисунков
изображают развитие рыбы,
свиньи и человека — их
превращение из яйца в эмбрион
и затем во взрослый организм.
Ранние стадиии развития
каждого из позвоночных
аналогичны, и это является
доказательством того, что все эти
три существа произошли от
одних и тех же древних
позвоночных животных.
1 — Рыба.
2 — Свинья.
3
![]() — Человек.
— Человек.
Несмотря на многочисленные преимущества,
форма нашего тела имеет свои изъяны. Один автор
даже заявил, что человек — это «несовершенное
создание, настоящий винегрет признаков».
Большинство наиболее важных проблем связано
с вертикальным положением скелета, первоначаль-
ная конструкция которого отчасти была похожа на
горизонтальный арочный мост. Теперь, для того что-
бы мы могли поворачивать туловище и наклоняться,
у нас появились клиновидные позвонки, опираю-
щиеся на свои массивные передние края. Но это
ослабляет нижнюю часть позвоночника, и подыма-
ние тяжестей может привести к смещению позвонка
или износившегося межпозвоночного хряща (диска),
что вызывает острую боль в спине. Кроме того, в
связи с изгибом основания позвоночника некоторые
позвонки заходят довольно глубоко в родовые пути,
доставляя неприятности матерям во время родов.
![]()
![]() Поскольку тело опирается лишь на две конечно-
Поскольку тело опирается лишь на две конечно-
сти, ноги иногда страдают от перегрузки, а свод
стопы прогибается, что приводит к плоскостопию,
изменению формы костей и даже к искривлению
пальцев ног и появлению сумок на их наружной
поверхности.
Выпадение межпозвоночного
диска.
Выпадение хрящевого диска,
являющегося буфером между
двумя спинными позвонками,
создает болезненную нагрузку
на спинномозговой нерв
и уменьшает подвижность.
1. Нормальный диск.
2.
Выпавший диск.
а — Позвонок.
б — Нерв.
в — Диск.
Части тела, создающие
особые проблемыдля человека.
Справа внизу показаны
некоторые части скелета,
системы кровообращения
и других органов, которые
создают особые проблемы для
человека.
А — Зубы.
Слишком тесное расположение
зубов является следствием
уменьшения размеров челюстей,
которое произошло в процессе
эволюции сравнительно недавно.
Б — Выпадение диска.
Заболевания нижней части
позвоночника обычно являются
следствием дегенеративных
возрастных изменений,
связанных с передачей нагрузки
через позвоночник на две, а не
на четыре конечности, как это
было у наших древних
млекопитающих предков.
В — Аппендицит.
Это заболевание связано
с инфекционным заражением
и воспалением аппендикса —
рудиментарного червеобразного
отростка слепой кишки.
Г— Грыжа.
Выпячивание кишок в слабых
частях брюшной стенки.
У мужчин и женщин грыжа
обычно возникает в несколько
разных местах.
Д — Варикозное расширение
вен.
Это заболевание клапанов вен,
приводящее к скоплению крови,
может поражать как бедра,
так и ноги.
Е — Плоскостопие.
Ослабление свода стопы широко
распространено и связано с тем,
что вес человеческого тела
приходится на ступни только
двух ног.
Еще одним наказанием за переход к хождению на
двух ногах является грыжа. У четвероногих живот-
ных кишечник прикреплен к позвоночнику широкой
связкой; у человека кишечник меньше и закреплен
не столь прочно. Поэтому кишки могут выпячиваться,
выпирая вперед через ослабленную брюшную стенку.
Вертикальное положение тела мешает даже кро-
воснабжению. Для того чтобы вернуться от ног к
сердцу, кровь должна преодолевать силу тяжести
на протяжении примерно 1,2 м. Если поврежден-
ные клапаны ножных вен не могут воспрепятство-
вать обратному стоку крови, то мы испытываем
тяжесть и усталость в ногах и другие симптомы вари-
козного расширения вен. Плохой отвод венозной
крови может также быть причиной воспаления бед-
ренной вены левой ноги при беременности, когда
усиленное давление внутренних органов прижимает
эту вену к острой кости в месте соединения двух
позвонков в нижней части позвоночника.
![]() Ухудшение проходимости родового канала в ре-
Ухудшение проходимости родового канала в ре-
зультате двуногого хождения и увеличение черепов
младенцев, создающие трудности при деторожде-
нии, а также другие неприятности — все это след-
ствия несовершенного устройства нашего тела.
|
|
Серологические реакции
(вверху).
По силе реакции антиген —
антитело при введении сыворотки
чужой крови можно судить
о биологическом родстве: чем
сильнее реакция, тем теснее
предполагаемое родство.
а — Введение сыворотки крови
человека кролику.
б — Сыворотка крови кролика
с антителами к сыворотке крови
человека.
в — Сыворотка крови человека
(реакция 100%).
г — Сыворотка крови шимпанзе
(97%).
д — Сыворотка крови бабуина
(50%).
![]() е — Сыворотка крови собаки (0).
е — Сыворотка крови собаки (0).
К числу наиболее убедительных доказательств
эволюционного происхождения человека относятся
данные, полученные в результате сравнения био-
химических структур — кирпичиков, из которых
слагаются тела всех живых организмов.
Клетки тела человека и почти всех других живот-
ных и растений содержат аденозинтрифосфат
(АТФ), используемый во внутриклеточных энерге-
тических процессах, а также дезоксирибонуклеино-
вую кислоту (ДНК) — наследственное вещество,
передаваемое через гены.
Различия между биомолекулами человека и других
организмов помогают установить большую или мень-
шую степень нашего родства с некоторыми другими
группами. Так, фосфокреатин имеется в мышцах
человека и других хордовых (животных со спинным
стержнем — хордой), но его нет у большинства бес-
позвоночных.
Специалисты придумали множество изощренней-
ших опытов, чтобы определить, какие из ныне живу-
щих хордовых наиболее близки нам в биохимиче-
ском отношении.
Молекулярное древо жизни.
Сравнение молекул гемоглобина
48 плацентарных
млекопитающих, относящихся
к семи группам, говорит
о наличии эволюционных связей,
показанных на этой схеме.
Шкала времени (в миллионах
лет) основана на информации,
полученной при изучении
ископаемых остатков.
а — Грызуны.
б — Копытные.
в — Насекомоядные.
г — Хищные.
д — Зайцеобразные (кролики
е — Полуобезьяны тупайя
(возможно, насекомоядные).
ж — Приматы (лемуры, лори,
долгопяты, низшие обезьяны,
человекообразные обезьяны
и, крайний справа, человек).
| и др.).
|
О том, насколько сильно протеины различных
видов животных отличаются друг от друга, можно
судить по иммунологическим исследованиям, пока-
зывающим, как различные животные реагируют на
чужую сыворотку крови.
В число других опытов входит сравнение после-
довательности аминокислот, из которых состоит дан-
ный протеин. Чем более схожа эта последователь-
ность у различных животных, тем, как предполага-
ется, в более тесном родстве друг с другом они на-
ходятся. Труднее выявить изменения последователь-
ности нуклеотидов, из которых построены длинные
цепочки молекул ДНК. В качестве так называемого
«конечного предка» ДНК является хранительницей
того, что было названо «окончательной истиной мо-
лекулярной эволюции».
Взятые в своей совокупности данные прямого и
косвенного сравнения молекул подтверждают выво-
ды анатомии и физиологии: ближайшие из ныне
живущих наших родственников относятся к той груп-
пе, в которую входят человекообразные и прочие
обезьяны, и, следовательно, у нас должны быть об-
щие с ними предки.
Далее (см. с. 88—89) мы познакомимся с раз-
личными, подчас спорными, гипотезами молекуляр-
ной биологии о времени расхождения генетических
ветвей, к которым принадлежат обезьяны и человек.
| 0 |
|||||||
| 1 |
0 |
||||||
| 9 |
8 |
0 |
|||||
| 'М |
11 |
10 |
6 |
0 |
|||
| А £ |
14 |
15 |
18 |
17 |
0 |
||
| 21 |
21 |
17 |
17 |
26 |
0 |
||
| 31 |
30 |
26 |
27 |
31 |
32 |
0 |
|
| 45 |
45 |
45 |
46 |
47 |
47 |
47 |
0 |
|
|
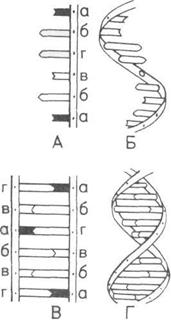
Генетический код (вверху).
Генетический код. определяющий
принадлежность к данному виду.
представляет собой
последовательность оснований,
связывающих две нити
нуклеотидов, которые образуют
двойную спираль ДНК
А — Основания на участке одной
из нитей нуклеотидов.
Б — Фрагмент одной из нитей.
В — Соединение
комплементарных
(дополняющих друг друга)
оснований.
Г — Две нити, соединенные
и скрученные в виде спирали.
а — Тимин.
б — Гуанин.
в — Цитозин.
г — Аденин.
Последовательности
аминокислот (слева).
Организмы различаются но
последовательности аминокислот
в белковом нитохроме c
ферменте, который необходим
для производства энергии. Чем
больше указанный на диаграмме
номер, тем значительнее
различия между любыми двумя
существами.
а — Человек.
б — Обезьяна макака-резус.
в — Кролик.
г — Утка.
д — Гремучая змея.
е — Тунец.
ж — Мотылек.
з — Дрожжевой грибок.
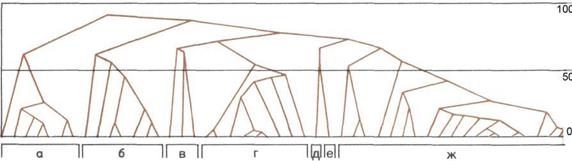
Эволюционное древо приматов.
На схеме показаны возможные
связи между современными
и вымершими группами приматов
по классификации Шалая
(Szalay) и Делсона (Delson).
Их схема несколько отличается
от традиционных классификаций.
Номерами 1—4 обозначены
подотряды: буквами — а—д —
подчиненные им группы; шкала
времени дана в миллионах лет.
1. Предок приматов,
2. Подотряд Plesiadapiformes.
3.
Подотряд Strepsirhini.
а — Adapiformes.
б — Lemuriformes (лемуры и
лори).
4.
Подотряд Haplorhini.
в—Tarsiiformes (долгопяты).
г — Platyrrhini (широконосые),
низшие обезьяны Нового Света.
д — Catarrhini (узконосые),
низшие обезьяны Старого Света,
человекообразные обезьяны
и человек.
Хотя по общим особенностям нашего организма
мы принадлежим к млекопитающим, по совокупно-
сти более детальных анатомических признаков нас
можно отнести к приматам, т. е. к тому же отряду
высокоподвижных млекопитающих, что и лемуры,
низшие и человекоподобные обезьяны. По всей види-
мости, большинство особенностей, характерных
для приматов, развились в результате приспособле-
ния к жизни, проводимой главным образом на вет-
вях деревьев.
Во-первых, на руках и ногах всех приматов име-
ются, как правило, пять гибких пальцев, на которых
растут не когти, а ногти. Большие пальцы рук и
обычно ног противопоставлены прочим пальцам, что
удобно для хватания предметов. Чувствительные
кисти на конце длинных подвижных рук служат
в качестве органов осязания.
Коренные зубы со сравнительно низкими высту-
пами и увеличенная нижняя часть кишечника с фер-
ментационной камерой, состоящей из слепой кишки
и аппендикса, приспособлены для измельчения и пе-
реваривания растительной пищи, хотя многие при-
маты всеядны.
При древесном образе жизни приматам бвио лег-
че осуществлять связь между собой при помощи
зрения, а не обоняния, и поэтому у них носы короче
и обоняние развито слабее, чем у многих других
млекопитающих. Зрение же приматов неимоверно
обострилось.
Большие, направленные вперед глаза с острвш
трехмерным видением (цветовым у ведущих днев-
| i ! |
4д| |
0 |
|||
| i 2 |
jf За • |
20 |
|||
| « |
„ • . i0 |
||||
| # 60 • и |
|||||
ной образ жизни) позволили приматам точно оце-
нивать расстояние при прыжках с ветки на ветку.
Увеличившийся в размере мозг имеет сильно раз-
витые участки, которые управляют деятельностью
органов ощущения и координируют движения, что
необходимо при акробатических прыжках, совер-
шающихся в считанные доли секунды.
Укорочение лицевой части, увеличение глаз и
мозга, а также вертикальное положение тела, идеаль-
но пригодное для того, чтобы сидеть на ветке, — все
это привело к изменению формы черепа приматов.
После длительного периода беременности у боль-
шинства самок рождается один детеныш, который
сосет молоко из двух молочных желез на груди
матери. Молодняк долго находится под надзором
матерей: такова стратегия выживания при обитании
на деревьях существ, чье поведение определяется
скорее навыками, приобретенными в процессе обуче-
ния, чем инстинктами.
И наконец, большинство приматов извлекают
выгоды из группового образа жизни, который позво-
ляет им обмениваться сведениями о местах нахожде-
ния пищи, объединяться для совместной защиты
и передавать друг другу усвоенные навыки поведе-
ния.
В нескольких последующих главах речь пойдет о
тенденциях развития различных групп приматов,
эволюция которых привела к появлению человека.
Признаки, характерные для
приматов.
Общими для человека и других
приматов являются следующие
характерные признаки
(обозначенные буквами д— з.
встречаются также у различных
низших млекопитающих).
а — Увеличенный мозг с двумя
уникальными складками.
б — Большие, обращенные
вперед глаза (глазницы
окружены костными
прикрытиями).
в — Специфическое строение
среднего уха.
г — Зубы «примитивного» типа.
д — Ключица.
е — Раздельные лучевая
и локтевая кости.
ж — Раздельные
большеберцовая и малоберцовая
кости.
з — Пять подвижных пальцев
на каждой конечности.
и — Большие пальцы рук обычно
противопоставлены остальным
пальцам, что позволяет
захватывать предметы.
к — Четко выраженная пяточная
кость.
л — Плоский ноготь
на больших пальцах ног и, как
правило, ногти (а не когти) на
остальных пальцах.
Место человека в природе.
![]()
![]()
![]() Многие специалисты считали,
Многие специалисты считали,
что классификационная система
приматов, к которым относится
человек, включает восемь
иерархических уровней.
Придерживаясь новой точки
зрения, мы помещаем человека
и человекообразных обезьян
в одно семейство. В этом случае
классификация современных
и некоторых вымерших приматов
обретает вид, показанный
на рисунке внизу слева.
![]()
![]()
![]()
![]() Мы начнем с вызывающей
Мы начнем с вызывающей
много споров гипотезы о
появлении приматов ранее
60
миллионов лет назад и дадим
краткий обзор подотряда
низших приматов, которых
называют также полуобезь-
янами. На этих страницах
пойдет речь как о вымерших
ранних формах, так и об их
современных потомках:
лемурах, лори и долгопятах.
Среди их предшественников
были и предки человека.
![]()
|
|
|
|

| Характерные признаки |
| Характерные признаки а — Зубы похожи на зубы б — Среднее ухо не менее Зубы приматов. А — Зубы предполагаемого Б — Зубы современной в — Предкоренные зубы |
Развитие среднего уха.
На рисунке показано, как
возможные изменения в ходе
эволюции различных
полуобезьян привели
к появлению эктвтимпанума
прочного кольца или трубки,
поддерживающих барабанную
перепонку уха приматов.
а — Эктотимпанум.
б — Продольное сечение
слуховой полости.
1.
Предполагаемый предок
приматов.
2. Низший примат.
3. Мегаладапид.
4. Лори.
5. Плезиадапид.
6.
![]()
![]() Древний долгопят.
Древний долгопят.
Первые приматы появились, вероятно, в Северной
Америке или Европе около 70 миллионов лет назад.
Они имели разные размеры: одни были величиной
с мышь, другие с домашнюю кошку. Произошли они,
возможно, от насекомоядных, похожих на тупайю
(см. с. 50—51).
У первых приматов, как и у их отдаленного потом-
ка — человека, сохранилось по пяти пальцев на каж-
дой конечности. Эта примитивная особенность, как
выяснилось, была более полезной, чем узко специа-
лизированные конечности таких млекопитающих,
как киты или лошади, оказавшихся навсегда свя-
занными с одной определенной средой обитания или
с определенным образом жизни.
Ранние приматы имели не ногти, а когти, и у них
не было таких больших, направленных вперед глаз
и такой укороченной лицевой части, как у большин-
ства поздних приматов, но их коренные зубы уже
походили на зубы обезьян и человека. До недавнего
времени ученые объединяли ранних приматов, а так-
же лемуров, лори и долгопятов, живущих ныне в
тропических лесах Африки, Мадагаскара и Азии, в
один подотряд, названный Prosimii («до-обезьяны»).
Согласно более современной классификации, низ-
шие приматы делятся на два подотряда: примитив-
ных Plesiadapiformes и более высокоразвитых Strep-
sirhini, которые научились лучше прыгать, хватаясь
при этом за ветки.
Расцвет полуобезьян совпал с эпохами палеоцена
и эоцена, когда возникали, сменяя друг друга, десят-
ки новых видов животных.
Некоторые из первых полуобезьян, вероятно, оби-
тали преимущественно на земле. Однако 60 миллио-
нов лет назад конкуренция со стороны появившихся
в это время грызунов вынудила полуобезьян перейти
к жизни на деревьях. Теперь острые зубы ранних
насекомоядных приматов сменились перемалываю-
щими молярами, приспособленными для измель-
чения листьев и фруктов; челюсти приводились
в движение с помощью мыши, прикрепленных к ши-
роким и сильным скуловым костям. Тем временем
конечности тоже изменились, приспосабливаясь к
прыжкам и лазанию. Глаза сдвинулись в переднюю
часть головы и стали фокусироваться на одном и
том же предмете, что позволило совершать более
точные прыжки с ветки на ветку и облегчило охоту
за насекомыми.
Однако 30 миллионов лет назад полуобезьяны на-
чали сдавать позиции, явившись жертвами похоло-
дания климата на континентах Северного полушария
и конкуренции со стороны своих собственных потом-
ков — обезьян.
| зиклмн о п ш 4*rt* W№gb. 3( 0 |
||||||||
| • в # |
• • |
• • |
• |
1 |
10 |
|||
| • |
• • |
• • « * * |
• • |
р ^20 |
||||
| • |
• * • • • • |
• • # • |
30 |
|||||
| а |
• |
» # • |
40 |
|||||
| ---- ------------------------------------------------------ ■ qflf |
• » • • • |
50 |
||||||
| (во . М- М^- 2 1 1 :1^ : |
3 « |
1 fin |
||||||
| ■ Г» 1ПГ— # • -Г- - - ч |
---- ------ --------------------------------------------------------------- „ 70 |
|||||||
| • |
||||||||
| • ^ ^ Миллионы лет назад * — — - ------------------------------------------------------------------------------------------------------------------------- |
||||||||
| Родословное древо полуобезьян. 2. Strepsirhini.
|
|||||||||||||||||||||||||||||||||||||||||
Дрейф континентов.
На трех картах показано, как
произошло разделение
континентов, которое привело
к изоляции некоторых популяций
ранних приматов.
1. 200 миллионов лет назад.
2.
65—46 миллионов лет назад
(палеоцен — средний эоцен).
3.
46—38 миллионов лет назад
(поздний эоцен).
![]()
![]() а — Лавразия.
а — Лавразия.
б — Еврамерика.
в — Азиамерика.
г — Северная Америка.
д — Евразия.
Ранние приматы унаследовали от своих предков
мир, подвергающийся серьезным изменениям. Когда
65 миллионов лет назад закончился меловой период,
два доисторических сверхконтинента Лавразия (се-
верный) и Гондвана (южный) раскололись на куски.
В палеоцене и эоцене массы суши начали приобре-
тать свои современные очертания и занимать совре-
менное положение. Северная Америка и Европа
отделились друг от друга, а Южная Америка превра-
тилась в остров, сначала лишь узкой полоской океана
отделенный от Африки. Между тем на некоторых
континентах мелководные, врезавшиеся в сушу моря
то отступали, то продвигались вперед. В это же
время выросли ранние Анды, Скалистые горы и дру-
гие горные хребты.
Перемещение континентов повлияло на климат.
Вначале Северная Африка и Европа купались в теп-
ле. Но к олигоцену (36—26 миллионов лет назад)
повсюду произошло похолодание климата. Это при-
вело к изменению растительного мира. Субтропиче-
ские леса, буйно разросшиеся ранее на значитель-
ной части Северной Америки и Европы, уступили
теперь место морозостойким деревьям и травам.
Все это повлияло на плацентарных млекопи-
тающих, среди которых начали быстро возникать
разнообразные виды, приспособленные к новому
образу жизни и к тем средам обитания, которые
освободились в результате вымирания динозавров
в конце мелового периода. Ранние плацентарные
млекопитающие смогли теперь широко распростра-
ниться по Евразии и Северной Америке, где и появи-
лось большинство ранних приматов. Если горы или
океаны разделяли один и тот же вид на отдельные
популяции, в некоторых из них возникали новые
разновидности, а другие виды просто вымирали.
Ко времени палеоцена от плацентарных млекопи-
тающих размером с крысу произошли крупные виды,
среди которых особенно выделяются примитивные
копытные травоядные — кондилартры и ранние хищ-
ные млекопитающие — креодонты. Ко времени эоце-
на они столкнулись со все возраставшей конкурен-
цией со стороны ранних современных копытных и
хищников. Тем временем грызуны вытеснили ранних
приматов с земли на деревья.
Животный мир палеоцена.
Здесь представлены девять
доисторических животных
палеоцена (масштабы не
соблюдены). Рисунки в—и
отражают взрывообразную
эволюцию млекопитающих.
Беспозвоночные.
а — Тентакуляты
(щупальцевые).
Земноводные.
б — Червяги.
Млекопитающие.
в — Амблиподы.
г — Шерстокрылы.
д — Приматы.
е — Кондилартры (древние
копытные).
![]()
![]()
![]() ж — Многобугорчатые
ж — Многобугорчатые
(мультитуберкуляты).
з — Непарнокопытные.
и — Грызуны.
~ ^ W-CC
![]()
![]()
![]() ■ ■
if
■ ■
if
Животный мир эоцена.
Эти 18 доисторических животных
были наиболее распространены
в эоцене (масштабы не
соблюдены).
Беспозвоночные.
![]()
![]() а — Кишечнополостные.
а — Кишечнополостные.
б — Черви нематоды.
к — Ламантины.
Хоботные.
Непарнокопытные.
Парнокопытные.
Неполнозубые.
Киты.
Птицы.
р — Бескилевые птицы.
Килевые птицы.
Считается, что первые приматы составляли под-
отряд Plesiadapiformes («близкие к Adapiformes»).
Большинство из них специалисты относят к прима-
там исходя из строения внутреннего уха. Но их череп,
конечности и длинные, более низко посаженные сред-
ние резцы, которые выступали вперед, обладали
примитивными чертами, не встречающимися у более
поздних приматов. Эта группа, вероятно, произош-
ла от ранних насекомоядных, но многие из состав-
лявших ее видов питались жесткой растительной
пищей. Появилось, по крайней мере, пять семейств,
в которые входило 20 сильно отличавшихся друг от
друга родов размерами от мыши до кошки. Многие
из них, возможно, были похожи на грызунов, но
большинство известно нам только по ископаемым
зубам и осколкам черепа. От какой-нибудь из этих
ранних форм, возможно, и произошли все остальные
приматы.
![]() Эта группа жила 70—40 миллионов лет назад
Эта группа жила 70—40 миллионов лет назад
и, очевидно, распространилась из Северной Америки
в Европу через Гренландию, которая представляла
собой тогда теплый, поросший лесами сухопутный
переход, связывавший эти два континента. Возьмем
в качестве примера представителей четырех из этих
семейств.
|
|
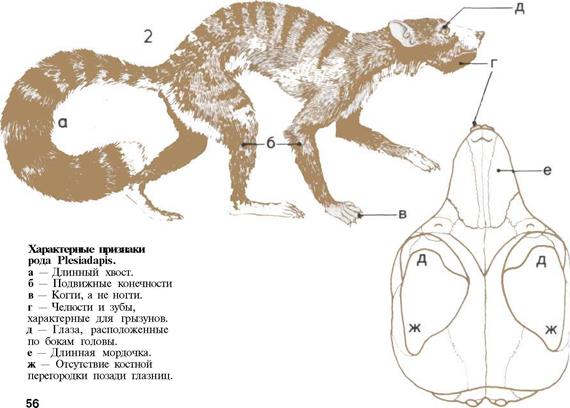
1. Purgatorius — древнейший из известных нам
приматов, величиной с крысу. Имел четыре предко-
ренных зуба (это больше, чем у позднейших при-
матов) и острые трехбугорные коренные зубы, та-
кие, как у насекомоядных, но с добавочным выступом,
именуемым «пяткой», что обеспечивало увеличение
жевательной поверхности. Время — поздний мел —
палеоцен. Место — штат Монтана (США). Семей-
ство Paromomyidae.
2. Plesiadapis был похож скорее на белку с длин-
ной мордочкой, расположенными по бокам глазами,
выступающими вперед долотообразными резцами,
пушистым хвостом и когтями на лапках, не приспо-
собленных для хватания. Он питался листьями, хоро-
шо прыгал и, возможно, жил стадами, часто не на
деревьях, а на земле. Время — средний палеоцен —
ранний эоцен. Место — штат Колорадо (США) и
Франция. Семейство Plesiadapidae.
3. Carpodaptes был величиной с мышь, имел ко-
ренные зубы с пилообразными краями и огромный
предкоренной зуб (премоляр) для резания жесткой
растительной пищи. Время — палеоцен — ранний
эоцен. Место — Северная Америка. Семейство Сагро-
lestidae.
4. ![]() Zanycteris был крошечным, похожим на мышь
Zanycteris был крошечным, похожим на мышь
приматом с коренными зубами, как у летучей мыши.
Он мог питаться нектаром цветов, фруктами или
насекомыми. Время — поздний палеоцен. Место —
Северная Америка. Семейство Picrodontidae.
Череп карподапта.
![]() Как и другие плезиадапиды,
Как и другие плезиадапиды,
Carpodaptes имел длинные
выступающие вперед нижние
резцы (а), а также крупные
нижние предкоренные зубы (б).
Длина черепа около 1 дюйма
(2,54 см).
|
|
|
1. Notharetus имел довольно длинную мордочку,
гибкую спину, длинные тонкие ноги; его большие
пальцы были отделены от прочих. Свой длинный
хвост он использовал в качестве балансира при ла-
зании на верхушки деревьев и при прыжках. У него
были небольшие клыки, маленькие вертикальные
резцы и полнь!й набор коренных зубов, включая
моляры с низкой коронкой, способные растирать
листья, превращая их в мягкую кашицу. Длина —
33 дюйма (84 см). Время — средний эоцен. Место —
штат Вайоминг, США. (Обитал преимущественно в
Северной Америке, но встречался также в Европе,
Африке и Азии). Подсемейство Notharctinae.
2. ![]() Adapts несколько похож на Notharctus, но его
Adapts несколько похож на Notharctus, но его
сильные короткие челюсти были способны двигаться
лишь вверх и вниз и не могли растирать пищу боко-
выми движениями. Adapts, вероятно, питался жест-
кими волокнистыми растениями. У него был не-
сколько примитивный головной мозг, но со сравни-
тельно более крупными височными долями, чем у
млекопитающих не-приматов. Длина — 16 дюймов
(40 см). Время — средний и поздний эоцен. Место —
Западная Европа. Подсемейство Adapinae (Европа
и Северная Америка).
Черепа адапид.
Черепа нотарктуса (А)
и смилодектеса (Б)
(разновидность с укороченной
лицевой частью) имеют
характерные для адапид
особенности, не встречающиеся
у плезиадапид.
а — Направленные вперед
глаза, обеспечивающие
стереоскопическое видение.
б — Костяная пластинка,
защищающая глаза сзади.
в — Относительно крупный мозг.
г — Укороченная лицевая
часть.
![]()
![]() д — Более вертикальное
д — Более вертикальное
положение резцов.
|
|

|
|
1. Lemur catta— ныне живущий лемур. У него
типичная собачья мордочка лемуроида, 36 зубов,
голый носик, большие пушистые уши, подвижные
конечности, цепкие кисти рук и стопы ног с ногтями,
длинный пушистый хвост, в окраске которого чере-
дуются темные и светлые кольца. Семейство Lemu-
ridae.
2. Indri indri — индриобразный лемуриформ,
имеет длинные конечности, короткий и толстый хвост;
цепляется за деревья и прыгает по ним вверх. Се-
мейство Indriidae.
3. Daubentonia madagascariensis, ай-ай, имеет за-
остренную мордочку, огромные уши, пушистый хвост,
когтистые пальцы и всего лишь 18 зубов. Своими рез-
цами грызуна он вскрывает кокосовые орехи, а гиб-
ким средним пальцем давит насекомых — древо-
точцев. Семейство Daubentoniidae.
4. Galago crasslcaudatus, толстохвостый галаго,
имеет собачью мордочку, большие глаза, подвиж-
ные уши, длинный пушистый хвост и широкие поду-
шечки на кончиках пальцев. Длина — 25 дюймов
(64 см). Место — Африка. Семейство Lorisidae (Этот
крупнейший из современных галаго, возможно, по-
хож на Progalago раннего миоцена.)
5. Nyctlcebus coucang, толстый лори, у него корот-
кая мордочка, большие глаза, тонкие запястья и ло-
дыжки; хвоста нет. Длина — 13 дюймов (33 см). Ме-
сто — Юго-Восточная Азия. Семейство Lorisidae.
Пяль лемуровдов.
Нумерация рисунков
соответствует принятой в тексте.
1. Lemur catta.
2. Indri indri.
3.
Daubentonia
madagascariensis.
4.
![]() Galago crassicaudatus.
Galago crassicaudatus.
5. Nycticebus coucang.
|
|

|
|
К Tarsiiformes относится свыше 30 родов мелких
приматов с укороченными мордочками; все они,
кроме одного рода, уже вымерли. Три сохранившихся
вида долгопятов имеют большие глаза, ведут ночной
образ жизни и похожи на сказочных домовых. Но
именно они, быть может, являются нашими ближай-
шими родственниками после обезьяны, так как у дол-
гопята, обезьян и человека много сходных черт в
строении носа и глаз, структуре головного мозга,
строении тканей и в биохимическом составе клеток
тела. И действительно, многие специалисты отно-
сят секцию Tarsiiformes наряду с высшими прима-
тами к одному и тому же подотряду Haplorhini
(«простые носы»).
![]()
![]()
![]() Tarsiiformes произошли в палеоцене от Adapifor-
Tarsiiformes произошли в палеоцене от Adapifor-
mes, а может быть, от Plesiadapiformes. Они дали
разнообразные формы и распространились по кон-
тинентам Северного полушария и Африке. Их рас-
цвет пришелся на эоцен, но в олигоцене большинст-
во из них вымерло, возможно, потому, что они не
могли соперничать с появившимися в это время
обезьянами.
а — Длинный тонкий хвост.
б — Длинная лодыжка.
помогающая прыгать по
верхушкам деревьев.
в — Наполовину сросшиеся
болыпеберцовая
и малоберцовая кости.
г — Длинные тонкие пальцы
кистей и стоп.
д — Присоединение
позвоночника к черепу снизу,
а не сзади (поэтому голова
обычно держится прямо).
Приведем примеры из двух семейств.
1. Necrolemur имел короткую мордочку, большие,
направленные вперед глаза, тяжелые выдвинутые
нижние передние зубы и длинные пяточные кости.
Он цеплялся вертикально за ствол дерева и, воз-
можно, не сгибаясь прыгал с ветки на ветку. Разме-
ры такие же, как и у современного долгопята (2).
Время — эоцен. Место — Европа. Семейство Ото-
myidae.
2. ![]() Tarsius, современный долгопят, имеет сухой,
Tarsius, современный долгопят, имеет сухой,
покрытый шерстью нос, 34 зуба, огромные глаза,
большие уши, короткие руки, длинные гибкие ноги
с длинными таранными костями, подушечки на кон-
чиках пальцев и короткие ногти (но на каждой из
ног есть по два пальца с когтями, служащими для
причесывания). Длина — 4,7 дюйма (12 см), не счи-
тая длинного тонкого хвоста. Питается насекомыми
и ящерицами. Место — Юго-Восточная Азия (вы-
мерший Afrotarsius жил в Африке). Семейство
Tarsiidae.
Долгопяты — предок и потомок
(масштаб не соблюден).
Нумерация рисунков
соответствует принятой в тексте.
1. Necrolemur.
2.
![]()
![]() Tarsius.
Tarsius.
![]()
![]() На этой гравюре XIX века
На этой гравюре XIX века
изображена бесхвостая
берберийская обезьяна Старого
Света (магот), которая
встречается в Северо-Западной
Африке и па мысе Гибралтар.
![]()
Низшие и человекообразные обезьяны н человек_____
высшие приматы — традиционно объединяются в
один большой подотряд приматов Anthropoidea.
Антропоиды произошли от предков ныне живущих
маленьких долгопятов (или, может быть, лемуров).
Впервые они появились в Северной Америке или
Евразии около 40 миллионов лет назад. Этот под-
отряд разделился на две секции: Platyrrhini, широко-
носые (низшие обезьяны Нового Света), и Catar-
rhini, узконосые (низшие обезьяны Старого Света,
человекообразные обезьяны и человек).
Современные антропоиды различны по размерам:
от крошечных игрунков (тамаринов) размером с
белку до горилл, весящих в три раза больше, чем
средний человек. Животные, принадлежащие к неко-
торым из видов антропоидов, ходят на четвереньках,
другие цепляются за деревья руками, ногами, хво-
стом. Только человек постоянно передвигается на
задних конечностях. Хвосты некоторых обезьян
длиннее их тела; у человекообразных обезьян, как и
у человека, хвостов нет. Один вид обезьян ведет ноч-
ной образ жизни, прочие антропоиды — дневной.
![]() Но если мы отвлечемея от этих различий, то уви-
Но если мы отвлечемея от этих различий, то уви-
дим, что большинству антропоидов присущи опре-
деленные общие основные черты. Большинство из
них могут сидеть прямо, что высвобождает руки для
держания различных предметов, захватывать кото-
рые помогают большие пальцы рук и ног, которые
могут загибаться внутрь. У большинства видов на
пальцах рук и ног имеются плоские ногти и отсутст-
Эволюционное древо
антропоидов.
![]() Линиями показаны возможные
Линиями показаны возможные
эволюционные связи между
семействами антропоидов
и их вероятными предками
(см. также главу 2).
а — Omomyidae.
б — Cebietae.
в — Atelidae.
г — Parapithecidae.
д — Oreopithecidae.
е — Cercopithecidae.
ж — Pliopithecidae.
з — Proconsulidae.
и — Hylobatidae.
к — Hominidae.
вуют те когти, которые служат лемурам для расче-
сывания своей шерсти.
Голова антропоидов наклонена вперед под углом
к спинному хребту. Лицевая часть, как правило,
укорочена, но глаза увеличены и направлены вперед,
причем каждый глаз не просто защищен костяной
пластинкой, как это было у более примитивных по-
луобезьян, а помещен в защитную костяную полость.
Округленный череп защищает мозг, который срав-
нительно крупнее, чем у полуобезьян; извилин в
мозгу антропоидов больше, и у них иной рисунок;
участок мозга, управляющий обонянием, имеет мень-
шие размеры, а те участки, которые отвечают за зре-
ние и мыслительную деятельность, наоборот, увели-
чены.
Все 30—36 зубов расположены вертикально. В их
число входят укрупненные коренные зубы с низкими
буграми, предназначенные не просто для разреза-
ния жесткой пищи и ввгжимания из нее сока, но и
для ее полного измельчения; низкие предкоренные
зубы (премоляры) часто цепляются за верхние клы-
ки, стачивая и заостряя их. Такие зубы позволяют
многим антропоидам питаться любой пищей.
![]() У антропоидов только две молочнвге железы, кото-
У антропоидов только две молочнвге железы, кото-
рые находятся на груди. Их плацента отличается от
плаценты полуобезьян; по сравнению с этими при-
матами, у антропоидов дольше период беременно-
сти, а также больше продолжительность жизни.
|
|
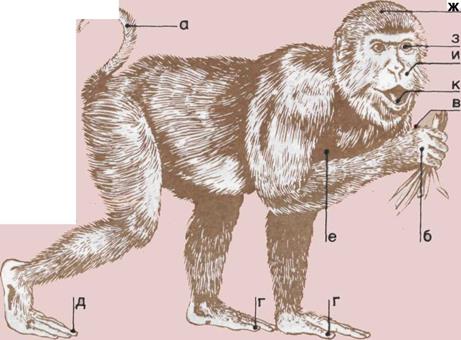
| Характерные признаки а — Хвост длинный, короткий и — Лицевая часть выступает |
Мир в миоцене.
На этой карте мира видно, что
в миоцене континенты заняли
свое современное положение.
Линиями отмечены экватор,
тропики и полярные круги.
Сокращение размеров морей
(внизу).
![]() На рисунке показана часть
На рисунке показана часть
Африки и Евразии около
16 миллионов лет назад.
Сокращение размеров моря
Тетис и появление новой
сухопутной перемычки
позволили антропоидам
мигрировать между Африкой,
Европой и Азией.
а — Море Тетис.
б — Море Пара-Тетис.
в — Аравийская сухопутная
перемычка.
Ко времени миоцена (25—5 миллионов лет назад)
число разнообразных видов антропоидов значитель-
но возросло и они заселяли уже Африку, Европу и
тропическую Америку.
В эту эпоху мир подвергся значительным измене-
ниям. Ледяной щит покрыл Антарктику, Индия при-
соединилась к Азии, выросли величайшие горные
хребты мира. В Африке разверзлась Рифтовая до-
лина под аккомпанемент извержений вулканов.
Огромное доисторическое море Тетис превратилось
в большие соленые лужи: Средиземное, Черное и Кас-
пийское моря. Животные легко мигрировали из Евра-
зии в Африку и обратно. Листопадные, широко-
лиственные вечнозеленые и хвойные деревья покры-
вали теперь континенты Северного полушария. Даль-
ше к югу размеры тропических лесов сокращались
по мере того, как климат становился более сухим.
В Восточной Африке тропические леса все еще окайм-
ляли реки и нижние склоны гор, но великие равнины
превратились в саванну — открытые, похожие на
парки пространства, заросшие высокой травой, среди
которой разбросаны отдельные деревья.
Похолодание климата Северного полушария,
которое началось в олигоцене (38—25 миллионов лет
назад), вероятно, способствовало эволюции антро-
поидов. Там, где уменьшалось количество листьев
и фруктов, которые можно было употреблять в пищу
круглый год, полуобезьяны большей частью либо
просто вымирали, либо двигались на юг вслед за
отступающим тропическим лесом. По всей видимости,
антропоиды произошли от тех приматов, которые
проявляли больше гибкости в выборе рациона пита-
ния и легче переносили холод. В тех местах, где
|
|
| Современная береговая |
в определенные сезоны фруктов становилось меньше,
их доля в пищевом рационе, естественно, должна
была сокращаться, а доля коры и листьев вечнозеле-
ных растений возрастать. Увеличивались размеры
тела антропоидов, что способствовало сохранению
в нем тепла. Вероятно, это осуществлялось наряду
с удлинением сроков беременности, сокращением
числа новорожденных и переходом от ночного образа
жизни к дневному, вследствие чего зрение приобре-
тало все большее значение по сравнению с обоня-
нием..
Миграция животных.
На этой диаграмме дана схема
миграций млекопитающих
в эпоху миоцена, когда
континенты Северного
полушария были соединены
сухопутными перешейками.
Однако южноамериканские
обезьяны так никогда и не
перешли в Северную Америку;
возможно, им помешали
климатические барьеры.
Эволюция обезьян Нового Света
проходила исключительно
в Южной Америке, которая
тогда была островом. Не
появились приматы и в
Австралии.
|
|
| СЕВЕРНАЯ |
| ЖIt |
![]()
Platyrrhini («плоские носы») — это обезьяны тро-
пиков Нового Света. У них широко расставленные,
вывернутые наружу ноздри; три предкоренных зуба
по обеим сторонам каждой челюсти; сравнительно
более крупный мозг с большим числом извилин, чем
у Tarsiiformes; несколько черепных костей, имеющих
отчетливо выраженные сочленения; длинные (иног-
да цепкие) хвосты; их большие и прочие пальцы сла-
бо развиты и плохо приспособлены для хватания.
Размеры этих обезьян различны: от крошечных
игрунков до ревунов, весящих около 22 фунтов
(10 кг).
Обезьяны Нового Света, возможно, произошли
около 40 миллионов лет назад от североамериканских
Tarsiiformes, но распространились только в Цент-
ральной и Южной Америке. Древнейшей из ископае-
мых находок является Branisella из нижнего олигоце-
на Колумбии (около 35 миллионов лет назад). Третья
часть всех 20 известных нам родов вымерла, но 35%
современных антропоидов относятся к широконо-
сым обезьянам.
![]() Мы даем ископаемые и современные примеры
Мы даем ископаемые и современные примеры
из двух семейств, классификация которых была не-
давно пересмотрена на основе строения их черепа
и зубов.
в — Относительно более
крупный и сложный головной
мозг, чем у долгопятовых.
г — 36 зубов, в том числе по три
предкоренных по обеим сторонам
каждой челюсти.
Для всего семейства Ateiidae
характерны следующие
признаки.
![]()
![]()
![]() д — Голые, без волос уши.
д — Голые, без волос уши.
е — Короткие кривые, но
похожие на ногти, когти.
1. Neosaimiri, возможно, являлись предками бе-
личьих обезьян (саймири), у которых маленькие
выразительные мордочки, а хвост в три раза длиннее
тела. Длина — 20 дюймов (50 см). Время — сред-
ний миоцен. Место — Колумбия. Семейство Atelidae.
2. Callithrix jacchus, обыкновенный игрунок, имеет
пучки волос на ушах, 32 зуба, длинные нижние резцы,
когти на большинстве пальцев и ногти на больших
пальцах ног. Семейные группы питаются лесными
растениями и насекомыми. Длина — 20 дюймов
(50 см). Время — настоящее. Место — Бразилия.
Семейство Cebidae.
3. Stirtonla, возможный предок ревунов, была,
вероятно, крупной сгорбленной бородатой обезьяной
с цепким хвостом; питалась листьями; издавала
громкий рев, чтобы отпугнуть соперников. Длина —
3 фута 7 дюймов (1,1 м). Время — средний миоцен.
Место — Колумбия. Семейство Atelidae.
4. Cebupithecia имела длинные, узкие, выступаю-
щие вперед резцы, крупные клыки и небольшие, тесно
посаженные предкоренные зубы (премоляры); пита-
лась фруктами. Возможно, она была похожа на со-
временных бородатых саки. Длина — 31 дюйм
(80 см).
Время — средний миоцен. Место — Колумбия. Се-
мейство Atelidae.
5. ![]()
![]() Tremacebus имел огромные глаза и, возможно,
Tremacebus имел огромные глаза и, возможно,
был похож на трехполосную дурукули — единствен-
ную из современных обезьян, ведущую ночной образ
жизни; питался фруктами и насекомыми. Длина —
30 дюймов (75 см). Время — поздний олигоцен.
Место — Аргентина. Семейство Atelidae.
Пять широконосых обезьян.
Нумерация рисунков
соответствует принятой в тексте.
1.
Neosaimiri (по внешнему виду
несколько напоминают белку).
2. Callithrix jacchus.
3.
![]() Stirtonia (похожа на
Stirtonia (похожа на
обезьяну-ревуна).
К узконосым Catarrhini («загнутые носы») отно-
сятся низшие обезьяны Старого Света, человекооб-
разные обезьяны и человек. Известно около 40 родов,
свыше половины из которых сейчас вымерли.
Ноздри узконосых посажены близко друг к другу
и направлены вниз или вперед. Их большие и прочие
пальцы лучше приспособлены для хватания, чем
пальцы низших обезьян Нового Света, но у них от-
сутствует цепкий хвост. У многих поздних низших
обезьян развились укороченные хвосты; современ-
ные человекообразные обезьяны, как и человек,
хвоста не имеют. У всех узконосых сравнительно
крупный мозг и сходно устроенные ушные отверстия;
все они, за исключением некоторых примитивных
видов, имеют по два предкоренных зуба (премоляра)
по обеим сторонам каждой челюсти. Узконосые раз-
личны по величине: от Apidium размером с белку до
огромной человекообразной обезьяны Giganto-
pithecus (обе давно вымерли).
![]() Эта группа обезьян, вероятно, произошла от Tar-
Эта группа обезьян, вероятно, произошла от Tar-
siiformes 40 миллионов лет назад. Родиной самых
ранних из известных нам узконосых была Южная
Азия, но затем они дали разнообразные формы и
размножились по Африке и всей Евразии.
|
|
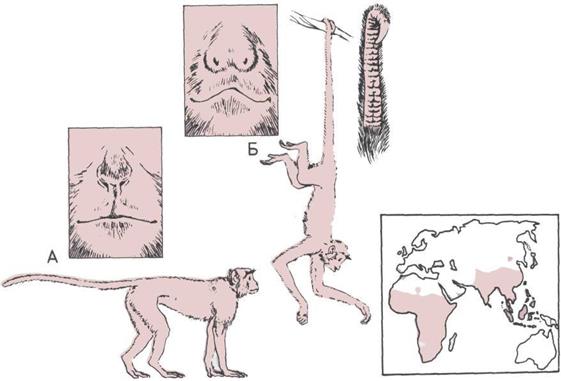
| Хвосты и ноздри. На рисунках показаны основные А — Обезьяна Старого Света Места обитания. На этой карте показано |
К приведенным здесь примерам относятся неко-
торые возможные предшественники человекооб-
разных и низших обезьян Старого Света.
1. Pondaungia известна только по истертым об-
ломкам ископаемой челюсти с широкими коренными
зубами (молярами), покрытыми толстым слоем
эмали. Она, возможно, была похожа на проплиопи-
тека (Propliopithecus) (см. с. 78—79). Вероятно,
эта обезьяна лазала по деревьям и питалась фрук-
тами. Время — поздний эоцен. Место — Пандаун
(Бирма). Семейство не определено.
2. Oligopithecus, которую называют по-разному:
то «первой несомненной человекообразной обезья-
ной», то «отчасти полуобезьяной», возможно, явля-
лась предком длинноруких гиббонов. Наличие греб-
ней на коренных зубах говорит о том, что она пита-
лась листьями и насекомыми. Размеры — как у
маленькой обезьянки. Время — ранний олигоцен.
Место — Египет. Семейство не определено.
3. ![]()
![]()
![]()
![]() Apidium была по размерам не крупнее беличьей
Apidium была по размерам не крупнее беличьей
обезьяны (саймири), на которую она, возможно,
была похожа. Вероятно, она бегала по ветвям на
четвереньках и разжевывала своими многобугор-
чатыми зубами фрукты и насекомых. Время — сред-
ний олигоцен. Место — Египет. Семейство Parapithe-
cidae (похожие на низших обезьян антропоиды;
вымерли к концу олигоцена).
Низшие обезьяны Старого Света семейства мар-
тышкообразных (Cercopithecidae) вместе со стран-
ной, ныне вымершей обезьяной Oreopithecus состав-
ляют надсемейство антропоидов Cercopithecoidea,
которое, возможно, произошло от семейства плио-
петицидовых.
![]()
![]()
![]() Миоценовые и более поздние ископаемые находки
Миоценовые и более поздние ископаемые находки
дают только слабое представление об истории низ-
ших обезьян Старого Света. Большинство из них —
это млекопитающие малых или средних размеров,
имеющие большой, хорошо развитый мозг, крупный
округлый череп; только по два предкоренных зуба
по обеим сторонам каждой челюсти; маленькие
наружные уши и длинный или короткий, но не цепкий
хвост. Существуют два надсемейства: Cercopitheci-
nae и Colobinae. Обезьяны Cercopithecinae (мартыш-
ковые) имеют защечные мешки, более всеядны и
менее связаны с древесным образом жизни, чем
колобины; живут они в Африке и Азии. К ним отно-
сятся геноны, бабуины, макаки и гелады. Вы можете
увидеть здесь на рисунках ореопитека (Oreopi-
thecus) и трех мартышковых обезьян.
1. Oreopithecus, крупный ранний антропоид, об-
ладал особенностями, характерными как для низ-
ших, так и для высших обезьян и гоминид. Он цеп-
лялся своими длинными руками за ветки, но иногда
мог и ходить на двух ногах. Рост 3 фута 11 дюймов
(1,2 м). Время — поздний миоцен. Место — Север-
ная Италия и, возможно, Юго-Западная Россия.
Семейство Oreopithecidae (эволюционный тупик, в
котором оказались предки низших обезьян Старого
Света).
2. Cercopithecus, геноны, маленькие, передвигаю-
щиеся на четвереньках обезьянки, хвосты которых
длиннее тела. Большинство из них живут на деревьях
и питаются плодами. Время — от позднего плиоцена
до сих пор. Место — Африка.
3. Papio, павианы, имеют крупные черты лица;
это ходящие на четвереньках наземные жители
(самцы значительно крупнее самок), с мощными
руками и ногами одинаковой длины, выгнутым дугой
хвостом, удлиненной мордой, похожей на собачью,
и мощными клыками. Павианы живут стаями, пита-
ются растениями и мелкими животными. Время —
от позднего плиоцена до наших дней. Место — Аф-
рика и юго-запад Аравии.
4. Масаса, макаки, сильные обезьяны малых и
средних размеров с длинными округлыми мордоч-
ками; их руки и ноги имеют одинаковую длину; у
некоторых отсутствует хвост. Время — от позднего
миоцена до наших дней. Место — от Северо-Запад-
ной Африки до Японии.
=9 Q
Зубная формула.
Каждая сторона верхней челюсти
павиана-самца соответствует
типичной зубной формуле
антропоидов Старого Света.
а — Два резца.
б — Один клык.
в — Два предкоренных зуба.
г — Три коренных зуба.
Представители семейства
церкопитекоидов.
Нумерация рисунков
соответствует принятой в тексте.
1. Oreopithecus.
2. Cercopithecus.
3. Papio.
4. Масаса.
|
|

Характерные признаки колобин.
![]()
![]() На скелете этой ископаемой
На скелете этой ископаемой
обезьяны (Mesopithecus)
наблюдаются следующие
характерные признаки, которые
позволяют отнести ее к
колобинам.
Низшие обезьяны Старого Света, у которых боль-
шие пальцы рук укорочены или отсутствуют, обра-
зуют подсемейство мартышковых Colobinae. Это
название происходит от греческого слова "kolobur",
что означает «покалеченный». У колобин обычно
длинные тела, конечности и хвосты; большинство из
них превосходно лазают по деревьям и совершают
прыжки на огромные расстояния высоко в воздухе.
Их зубы и желудки хорошо приспособлены к пере-
работке большого количества пищи, состоящей в
основном из листьев.
![]()
![]()
![]() Колобины, вероятно, появились в Африке пример-
Колобины, вероятно, появились в Африке пример-
но в середине миоцена и распространились по Евра-
зии. Имелось около десяти родов колобин, половина
из которых вымерла, но оставшиеся насчитывают
много видов и распространены главным образом в
Южной Азии.
В наших примерах представлены различные под-
группы.
1. Victoriapithecus, одна из самых ранних извест-
ных нам мартышковых; являлась, возможно, пред-
ком колобин. Но те несколько ископаемых находок,
которые объединены этим именем, относятся к раз-
личным родам. Размер — маленький. Время — от
раннего до среднего миоцена. Место — Восточная
Африка.
2. Mesopithecus имел длинные сильные конечности;
большие пальцы рук у него длиннее, чем у какой-
либо из современных колобин. Возможно, от него
произошли худощавые длинноногие длиннохвостые
лангуры современной Азии. Mesopithecus обитал
в похожих на парк ландшафтах и часто ходил на
четвереньках по земле. Размер — от маленького до
среднего. Время — от позднего миоцена до позднего
плиоцена. Место — Европа и Юго-Восточная Азия.
3. Colobus - это родовое название обитающих
в Африке проворных колобин с длинными конечно-
стями, которых называют также гверецами. У них
отсутствует большой палец руки; мордочка корот-
кая, а верхняя губа немного нависает над нижней.
Большинство из них живут высоко на деревьях.
Размеры — от 18 до 24 дюймов (46—60 см), не счи-
тая еще более длинного хвоста.
4. ![]() Presbytis, лангуры и другие питающиеся ли-
Presbytis, лангуры и другие питающиеся ли-
стьями азиатские обезьяны, похожи на Colobus,
но у них имеются короткие большие пальцы на руках,
а у многих на шее есть волосатая грива или хохо-
лок. Лангуры ведут дневной образ жизни; живут ста-
ями. Длина 16—31 дюйм (40—80 см), не считая еще
более длинного хвоста. Время — от позднего миоце-
на до наших дней. Место — от Пакистана до Индо-
незии.
|
|
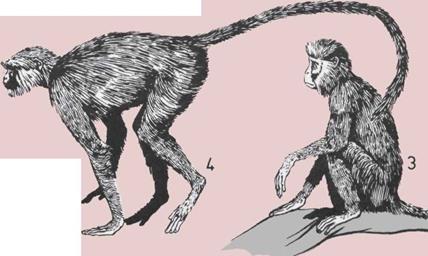
| Четыре колобины. Нумерация рисунков 1.
Victoriapithecus 2. Mesopithecus. 3. Colobus. 4. Presbytis. |
| л |
|
| ^ а |
|
| i |
& |
| Места обитания гилобатидов. а — Материковая часть |
Признаки гиббонов.
Большинство из них присуще
также человекообразным
обезьянам и человеку.
а — Крупные размеры тела.
б — Отсутствие хвоста.
в — Широкая грудная клетка.
г — Очень подвижный плечевой
сустав.
![]()
![]() д — Крупная мозговая коробка.
д — Крупная мозговая коробка.
Человекообразные обезьяны и человек относятся
к надсемейству узконосых Hominoidea. Их передние
зубы не такие, как у низших обезьян Старого Света,
а руки и плечевой пояс очень подвижны, и на них
удобно висеть. У современных гоминоидов нет хво-
стов; их мозг и тело больше, чем у низших обезьян.
Кроме того, скорость их размножения и развития
более медленная.
Гоминоиды и низшие обезьяны Старого Света,
вероятно, появились в Африке к концу олигоцена;
их предками были маленькие ранние антропоиды
из семейства Pliopithecidae. Специалисты придер-
живаются различных мнений относительно при-
надлежности, значения и родственных связей тех или
иных ископаемых форм. В этой книге мы исходим
из наличия трех семейств гоминоидов: Hylobatidae
(гиббоны и сиаманги), Proconsulidae (гоминоиды
«основного ствола») и Hominidae (крупные чело-
векообразные обезьяны, человек и их ближайшие
предки).
![]() е — Руки длиннее ног.
е — Руки длиннее ног.
ж — Подвижное запястье.
з — Длинные цепкие кисти.
и — Большой палец руки
противопоставлен остальным
пальцам.
Мы видим на этих рисунках трех плиопитекидов
и одного гилобатида.
1. Pliopithecus, плиопитек, когда-то считавшийся
предком гиббонов, был величиной с гиббона и имел
такую же короткую мордочку, как у гиббона; боль-
шие глаза и крупные острые клыки, но, в отличие от
гиббона, у него не было столь длинных рук и, воз-
можно, был хвост. Он бегал, лазал, прыгал и висел
на деревьях. Время — миоцен. Место — Европа.
Семейство Pliopithecidae.
2. Propliopithecus (называемый также Aegypto-
pithecus) мог быть малой или средней величины; те-
лосложением походил на обезьяну ревуна и имел
длинное, опущенное вниз лицо. Он бегал на четве-
реньках и лазал, хватаясь за деревья руками и но-
гами. Время — средний олигоцен. Место — Египет.
Семейство Pliopithecidae.
3. Dendropithecus был не больше гиббона, имел
хрупкое телосложение, как у паукообразной обезь-
яны (коаты). Он лазал, прыгал и висел на деревьях.
Время — ранний миоцен. Место — Кения. Семейство
Pliopithecidae.
4. ![]()
![]()
![]()
![]()
![]() Hylobates, гиббоны, — это небольшие широ-
Hylobates, гиббоны, — это небольшие широ-
когрудые так называемые «меньшие человекообраз-
ные обезьяны». Живут семейными группами, пита-
ются плодами, громко кричат; висят, цепляясь за
деревья руками, которые у них значительно длиннее
ног. Гиббоны изредка ходят на двух ногах. Длина
головы и тела 16—26 дюймов (40—65 см). Время —
от плейстоцена до наших дней. Место — Юго-Восточ-
ная Азия. Семейство Hylobatidae.
В этой книге вымершие Dryopithecinae рассмат-
риваются как одно из трех подсемейств, образую-
щих семейство Hominidae (в два других подсемей-
ства входят крупные человекообразные обезьяны,
человек и вымершие «почти люди»). У гоминид, как
правило, руки, плечевой пояс и запястья в некоторой
степени приспособлены для того, чтобы качаться на
деревьях; грудь широкая, поясница укороченная,
но основание спины (сросшиеся кости крестца) длин-
ное; есть рудиментарная хвостовая кость (копчик)
и расширенные верхние тазовые кости. Зубы и че-
реп с широкой и часто удлиненной лицевой частью
очень похожи на зубы и череп других антропоидов
Старого Света, но размеры тела и относительная
величина мозга обычно больше.
![]()
![]()
![]()
![]()
![]()
![]() Dryopithecinae («древесные обезьяны») — это ран-
Dryopithecinae («древесные обезьяны») — это ран-
ние человекообразные обезьяны, которые, вероятно,
появились в Африке в миоцене и пришли в Европу
|
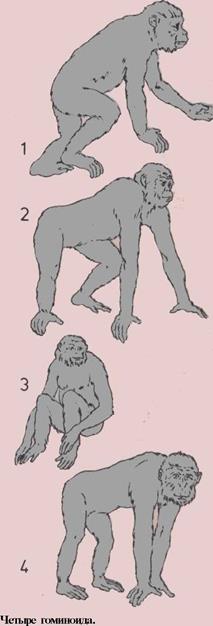
|
|
|
Морды и зубы (вверху).
А — Горилла, сравнительно
длинная морда и крупные клыки.
Б — Рамапитек, более короткая
морда и меньшие по размерам
клыки.
Схема родственных связей.
На этом упрощенном
эволюционном древе гоминоидов
буквами А—В — обозначены
семейства, а буквами а—в —
подсемейства.
![]()
![]()
![]() А — Proconsulidae.
А — Proconsulidae.
Б — Hylobatidae (гиббоны).
В — Hominidae
(человекообразные обезьяны
и человек).
а — Dryopithecinae
б — Ponginae.
в — Homininae.
К подсемейству гоминид, которое называется
Ponginae, относятся предки единственной совре-
менной крупной азиатской человекообразной обезь-
яны — орангутана (см. с. 86—87). Ponginae, воз-
можно, появились в Африке, но распространились
в Европе и Азии и существовали примерно от 17
до 1 миллиона лет назад. Большинство из них были
меньше человека, но некоторые доисторические пон-
гины превосходили самцов горилл.
Ископаемые находки, среди которых имеются
редуцированные (уменьшенные в размерах) резцы
и крупные коренные зубы, покрытые толстым слоем
эмали, говорят о том, что эти животные обитали
в более открытой местности, чем дриопитецины, и
питались более сухой и грубой пищей. Это, вероятно,
явилось следствием приспособления к условиям оби-
тания в открытой лесистой местности, которая сме-
нила дождевые леса в тех районах, где в позднем
миоцене сократилось количество атмосферных
осадков.
![]()
![]() Гипотеза о человекообразной обезьяне, которая
Гипотеза о человекообразной обезьяне, которая
вела наземный образ жизни, наряду с предполагае-
мым сходством зубов, челюстей и строения черепа
с человеческими, привела многих специалистов к
выводу о том, что Ratnapithecus ходил на двух ногах,
пользовался орудиями труда и являлся прямым пред-
шественником той эволюционной линии, от которой
произошел человек. Скептики не согласны с этой
точкой зрения, хотя один специалист и отождествил
Ramapithecus с Sivapithecus, назвав его человеко-
образной обезьяной4
от которой шел путь к человеку.
1. Sivapithecus был похож лицом на орангутана,
костями ступни — на шимпанзе и мог вращать за-
пястьями. Он, возможно, лазал по деревьям, мог
висеть на ветках и ходил на четвереньках. Размеры —
примерно, как у орангутана. Время — около 15—9
миллионов лет назад. Место — Европа и Южная
Азия.
2. Ramapithecus, как предполагают, являлся
уменьшенным вариантом Sivapithecus, но у него были
укороченная впалая лицевая часть и более мелкие,
несколько плоские зубы со сравнительно большей
поверхностью кусания, чем у человека и крупных
человекообразных обезьян. Причем эти зубы сидели
в глубокой челюсти, которая была короче и острее,
чем у человека, крупных человекообразных обезьян
и Dryopithecinae. Масса — около 31 фунта (14 кг).
Время — 15—7 миллионов лет назад. Место — Ев-
ропа и Азия.
3. ![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]() Gigantopithecus, последняя и самая крупная
Gigantopithecus, последняя и самая крупная
в своей группе огромная человекообразная обезь-
яна со впалым коротким лицом, мощными челюстя-
ми, довольно маленькими клыками, которыми она
пользовалась, как и коренными зубами, для расти-
рания пищи, и с очень большими молярами. Высо-
та — до 9 футов 10 дюймов (3 м). Время — 10—1
миллион лет назад. Место — Азия, от Пакистана
до Южного Китая.
![]() ЧЕЛОВЕКООБРАЗНЫЕ
ЧЕЛОВЕКООБРАЗНЫЕ
ОБЕЗЬЯНЫ И ЧЕЛОВЕК
![]()
![]() К середине 1980-х годов у
К середине 1980-х годов у
ученых не было бесспорных
ископаемых доказательств
наличия общего предка у
современных человекообраз-
ных обезьян и человека. Но
свидетельством общего про-
исхождения является сход-
ство в строении тела, пове-
дении и прежде всего в био-
химических веществах, из ко-
торых состоит организм.
Биомолекулярные исследо-
вания указывают на то, что
ближайшими из наших сов-
ременных родственников яв-
ляются крупные человекооб-
разные обезьяны Африки.
Кроме того, молекулярные
данные говорят о том, что
эволюционные линии челове-
ка и человекообразных
обезьян разошлись всего
лишь
5—8 миллионов лет
назад
— значительно ближе
к нашему времени, чем это
когда-то предполагалось.
На этих 16 гравюрах
изображены почти человеческие
позы и жесты, которые можно
наблюдать у современных
человекообразных обезьян
(иллюстрации взяты из книги
А. Брема «Жизнь животных»).
![]()
Крупные человекообразные обезьяны
Признаки орангутана.
а — Отсутствие хвоста.
б — Удлиненный таз.
в — Широкая и вместительная
грудная клетка.
г — Ноги короче рук.
д — Длинные кисти рук
с короткими большими пальцами.
противопоставленными прочим
пальцам.
![]() е — Костный гребень на черепе.
е — Костный гребень на черепе.
ж — Большой, по сравнению
с размерами тела, головной
мозг.
В число вымерших дриопитецинов и понгин, о
которых шла речь в предыдущей главе, несомненно,
входили предки человека и современных крупных
человекообразных обезьян - этих больших волоса-
тых умных обитателей тропических лесов Африки
и Юго-Восточной Азии. Ископаемые данные о пред-
ках крупных человекообразных обезьян немного-
численны, если не считать находок, позволяющих
связать орангутана с той группой ископаемых обезь-
ян, в которую входил Ramapithecus. Но в ходе био-
логических исследований было доказано, что у круп-
ных человекообразных обезьян и человека был не-
давний общий предок.
![]() По традиции зоологи объединяли крупных чело-
По традиции зоологи объединяли крупных чело-
векообразных обезьян - орангутана, шимпанзе и
гориллу -- в семейство Pongidae, а современного
человека вместе с ископаемым человеком и «обезья-
нолюдьми» помещали в другое семейство - Homi-
nidae. Однако на основании тщательного изучения
строения тела, образа жизни и молекулярной биоло-
гии большинство специалистов приходят сейчас к
убеждению, что человекообразные обезьяны и чело-
век относятся к одному семейству — Hominidae.
В его составе устанавливаются два подсемейства:
Ponginae, к которому относится орангутан, и Ho-
mininae, объединяющее гориллу, шимпанзе и чело-
века.
1. Pongo, орангутан, у него косматая рыжеватая
шерсть, длинные руки, сравнительно короткие ноги,
короткие большие пальцы на руках и ногах, крупные
коренные зубы с низкими коронками. Он питается
плодами, качается на ветвях деревьев, цепляясь за
них руками или ногами. Размеры — самцы бывают
ростом до 55 дюймов (1,4 м) и весят до 165 фунтов
(75 кг) — в два раза тяжелее самок. Время — совре-
менное. Место — Борнео (Калимантан), Суматра и
когда-то Юго-Восточный Китай. Подсемейство
Ponginae.
2. Pan, шимпанзе, у него длинная косматая чер-
ная шерсть, руки длиннее ног, лицо не покрыто
шерстью, с большими надглазничными валиками,
крупными выступающими ушами, плоским носом и
подвижными губами. Питается плодами, листьями,
семенами и мелкими животными; лазает по деревь-
ям, цепляясь руками и ногами, висит на ветках и
ходит по земле, опираясь на костяшки пальцев. Раз-
меры — до 5 футов (1,5 м), если выпрямится; масса
110 фунтов (50 кг). Время — современное. Место —
Экваториальная Африка. Подсемейство Homininae.
Триба Panini.
3. ![]() Gorilla, горилла, — крупнейшая из современ-
Gorilla, горилла, — крупнейшая из современ-
ных человекообразных обезьян. Самцы в два раза
крупнее самок, достигают роста 6 футов (1,8 м) и
массы 397 фунтов (180 кг). У этой сильной, плотно
сложенной обезьяны густая черная шерсть, голые
лицо и грудь, большие расширяющиеся ноздри и
сравнительно маленькие уши; у взрослых самцов
костный гребень на затылке и массивные челюсти.
Гориллы ходят, опираясь на костяшки пальцев; пи-
таются листьями, побегами, стеблями и корнями.
Время — современное. Место — Экваториальная
Африка. Подсемейство Homininae. Триба Panini.
|
|
![]()
![]()
О чем нам говорят
биохимические соединения.
Ниже на трех группах диаграмм
изображены биохимические
методы, при помощи которых
устанавливается, что крупные
человекообразные обезьяны
Африки являются нашими
ближайшими родственниками.
Молекулярные и другие исследования свидетель-
ствуют об удивительно близком родстве человека
и крупных человекообразных обезьян, особенно
африканской гориллы и шимпанзе.
Возьмем, например, нагревание ДНК. Ученые
смешивают отделенные друг от друга цепи двойных
спиралей, которые составляют ДНК, взятые от жи-
вотных, принадлежащих к различным видам. Сме-
шанные цепи соединяются, образуя гибридные двой-
ные спирали, но сочетаться могут лишь соответствую-
щие друг другу участки двух цепочек. После этого
с помощью нагревания, разделяющего цепочки, опре-
деляют теплостойкость каждой гибридной спирали.
Очевидно, что чем больше требуется тепла, тем боль-
ше имеется соответствующих друг другу участков и,
следовательно, тем теснее родство тех животных, от
которых были взяты цепочки ДНК. Этот опыт пока-
зывает, что ДНК шимпанзе и человека сходны на
99%.
Сравнение процентной доли различий в аминокис-
лотах, составляющих протеин, дает аналогичные
результаты. Исследование иммунологической реак-
ции приматов на антитела, образующиеся в орга-
низме кроликов в ответ на белковое вещество из чело-
веческой крови, подтверждает эти выводы.
![]()
![]()
|
|
|
|
|
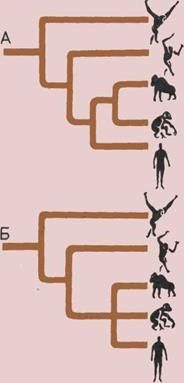
|
|
|
|
|
|
|
|
|
|
|
Сравнение анатомических особенностей убедитель-
но говорит в пользу того, что тело человека — не
что иное, как тело человекообразной обезьяны, спе-
циально приспособленное для хождения на двух но-
гах. Наши руки и плечи мало отличаются от рук
и плеч шимпанзе. Однако, в отличие от человекооб-
разных обезьян, ноги у нас длиннее рук, а таз, позво-
ночник, бедра, ноги, ступни и пальцы ног подверг-
лись изменениям, которые позволили нам стоять и
ходить, держа тело в вертикальном положении.
(Крупные человекообразные обезьяны могут стоять
на двух ногах, только согнув колени, и ходить на но-
гах, пошатываясь из стороны в сторону.)
Приспособление ног к новой функции означало,
что мы уже не могли использовать большие пальцы
ног подобно большим пальцам рук. Большие пальцы
на наших руках сравнительно длиннее, чем у чело-
векообразных обезьян, и могут, согнувшись над
ладонью, прикоснуться своими кончиками к кончи-
кам других пальцев, что обеспечивает точность хва-
тания, которая нам нужна при изготовлении и ис-
пользовании орудий.
Хождение на двух ногах, более развитый интеллект
и разнообразная пища — все это способствовало
появлению различий черепа, мозга, челюстей и зубов
у человека и обезьян. Сравнительно с размерами
тела мозг и черепная коробка человека значительно
больше, чем у обезьяны; кроме того, мозг человека
более высокоорганизован, а его сравнительно более
крупные лобные, теменные и височные доли совмест-
но осуществляют функции мышления, управления
общественным поведением и человеческой речью.
Челюсти современного всеядного человека зна-
чительно короче и слабее, чем у человекообразный
обезьян, которые придерживаются в основном веге-
тарианской диеты. У обезьян есть амортизирующие
удары надглазничные валики и костные черепные
гребни, к которвш прикреплены мощные челюстные
мышцы. У человека отсутствуют толстые шейные
мышцы, которые у взрослый обезьян поддерживают
выступающую вперед морду. Ряды наших зубов рас-
положены в виде параболы, отличаясь этим от распо-
ложенный в виде латинской буквы U зубнык рядов
человекообразных обезьян; кроме того, клыжи обезь-
ян значительно крупнее, а коронки коренных зубов
значительно выше, чем у нас. Но зато коренные зубы
человека покрыты более толстым слоем эмали, что
делает их более износоустойчивыми и позволяет пе-
режевывать более твердую пищу.
![]()
![]()
![]()
![]()
![]()
![]() Различия в строении языка и глотки у человека
Различия в строении языка и глотки у человека
и шимпанзе позволяют нам издавать более разно-
образные звуки, хотя как у человека, так и у шим-
панзе черты лица могут принимать различные вы-
ражения.
![]()
![]() В следующих главах мы покажем, что некоторые
В следующих главах мы покажем, что некоторые
особенности, присущие человекообразным обезья-
нам, сохранились и у ранних представителей
человеческого рода.
мозговой коробкой, вертикально
сбалансированный
на позвоночнике.
б - Небольшие челюсти.
маленькие зубы, покрытые
толстым слоем эмали, коренные
чубы с низкой коронкой;
зубная дуга, имеющая форму
параболы.
в — Длинные пальцы руки,
позволяющие точно захватывать
мелкие предметы.
г — Короткая поясница.
д — Широкий короткий таз.
е — Ноги длиннее рук.
ж — Большой пален ноги
расположен параллельно прочим
пальцам и помогает во время
ходьбы переносить вес тела.
Б. Особенности анатомии
гориллы.
а — Череп с выступающей
вперед удлиненной лицевой
частью расположен спереди
от спинного хребта и имеет
костный гребень,
поддерживающий мощную
челюсть и мышцы шеи.
Сравнение головного мозга.
А — Головной мозг человека.
Б — Головной мозг шимпанзе.
а — Лобная доля (различные
функции).
б — Центр Брока (центр речи),
у человекообразных обезьян
развит слабо.
в — Теменная доля
(деятельность органов чувств
и др.).
г — Височная доля (намять
и др).
![]() д — Затылочная доля (зрение).
д — Затылочная доля (зрение).
е — Мозжечок (координация).
б — Массивные челюсти
с большими клыками, крупными
коренными зубами, которые
имеют высокие коронки и тонкий
слой эмали, и с зубными дугами
в форме латинской буквы U.
в — Короткий большой палец
и длинные прочие пальцы рук.
г — Поясница сравнительно
более короткая, чем
у человека.
д — Удлиненный таз.
е — Ноги короче рук.
ж — Отставленный в сторону
большой палец ступни
приспособлен для хватания.
Человек — родич обезьяны
Внушительные косвенные данные о наличии об-
щего предка у человека и крупных человекообразных
обезьян дает нам неотения — сохранение во взрос-
лом возрасте тех особенностей, которые у других
видов встречаются только среди детенышей. Так,
у взрослых людей имеются многочисленные харак-
терные черты, которые есть у детенышей шимпан-
зе, но утрачиваются ими в зрелом возрасте.
Как и у людей, у детенышей шимпанзе тело по-
крыто редкими волосами. Как и у нас, у них сравни-
тельно крупный мозг, защищенный выпуклым чере-
пом. Кости их черепа, как и нашего, тонкие и не
имеют ярко выраженных надглазничных валиков и
затылочных гребней. Лицо у них еще не развилось,
и поэтому оно не выступает вперед; челюсти и зубы
небольшие, но подбородок выдвинут вперед. У де-
тенышей шимпанзе, как и у человека, мозговой ствол
соединяется с головным мозгом через отверстие,
расположенное под средней частью черепа, и таким
образом при ходьбе на двух ногах череп находится
в равновесии над спинным хребтом. Как и у жен-
щин, влагалище юных самок шимпанзе направлено
вперед, а не назад.
У человека и шимпанзе продолжительное детство,
что обеспечивает преимущество существам, поведе-
ние которых основано не столько на инстинкте,
сколько на навыках, полученных при обучении. Столь
длительное детство человека приводит к тому, что
продолжительность его жизни дольше, чем у любого
из прочих млекопитающих.
Таким образом, крупный мозг, маленькие челю-
сти и прямостояние — все эти наиболее характерные
черты, которые, как считается, отличают нас от
обезьян, являются, по крайней мере частично, ре-
зультатом неотении.
![]() Механизмы, превратившие обезьян в человека,
Механизмы, превратившие обезьян в человека,
вероятно, состояли просто в изменении наборов
генов, которые то подключались, то отключались,
регулируя темпы и величину роста различных си-
стем организма. Эти изменения в основном влияют на
организм через такие гормоны, как соматотрофин,
стимулирующий рост костей, и мелатонин, который
способствует достижению половой зрелости. В ходе
естественного отбора во взрослом возрасте просто
сохранялись те детские черты, которые спбсобство-
вали выживанию наших предков.
Развитие черепа.
Очень похожие у зародышей
черепа шимпанзе и человека
в дальнейшем развиваются
совершенно по-разному.
Искажение линий сетки
координат показывает те части
черепа, которые с возрастом
подвергаются наиболее сильным
изменениям.
1. Череп шимпанзе.
а — Череп зародыша.
б — Череп детеныша.
в — Череп взрослого шимпанзе.
2. Череп человека.
![]()
![]()
![]() г — Череп зародыша.
г — Череп зародыша.
д — Череп взрослого человека.
![]()
|
Выражение лица.
Хотя шимпанзе не обладают
даром речи, они могут выражать
свои эмоции мимикой, которая
очень похожа на человеческую.
На наших рисунках
представлены четыре выражения
лица шимпанзе.
1. Спокойствие.
2. Приветствие.
3.
Улыбка, видны только нижние
зубы.
4.
![]() Гнев, обнажаются как
Гнев, обнажаются как
верхние, так и нижние зубы.
В образе жизни обезьян-приматов также прояв-
ляется сходство между ними и человеком. Как низ-
шие, так и человекообразные обезьяны живут со-
циальными группами, которые подчиняются прави-
лам, связанным с возрастом, полом и определенной
иерархией. В отличие от групп других животных
(скажем, птиц), группы обезьян большей частью
постоянны, охватывают представителей обоих по-
лов и ведут себя следуя не инстинктам, а приобре-
тенным навыкам поведения.
Ближе всего к человеку в этом отношении стоят
шимпанзе — умньге, в высшей степени общественные
животные, живущие стадами. Ядро стада состоит
из групп самцов с молодыми самками, которых
набирают со стороны.
Шимпанзе испытывают сильную прочную при-
вязанность друг к другу и общаются между собой,
пользуясь разнообразной мимикой, жестами, по-
зами и звуками. Хотя у шимпанзе отсутствуют точ-
ные эквиваленты звуковой речи, они могут привет-
ствовать и подбадривать друг друга, проявлять
почтительность или агрессивность, почти как че-
ловек.
Некоторые шимпанзе изготовляют орудия; когда-
то считалось, что эта способность присуща только
человеку. Шимпанзе, в особенности самки, питаются
термитами, пойманными при помощи веточек, за-
остренных так, чтобы ими можно было расковыри-
вать ходы в термитниках. Отдельные шимпанзе
высасывают воду из губок, сделанных из листьев;
пользуются палками как рычагами; замахиваются
дубинками на врагов.
В некоторых группах самцы помогают друг другу
при охоте на низших обезьян, а затем делят между
собой их мясо. И хотя шимпанзе не ведут между
собой междоусобных войн, известен случай, когда
![]()
![]() Шш
1
Шш
1
u а
'QLIч
одна группа самцов устроила набег на другую и пере-
била ее членов. Правда, возможно, это было вызва-
но тем, что соседние фермеры вырубили часть леса,
где жили шимпанзе, что привело к перенаселенности.
![]() Следовательно, говоря вкратце, шимпанзе обра-
Следовательно, говоря вкратце, шимпанзе обра-
зуют группы со сравнительно постоянным составом,
обитающие в защищенных от нападения участках
леса, где их члены собирают пищу, охотятся, размно-
жаются и иногда изготовляют простые орудия. Если
добавить к этому хождение на двух ногах и зависи-
мость от применения орудий, то мы можем пред-
ставить себе образ жизни тех доисторических су-
ществ, которые перешли черту, отделявшую нашего
предка обезьяну от раннего человека.
Семейная жизнь.
Как и люди, шимпанзе
испытывают чувства сильной
привязанности друг к другу.
Родительские отношения матери
к ребенку помогают детенышам
во время длительного периода их
обучения. Необходимость
защиты молодняка способствует
укреплению
![]() внутригрушювых связей
внутригрушювых связей
в каждом стаде шимпанзе.
![]() «ЧЕЛОВЕКООБЕЗЬЯНЫ»
«ЧЕЛОВЕКООБЕЗЬЯНЫ»
И РАННИЕ ЛЮДИ
![]() Около
4 миллионов лет назад
Около
4 миллионов лет назад
от человекообразных обезьян
в Африке произошли гоми-
ниды, которые ходили только
на двух ногах. Хождение
на двух ногах способствовало
координации движений рук
и глаз и развитию головного
мозга. Так возникла чело-
веческая триба
Hominini.
К ее старейшим представи-
телям, очевидно, относился
род
Australopithecus.
Мно-
гие ученые считают, что от
одной из таких «человеко-
обезьян» произошел малень-
кий
Homo habitis
— первый
из известных нам видов на-
шего рода
Homo
(«чело-
век»). В этой главе рассмат-
риваются строение тела и
образ жизни этих существ,
живших более миллиона лет
назад или даже еще раньше.
На этой картине-реконструкции
художника Мориса Уилсона
показано столкновение между
древнейшими людьми
и австралопитеками. Место
действия — Олдувай
в Восточной Африке; время —
около 2 миллионов лет назад.
![]()
Климат и гоминиды.
Основные этапы эволюции
человека, по-видимому,
отражают климатические
изменения, вызванные
событиями эпох оледенений
(шкала дана в миллионах лет).
а — Предки человекообразных
обезьян.
б — Ранние австралопитеки.
в — Поздние австралопитеки.
г — Виды рода Homo.
А — Образование
антарктической ледовой шапки.
Б — Образование арктической
ледовой шапки.
В — Чередование оледенений
и межледниковых периодов.
Первые отпечатки следов ног.
1.
Ископаемый след ноги из
Лаэтоли в Танзании. Он остался
в свежевыпавшем мягком
вулканическом пепле, который
вскоре после этого превратился
в твердую породу.
2.
Очертания ископаемого следа
ноги.
3.
След ноги современного
человека.
![]() По рисункам 2 и 3 можно
По рисункам 2 и 3 можно
заключить, что эти следы
оставлены существами,
сходными по размерам и по весу.
4.
![]() Следы трех прямоходящих
Следы трех прямоходящих
гоминид, совершавших
совместную прогулку более
3,5 миллиона лет назад.
Три с половиной миллиона лет назад все усили-
вающийся холод ледникового периода сковал так
много воды в виде льда, что количество дождевых
осадков в мире резко уменьшилось и тропические
саванны начали сменять леса, площадь которых
сократилась. В Африке стали размножаться круп-
ные травоядные и другие животные, приспособлен-
ные к жизни в саванне, в то время как обитатели
лесов пострадали, лишившись своей естественной
среды обитания.
Некоторые существа, произошедшие от дриопи-
тецинов, но не вполне приспособленные к жизни
на деревьях, по-видимому, начали приноравливать-
ся к существованию в открытой местности. Этими
«человекообезьянами» были австралопитеки (о них
пойдет речь дальше в этой главе) и первый «насто-
ящий человек», вероятно, произошедший от австра-
лопитеков. Гоминизация (становление человека),
по-видимому, была связана с совокупностью из-
менений, которые взаимно подкрепляли друг друга.
О некоторых из них можно судить по ископае-
мым находкам, о других можно только догадываться.
![]()
![]()
![]()
![]() Ископаемые отпечатки стоп возрастом 3,8 миллио-
Ископаемые отпечатки стоп возрастом 3,8 миллио-
на лет говорят о том, что к этому времени хождение
на двух ногах освободило руки. Это позволило перей-
ти к производству и регулярному использованию
орудий, самые ранние из которых имеют возраст
2,5 миллиона лет. Размеры клыков уменьшались,
|
|
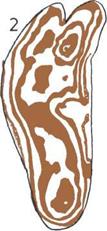
вероятно, вследствие того, что орудия все больше
заменяли эти зубы, выполняя их задачи. Изготовле-
ние орудий и прямохождение стимулировали раз-
витие головного мозга и привели к новой стратегии
поведения, направленной на выживание, которая
состояла в заготовке растительной пищи и дичи.
Со временем некоторые из гомининов утратили воло-
сяной покров на теле, и у них появились потовые
железы, а эти изменения помогли бороться с пере-
гревом тела на жарких, лишенных тени равнинах.
![]() Охота на крупную дичь, дающую много мяса, или
Охота на крупную дичь, дающую много мяса, или
поедание трупов животных, убитых хищниками, при-
вели к тому, что целые группы гомининов смогли
теперь делиться запасами пищи на «домашних ба-
зах», где их совместная деятельность способствовала
взаимному общению, которое предшествовало появ-
лению речи. Здесь молодежь училась у старших из-
готовлению и использованию орудий. Впервые в исто-
рии эволюции преимущество получил интеллект: те-
перь уже от мозга, а не от мускульной силы или ско-
рости ног зависело, какие из гомининов выживут, а
какие вымрут.
Процессы с обратной связью.
Изменения в окружающей
среде, возможно,
способствовали эволюции
гоминид, послужив началом
ряда процессов, которые в свою
очередь взаимно влияли друг
на друга.
а — Групповое сотрудничество
помогало выжить в условиях
открытой саванны.
б — Наличие домашних стоянок,
где проводился раздел пищи,
скрепляло общественные связи.
в — Изготовление
и применение орудий развивало
координацию зрения и ручных
операций, помогало в собирании
пищи и в охоте, стимулировало
ходьбу на двух ногах.
г — Усложнение мозга облегчало
общение и повышало
способность к обучению при
изготовлении и использовании
новых орудий.
![]()
![]() д — Удлинение сроков ухода
д — Удлинение сроков ухода
за детьми и их обучения
способствовало выживанию
гоминид.
Увеличение размеров мозга, изменение зубов и
приспособление бедер и нижних конечностей к хож-
дению — таковы основные признаки, отличающие
человеческую трибу Hominini. В нее входят, по край-
ней мере, два рода: вымерший Australopithecus
(«южная обезьяна») и Homo («человек»). Некото-
рые ученые добавляют к ним и род Ramapithecus
(описанный на с. 82—83).
Австралопитеки, так называемые «обезьянолюди»,
появились в Африке 4 миллиона лет назад или даже
раньше. Они, очевидно, произошли от дриопитеци-
нового предка человекообразных обезьян, которого
еще предстоит найти. Всего возникло три или четыре
вида (мнения специалистов расходятся). Два мил
лиона лет назад от одного из них, вероятно, произо-
шел первый вид, относящийся к нашему роду (см.
с. 110). Оба рода (Australopithecus и Homo) суще-
ствовали бок о бок на протяжении еще 1 миллиона
лет, пока не вымерли последние австралопитеки,
которые, возможно, были истреблены своими более
умными преемниками.
Вполне вероятно, что тело австралопитеков было
покрыто волосами; одни из них были хрупкого тело-
сложения и не крупнее шимпанзе, а другие имели
мускулистое тело и по размерам приближались к
современному человеку. Соотношение величины
мозга и тела было не намного лучше, чем у челове-
кообразных обезьян, а обезьяноподобная голова
имела вогнутую лицевую часть, плоский нос и сре-
занный подбородок. Челюсти и коренные зубы были
большими и мощными, причем зубные дуги имели
форму латинской буквы V или заостренной U. Но
в отличие от человекообразных обезьян, нижние
конечности австралопитека были длиннее рук и слу-
жили прежде всего для прямохождения.
Австралопитеки, очевидно, приввжли пользоваться
руками для переноски грузов и изготовления камен-
ных орудий, которыми можно бвио резать мясо,
хотя их зубы бвии пригодны для дробления зерен и
разжевывания листьев.
Собирая плоды, питаясь трупами убитвк хищни-
ками животных, а иногда, возможно, даже охотясь
на них, небольшие группы австралопитеков бро-
дили по тропическим пастбищам Восточной и Южной
Африки. Некоторые из них, может быть, переходили
также в Азию и Европу.
![]()
![]()
![]()
![]()
![]()
![]()
![]() Следующие восемь страниц посвящены более
Следующие восемь страниц посвящены более
подробному описанию четырех видов этих первобыт-
ных гомининов.
д — Характерный изгиб
позвоночника в форме буквы S.
е — Короткий широкий таз.
ж — Длинная берцовая кость,
направленная под углом внутрь,
з — Коленный сустав с плоской
поверхностью.
и — Длинные нижние
конечности.
к — Стопа ноги в виде
«платформы».
Взгляды на
|ю.т
ословную
человека.
Ра личные пре (положения
о происхож дении
австралопитека, человека
и африканских
человекообразных обезьян
представлены в виде двух
альтернативных эволюционных
|рев. Имеются значительные
разногласия в оценке
длительности существования
некоторых видов. Пунктиром
пока заны линии, ведущие от
неизвестных предков (шкала
времени дана в миллионах лет).
А — Человек произошел
от австралопитека.
Б — Человек произошел не от
австралопитека.
а — Австралопитек из Афара
(Australopithecus ajarensis).
б — Австралопитек африканский
(Australopithecus africanus).
в — Австралопитек могучий
(Australopithecus robuslus).
г — Австралопитек Бойса
(Australopithecus boisei).
д — Человек умелый (Homo
habilis).
е — Человек прямоходящий
(Homo erect us).
ж — Человек разумный (Homo
sapiens).
з — Африканские
![]()
![]()
![]()
![]() человекообра зные обезьяны.
человекообра зные обезьяны.
| •U 1 е 1 |
«1 г б |
L |
3 1 • |
||
| , |
1 |
1 |
j |
а |
• д • |
| 1 3 |
• Ч • &L |
1 • • |
• |
||
| U а | |
• |
• |
|||
| • 5 • |
•• |
• • |
|||
| • А • |
Б |
• • • |
|||
|
|
|
|
|
|
|
|
зе, и, вероятно, еще не мог управлять речью. Лицо
было таким, как у человекообразных обезьян: с низ-
ким лбом, надглазничным валиком, плоским носом,
срезанным подбородком, но выступающими вперед
челюстями с массивными коренными зубами. Перед-
ние зубы имели щербины, очевидно, потому, что ими
пользовались как орудиями для хватания.
Австралопитек из Афара ходил на слегка согну-
тых ногах, а его бедра, несколько похожие на бедра
шимпанзе, и изогнутые кости пальцев рук и ног сви-
детельствуют о том, что он проводил много времени
на деревьях и, возможно, спал высоко среди ветвей
вне пределов досягаемости хищников. Самки имели
значительно более близко поставленнвге бедра и,
следовательно, более узкие родовые пути, чем у со-
временный: женщин, а поэтому они, должно быть,
рожали детенышей со значительно меньшими голов-
ками и объемом мозга, чем у современный новорож-
денный: человеческих младенцев.
Семейные группы занимались сбором растительной
пищи, в которую входили жесткие, твердые или во-
локнистые семена и плоды. Некоторые отдельные
особи, возможно, изготавливали грубые орудия из
дерева и камня, чтобы отделять мясо от костей жи-
вотных, убтых хищниками.
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]() Почти нет сомнения, что до того, как афарские
Почти нет сомнения, что до того, как афарские
австралопитеки выгмерли 2,5 миллиона лет назад,
от них прямо или косвенно произошли другие австра-
лопитецины и наш род Homo.
Australopithecus africanus («африканская южная
обезьяна») расселился на Земле приблизительно
3 миллиона лет назад и прекратил свое существо-
вание около миллиона лет до наших дней. Он, веро-
ятно, произошел от Australopithecus afarensis, и не-
которые авторы полушутя высказывают предполо-
жение, что он явился предком шимпанзе.
Маленький, хрупко сложенный africanus, похожий
на человекообразную обезьяну, был ростом с неболь-
шого африканского бушмена, а весил не больше 12-
летней европейской девочки. Он ходил выпрямив-
шись, хотя мускулы его ног и отличались от наших.
Руки были сравнительно длинные, а их большие и
прочие пальцы, вероятно, менее умело обращались
с предметами, чем это получается у нас.
![]() Нижняя часть его лица выступает вперед, но не
Нижняя часть его лица выступает вперед, но не
столь значительно, как у человекообразных обезьян.
На некоторых черепах заметны следы затылочного
гребня, к которому были прикреплены сильные мыш-
цы шеи. Мозг был не больше, чем у гориллы, но слеп-
ки свидетельствуют о том, что по строению мозг не-
сколько отличался от мозга человекообразных обе-
зьян. По сравнительному соотношению размеров
мозга и тела африканус занимает промежуточное по-
ложение между современными человекообразными
обезьянами и людьми.
Телосложение.
На этом рисунке австралопитек
африканский для сравнения
изображен рядом с современным
человеком. Рост 3—4 фута
(1 — 13 м). Масса 45—90 фунтов
(20—40 кг).
Сравнение зубных дуг.
![]() А — Крупная зубная дуга
А — Крупная зубная дуга
с очень большими коренными
зубами, принадлежащая
австралопитеку африканскому.
Б — Меньшая зубная дуга
с более мелкими зубами,
принадлежащая современному
человеку.
|
|
Ученые расходятся во мнении относительно того,
какой образ жизни вел африканус. Строение зубов
и челюстей позволяет думать, что этот обезьяноче-
ловек пережевывал растительную пищу, но, воз-
можно, также обгладывал мясо животных, убитых
хищниками. Специалисты оспаривают его способ-
ность к изготовлению орудий: большинство костя-
ных «орудий», обнаруженных около ископаемых
остатков африкануса, оказались просто остатками
пищи гиен или других хищников.
Некоторые авторы предполагают, что африканус—
не самостоятельный вид, а попросту самки гоминид,
которые должны быть отнесены к другому виду —
Australopithecus robustus. Однако представители
этого вида явно относятся к более позднему вре-
мени, чем большинство известных нам экземпляров
африкануса. Как считают некоторые специалисты,
самой древней находкой африкануса является обло-
мок челюсти, датируемый 5,5 миллиона лет, из
Лотегама в Кении, а самый молодой экземпляр
имеет возраст 700 тысяч лет. Но нельзя с полной
уверенностью утверждать, что эти австралопитеки
действительно относятся к виду африканус.
![]() Большинство ископаемых находок обнаружены в
Большинство ископаемых находок обнаружены в
пещере Стеркфонтейн в Южной Африке. Другие
находки говорят о том, что африканус обитал также
в Эфиопии, Кении и Танзании.
![]()
р.Нил
оз. 1 уркаид
оз. Виктора я
р.Лимпопо
5*
![]()
![]()
![]() g •Претория
g •Претория
Австралопитек могучий (Australopithecus ro-
bustus), которого одно время называли парантро-
пусом (Paranthropus — «около-человек»), был
более крупным и лучше развитым физически, не-
жели австралопитек африканский. Как мы уже гово-
рили, некоторые авторы считают, что обе эти «юж-
ные обезьяны» являются соответственно самцами
и самками одного и того же вида, но большинство
специалистов не поддерживают это предположение.
Австралопитек могучий был такого же роста, как
многие европейские женщины — 1,6 м, но весил
меньше, чем большинство из них, — около 50 кг.
По сравнению с африканусом у него был более круп-
ный и плоский череп, вмещавший более крупный
мозг — около 550 см3
, и более широкое лицо. К вы-
сокому черепному гребню были прикреплены мощ-
ные мышцы, приводившие в движение массивные
челюсти. Передние зубы были такими же, как у
африкануса, а коренные — более крупными. При
этом коренные зубы у большинства известных нам
экземпляров обычно сильно изношены, несмотря на
то, что они были покрыты толстым слоем прочной
эмали. Это может свидетельствовать о том, что жи-
вотные питались твердой, жесткой пищей, в частно-
сти зернами злаков.
![]() По всей видимости, австралопитек могучий поя-
По всей видимости, австралопитек могучий поя-
вился около 2,5 миллиона лет назад. Все останки
представителей этого вида найдены в Южной Аф-
рике, в пещерах, куда их, вероятно, затащили хищ-
ные звери. Вымер этот вид около 1,5 миллиона лет
назад. От него, возможно, произошел австралопитек
Бойса (см. с. 108—109). А строение черепа австра-
лопитека могучего позволяет предположить, что
он был также предком гориллы.
Телосложение.
На этом рисунке австралопитек
могучий показан для сравнения
рядом с современным
человеком. Рост 4 фута
11 дюймов — 5 футов
7 дюймов (1,5—1,7 м).
Зубная дуга.
![]() Судя по массивным коренным
Судя по массивным коренным
зубам и маленьким резцам,
австралопитек могучий был
травоядным.
Находки в расщелинах.
Найденные в расщелинах
известняка кости
австралопитека могучего,
вероятно, оказались там
следующим путем.
|
|
Места, где обитал
австралопитек могучий.
1. Кромдраай.
2. Сварткранс.
![]() Многие специалисты считают.
Многие специалисты считают.
что местами обитания этого
австралопитека были также
стоянки, перечисленные на с. 109.
![]()
![]() А — Уровень дневной
А — Уровень дневной
поверхности в древности.
Б — Современный уровень
дневной поверхности.
а — Леопард тащит труп
австралопитека вверх на дерево,
где его не смогут достать гиены.
б — Кости австралопитека
падают в расщелину
известняков.
в — Кости захораниваются под
слоями осадков.
а — Отверстия в черепе
австралопитека могучего.
б — Размеры отверстий
соответствуют величине клыков
леопарда.
Сравнение черепов
(внизу
справа).
![]() А — На изображенном
А — На изображенном
с поворотом в три четверти черепе
Australopithecus robustus видны
костный гребень и массивные
челюсти. Объем черепа 500 см3
.
Б — Изображение черепа Homo
sapiens sapiens с поворотом
в три четверти. Объем мозга
1400 см3
.
Этот наиболее крупный и сильный из всех австра-
лопитеков получил свое название в честь английского
бизнесмена Чарльза Бойса, помогавшего финанси-
ровать палеонтологические исследования в Восточ-
ной Африке, в результате которых в 1959 г. был
найден неизвестный ранее науке вид «южных обезь-
ян». Первоначально Australopithecus boisel имено-
вался Zinjanthropus, что означает «восточно-афри-
канский человек». Время его существования — от
2,5 до 1 миллиона лет назад.
Некоторые палеоантропологи считают, что австра-
лопитек Бойса не является самостоятельным видом,
а представляет собой лишь местную разновидность
австралопитека могучего. Тем не менее для него
характерны многие отличительные признаки. Судя
по реконструкциям, сделанным на основании изу-
чения ископаемых костей черепа и конечностей, неко-
торые экземпляры этого вида достигали роста совре-
менного человека. Мозг у них был таких же разме-
ров, как у австралопитека могучего, т. е. примерно
втрое меньше нашего мозга. Эти существа ходили
прямо. Своим мощным телосложением они напоми-
нали гориллу. Как и у горилл, самцы были, по-види-
мому, значительно крупнее самок.
![]()
![]() Подобно горилле, австралопитек Бойса имел
Подобно горилле, австралопитек Бойса имел
большой череп с надглазничными валиками и цент-
ральным костным гребнем, служившим для при-
крепления мощных челюстных мышц. Но по сравне-
нию с гориллой, гребень у австралопитека Бойса
был меньше и более выдвинут вперед, лицо — более
плоское, а клыки — менее развитые. Из-за огром-
ных коренных и предкоренных зубов это животное
получило прозвище «щелкунчик». Но биомеханиче-
ские исследования показывают, что эти зубы не могли
Телосложение.
Крупный австралопитек Бойса
показан рядом с современным
человеком. Рост 5 футов
3 дюйма — 5 футов
Юдюймов (1,6—1,78 м), но
мог быть и меньше. Масса
132-176 фунтов (60—80 кг).
Зубная дуга.
На этой неполной зубной дуге
(показанной в уменьшенном
масштабе) заметны признаки,
характерные для вида
Australopithecus boisei.
а — Маленькие резцы,
предназначенные для
откусывания.
![]() б — Огромные коренные зубы,
б — Огромные коренные зубы,
способные размалывать пищу.
оказывать сильного давления на пищу и были при-
способлены для пережевывания не очень жесткого
материала, например листьев.
![]()
![]() Поскольку вместе с костями австралопитека Бой-
Поскольку вместе с костями австралопитека Бой-
са, возраст которых составляет 1,8 миллиона лет,
была найдена оббитая галька, можно предполо-
жить, что эти существа могли использовать камень
в практических целях. Однако не исключено, что
представители данного вида попросту пали жертвой
своего современника — человека умелого, преуспев-
шего в применении каменных орудий (см. с. 110—
111).
Четыре изображения черепов.
1. Череп самца Australopithecus
boisei.
а — Сагиттальный
(стреловидный) гребень.
б — Реконструкция нижней
челюсти.
2. Череп, принадлежавший,
вероятно, самке, которая была
значительно меньших размеров.
3. Боковой вид черепа самца.
в — Надглазничный валик.
4. Вид черепа сверху.
Места, где обитал австралопитек
Бойса.
А — Африка (район находок
отмечен четырехугольником).
Б — Район находок, данный
с увеличением.
1. Омо.
2. Кооби Фора.
3. Олдувайское ущелье.
4. Пениндж.
5.
![]()
![]() Чесованджа.
Чесованджа.
Ступня ноги и кисть руки
(вверху
слева).
Судя по найденным костям
ступни и кисти (на рисунке они
окрашены), человек умелый
ходил на двух ногах, а пальцы
его кистей имели сильную
и точную хватку.
Телосложение.
Человек умелый показан для
сравнения рядом с современным
человеком. Рост 4—5 футов
(1,2— 1,5 м). Масса — около
110 фунтов (50 кг).
Зубная дуга.
![]()
![]()
![]()
![]() Изгиб дуги имеет более
Изгиб дуги имеет более
«современный» вид, а коренные
зубы несколько уже, чем
у австралопитека.
Человек умелый жил в Восточной Африке и, воз-
можно, в Южной Африке ("Telanthropus") и в
Юго-Восточной Азии ("Meganthropus").
Остатки материальной культуры, найденные около
костей человека умелого, позволяют думать, что
эти существа занимались изготовлением каменных
орудий, строили несложные укрытия, собирали расти-
тельную пищу, отрубали крупные куски мяса от
туш животных, убитых хищниками, и охотились на
малую, а может быть, и на достаточно крупную
дичь.
![]() От Homo habilis,
вероятно, произошел Homo
От Homo habilis,
вероятно, произошел Homo
ereclus.
|
|
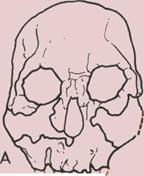
Три виза черепа.
У человека умелого голова
была округлее, мозг крупнее,
а лицевая часть меньше, уже
и длиннее, чем у австралопитека,
однако она имела и некоторые
архаичные черты. На рисунках
отсутствует нижняя челюсть.
Недостающие фрагменты черепа
показаны штрихами.
А — Вид спереди.
Б — Вид сбоку.
В — Вид сверху.
Места, где обитал человек
умелый.
1. Кооби Фора.
2. Олдувайское ущелье.
3.
![]() Сварткранс (возможно).
Сварткранс (возможно).
Некоторые ученые относят
к Homo habilis находки,
сделанные и в других районах
Восточной Африки.
![]()
|
Ill
Орудия из Олдувая.
Ранние каменные орудия из
Олдувайского ущелья
в Танзании.
А — Грубое рубило (чоппер)
из лавы; им пользовались для
резки мяса или для
раскалывания костей.
Б — Многогранник (полиэдр)
с тремя или с большим числом
режущих краев.
В — Дискоид с острыми краями.
Г — Скребло для обработки
шкур.
![]() Д — Каменный молоток.
Д — Каменный молоток.
Ранние следы
материальной культуры
Ранний человек был медлительнее и слабее, чем
крупные хищники, и не имел такого естественного
оружия, как клыки и когти. И все-таки ранние го-
минины научились компенсировать эти недостатки.
Они стали придавать кускам камня, костей и дерева
форму, позволяющую резать, скрести и копать. В
отличие от клыков и когтей, такие орудия можно
было собирать, хранить или по своему желанию
менять одно на другое. Со временем эти орудия
дали человеку беспрецедентную власть над окружаю-
щей его средой.
Первыми орудиями, вероятно, были осколки ко-
стей, острые палки и подносы из коры для сбора
пищи. Такие, в основном непрочные, орудия не со-
хранились. Но камень оказался долговечнее. Мы
знаем, что ранние гоминины в Эфиопии сознательно
расщепляли небольшие камни, возможно, для того,
чтобы получить твердые, острые края для резки мяса.
С того времени (2,5 миллиона лет назад) примерно
до 5 тысяч лет назад камень доминировал в чело-
веческой технике.
Орудия палеолита, или древнекаменного века,
можно найти повсюду, куда только ступала нога
человека примерно до 10 тысяч лет назад. Мастера
раннего палеолита научились создавать острые ору-
дия, сильно ударяя камнем о камень. Они отбирали
только те камни, которые легко расщеплялись при
ударе и именно вдоль тех трещин, которые были
нужны мастеру. Выбирались достаточно твердые
горные породы, которыми можно было резать, дро-
бить, расщеплять или скрести растительные и жи-
вотные материалы.
![]() Кремень, кремнистый сланец (черт), кварцит
Кремень, кремнистый сланец (черт), кварцит
и горный хрусталь оказались наиболее пригодными
для этого горными породами, но ни одна из них не
встречается повсюду. В Восточной Африке самые
древние орудия изготовлялись из лавы, которая
дает более грубую поверхность, чем кремень.
Наиболее известными древними инструментами
являются типичные орудия «олдованской» культуры
из Олдувайского ущелья в Танзании. Здесь 1,9 мил-
лиона лет назад Homo habilis (или какой-либо из
современных ему австралопитеков) расщеплял ба-
зальт и кварцитовую гальку, придавал им формы,
которые сейчас называют грубыми рубилами (чоп-
перами), скреблами, резцами, топоровидными ору-
диями, а по их очертаниям делят на дискоиды, мно-
гогранники (полиэдры) или субсфероиды.
Некоторые ученые считают, что на самом деле
истинными орудиями были не перечисленные формы,
а так называемые «бросовые» осколки, отщеплен-
ные от этих ядер. Без сомнения, 1,5 миллиона лет
назад около озера Туркана в Кении гоминины исполь-
зовали маленькие каменные отщепы для того, чтобы
разделывать туши антилоп. Царапины от зубов, на-
ходящиеся на костях животных под царапинами от
режущих орудий, наводят на мысль, что ранние
гоминины обычно соскабливали мясо с туш живот-
ных, убитых хищниками. Но вполне вероятно, что
ранние люди изготовляли также и орудия для охоты.
Каменные шары из Олдувая, связанные ремнями,
превращались в охотничий снаряд, метнув который,
можно было сбить с ног антилопу, подобно тому как
в наши дни ковбои ловят скот с помощью метатель-
ных шаров-бола.
![]()
![]() Олдованская индустрия (называемая также ру-
Олдованская индустрия (называемая также ру-
било-осколочной или галечной) и ее более поздние
варианты широко распространились по значитель-
ной части Африки и Евразии раннего каменного века.
В некоторых местах эта культура процветала еще
200 тысяч лет назад — уже много лет спустя после
изобретения более совершенных технологических
приемов каменного века.
Олоувайская временная шкала.
1.
Слой № I: 2,0—1,7 миллиона
лет назад.
2.
Слой № 2: 1,7—1,1 миллиона
лет назад.
3.
Слой № 3: 1,1—0.8 миллиона
лет назад.
4.
Слой № 4: 0,8—0,6 миллиона
лет назад.
а — Типичные олдувайские
орудия.
![]()
![]() б — Более совершенные
б — Более совершенные
и разнообразные орудия.
частично похожие на орудия,
характерные для человека
прямоходящего.
в — Австралопитеки.
г — Человек умелый.
![]()
![]() Древнейшая хижина? (справа).
Древнейшая хижина? (справа).
Возраст этого нагромождения
камней и ископаемых остатков
1,8 миллиона лет, и оно,
возможно, представляет собой
следы древнейшего из известных
нам человеческих жилищ. Камни,
вероятно, прижимали к земле
ветки, составлявшие каркас
хижины на берегу озера
в Олдувае.
Древнее жилище (внизу).
Эта реконструкция древней
доисторической хижины
основана на находках кругов
из камней и на изучении тех
строительных приемов, которые
и сейчас еще в ходу у групп
охотников-собирателей
в отдельных районах Африки.
Крышей мог служить сделанный
на скорую руку настил из веток
с листьями.
|
|

убитых хищниками, и куски лавы с близлежащих
гор, чтобы обработать их на месте. Около 1,7 миллио-
на лет назад они, кажется, пользовались также ору-
диями, которые приносили уже готовыми с гор, рас-
положенных на расстоянии 10 миль (16 км), что
может быть, означало даже и зачатки очень ранней
торговли.
![]()
![]()
![]()
![]() Тщательные исследования указывают на то, что
Тщательные исследования указывают на то, что
места большинства из примерно 20 очень ранних
стоянок в Олдувае были тщательно выбраны там,
где потоки свежей питьевой воды вливались в богатое
дичью солоноватое озеро. Возможно, наиболее за-
мечательным открытием явилось то, что эти древние
люди начали строить жилища. На одной стоянке
расположение костей и камней свидетельствует о
том, что с наветренной стороны люди были защи-
щены от ветра изгородью или щитом из кустарника.
На другой стоянке имеется круг из камней около
13 футов (4 м) диаметром, поддерживавших теперь
уже давно исчезнувшие ветки, из которых была со-
оружена хижина, подобная тем, которые и сейчас
еще строят кое-где в Африке. Это укрытие, возраст
которого примерно 1,8 миллиона лет, является пер-
вым известным нам человеческим сооружением в
мире.
Животный мир Олдувая.
Внизу и справа даны
изображения восьми животных,
нумерация которых
соответствует принятой в тексте
(относительные масштабы
изображений животных
не соблюдены).
1. Tilapia.
l
.Xenopus.
3. Chamaeleojacksonii.
4. Phoeniconaias.
5. Megantereon.
6.
Deinotherium.
l
.Sivatherium.
8. Pelorovis.
Тысячами расколотых костей животных усеяна
земля в местах ранних стоянок в Восточной Африке
где ранние люди жили и занимались разделкой туш
животных. Тщательное изучение стоянок Танзании
Кении и Эфиопии говорит о том, что 2— 1,5 миллиона
лет назад наряду с ранними людьми десятки крупных
и мелких животных обитали в саванне, в лесу и на
берегах озер. Наиболее детальные исследования
были проведены на местах бывших стоянок, распо
ложенных на берегах уменьшающегося в размерах
доисторического озера в том районе, который сейчас
прорезается Олдувайским ущельем. Расскажем
например, о некоторых из этих животных, как о сов
ременных, так и о вымерших (их сравнительные
размеры на рисунках не соблюдены).
1. Tilapia, цихлида, — пресноводная рыба, кото
рая может существовать и в солоноватой воде. Дли
на — до 12 дюймов (36 см).
2. Xenopus, шпорцевая лягушка, — водяная серая
лягушка с бледным брюшком. Длина — до б дюймов
(15 см).
3. Chamaeleo jacksoni — медлительная ящерица
которая ловит мух своим длинным клейким языком
у самцов — три рога на голове. Длина — до 12 дюй
мов (30 см). Среда обитания — кустарник и откры
тая лесистая местность.
4. Phoeniconaias, мелкий фламинго, — длинноно
гая болотная птица с кривым клювом. Длина 40 дюй
мов (1 м). Среда обитания — солоноватые озера
5. Megantereon, вероятно, был саблезубой кошкой
размером со льва, которая пронзала крупных тра
воядных животных своими кинжалоподобными
верхними клыками.
|
|

6. Deinotherium — «слон» с направленными вниз
бивнями в нижней челюсти, которые, возможно,
служили для выкапывания корней. Рост 13 футов
(4 м).
7. Slvatherium — жираф с короткой шеей и «олень-
ими рогами». Рост 7 футов (2,2 м).
8. ![]() Pelorovis — гигантский африканский буйвол с
Pelorovis — гигантский африканский буйвол с
размахом рогов 6 футов 7 дюймов (2 м). Он отно-
сился к семейству Bovidae (полорогие: крупный
рогатый скот, антилопы и т. д.); кости этих буйволов
составляют большинство из найденных в Олдувае.
|
|

![]() Глава
6 ЧЕЛОВЕК ПРЯМОХОДЯЩИЙ
Глава
6 ЧЕЛОВЕК ПРЯМОХОДЯЩИЙ
1,6 миллиона лет назад от
Homo habilis,
наиболее ве-
роятно, произошел более
крупный, обладающий боль-
шим мозгом
Homo erectus —
«человек прямоходящий».
Более высокоразвитый ин-
теллект и более совершен-
ная техника изготовления
орудий помогли этому охот-
нику раннего каменного
века колонизовать новые
места обитания
— заселить
небольшими группами Аф-
рику, Европу и Азию (глав-
ным образом Южную). Раз-
витие местных популяций,
по-видимому, проходило
различными путями. В Ев-
ропе
400 тысяч лет назад у от-
дельных особей появились
черты, встречающиеся у
ранних представителей
нашего вида
Homo sapiens.
200 тысяч лет назад
Homo
erectus,
вероятно, уже вы-
мер; возможно, он стал
жертвой конкуренции со
стороны своих собственных
потомков.
Два характерных для палеолита
топоровидных рубила (одно
из них — левое — обработано
более тщательно, другое —
менее тщательно); такие орудия
изготовляли человек
прямоходящий и ранний человек
разумный. Рубило, изображенное
на с. 119, обнаружено в Лондоне
около 1690 г. Это первое
изготовленное древним
человеком орудие, которое было
найдено археологами. Оба
орудия находятся в Британском
музее (Отделение
естествознания) в Лондоне.
![]()
Телосложение.
Человек прямоходящий показан
для сравнения рядом
с современным человеком.
![]() Рост 5—6 футов (1,5—1,8 м).
Рост 5—6 футов (1,5—1,8 м).
Масса 88—160 фунтов (40—
72,7 кг).
Homo erectus («человек прямоходящий») имел
более крупные мозг и тело, чем его вероятный предок
Homo habilis, и во многих отношениях уже был по-
хож на своего крепко сложенного непосредственного
потомка — современного человека. Его череп, хотя
и имел самые толстые стенки по сравнению с чере-
пами всех прочих представителей человеческой
трибы, сохранил архаические черты. Череп Homo
erectus был длинным и низко посаженным, с кости-
стой выпуклостью сзади, со скошенным лбом, тол-
стыми надглазничными валиками, более плоской,
чем у нас, лицевой частью, с крупными выдвинутыми
вперед челюстями, более массивными, чем у нас,
зубами (но все-таки немного меньшими, чем у Homo
habilis); подбородок отсутствовал. Сильные мышцы
сзади шеи были присоединены к заднему черепному
бугру и поддерживали голову с тяжелой лицевой
частью, не давая ей провисать вперед. Объем мозга
составлял в среднем 880—1100 CMJ
(мнения спе-
циалистов расходятся), что больше, чем у человека
умелого, хотя и меньше, чем у современного человека.
Некоторые взрослые особи, вероятно, достигали
роста б футов (1,8 м) и весили по крайней мере
столько же, сколько и мы.
Homo erectus жил от 1,6 миллиона до 200 тысяч
лет назад, а возможно, и в течение более длитель-
ного периода. Появившись впервые, вероятно, в Аф-
рике, отдельные группы распространились потом в
Европу, Восточную Азию (сюда относится пекин-
ский человек, или Sinanthropus) и Юго-Восточную
Азию (яванский человек, или Pithecanthropus).
Н По всей видимости, темпы эволюции отдельных
изолированных популяций были различны.
Передовая технология, включавшая применение
стандартного набора орудий труда, охота на круп-
ную дичь, пользование огнем и усовершенствование
методов строительства продвинули человека пря-
моходящего далеко вперед по сравнению с пред-
шествовавшими ему гоминидами, предоставив этому
виду возможность существовать в новых природных
и климатических условиях.
| А |
||||||
| £ |
I |
Ь\ |
V |
1 |
||
| 1 |
Б |
|||||
| № |
||||||
| 2млн. лет |
15 |
1 |
0,5 |
0 |
||
| Два возможных пути эволюции.
1 —
Homo habilis. предок. 3 — Homo sapiens. Б — Homo erectus как |
Сравнение черепов.
|
|
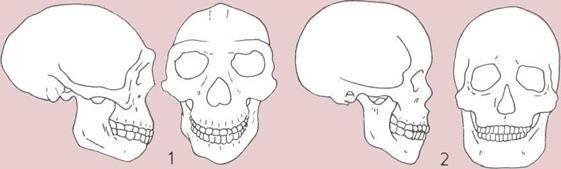
1. Череп Homo erectus:
с надглазничными валиками,
срезанным подбородком,
выступающими вперед
челюстями и с более мелкими
зубами, чем у Homo kahilis.
2. Череп Homo sapiens sapiens.
Головной мозг и м>||||||>|.
![]() На рисунках видно, что форма
На рисунках видно, что форма
черепа соотносится с размером
головного мозга и с величиной
мыши, уравновешивающих
голову и приводящих в движение
челюсти.
![]()
![]() А — Homo erectus (маленький
А — Homo erectus (маленький
мозг, крупные мышцы).
Б — Homo sapiens sapiens
(большой мозг, маленькие
мышцы).
Центры речи (внизу слева).
Над связанными между собой
центрами речи на поверхности
черепа образуются выпуклости,
заметные на ископаемых
костных остатках Homo (такие
же выпуклости, хотя и менее
ясно выраженные, встречаются
у человекообразные обезьян).
а — Поле Брока,
контролирующее речевую
деятельность.
б — Поле Вернике.
контролирующее понимание
речи.
| ГчГ-Чь |
va |
в |
|
| а |
6 |
||
Изменения климата.
На графике показаны колебания
температуры в июле на
территории Центральной Европы
за последние 1 200 000 лет.
(Согласно новейшим
исследованиям, чередование
пиков и понижений температуры
было более частым). Человек
прямоходящий существовал
в течение большей части этого
периода, за исключением
последних 200 000 лет.
Вероятный период существования человека пря-
моходящего примерно от 1,6 миллиона до 200 тысяч
лет назад совпадает с большей частью раннего и
среднего плейстоцена — геологической эпохи, про-
должавшейся примерно от 2 миллионов до 10 тысяч
лет назад. В течение этого ледникового века во
время периодов интенсивного похолодания, назы-
ваемых стадиями (фазами) оледенения, ледниковые
покровы и горные ледники расползались по значи-
тельной части территории севера Северной Америки
и Северо-Западной Евразии, отступая затем назад
во время перемежающихся с похолоданиями более
теплых промежутков, которые получили название
межледниковых стадий (фаз).
Во время оледенений даже не подвергавшиеся им
части Европы и Западной и Восточной Азии каж-
дый год были свободны от морозов лишь на протя-
жении едва ли одного месяца. Поэтому их ланд-
шафты превратились в тундру или леса с умеренно
холодной температурой; преобладали там такие де-
ревья, как ель и бук. Но условия похолодания бла-
гоприятствовали распространению крупных млеко-
питающих; например, в Китае это были гиены, ги-
гантские бобры, благородные олени и доисторические
виды носорогов и слонов.
В периоды оледенений в субтропики поступало
значительно больше дождевых осадков, чем они
получают сейчас, а тропики высыхали и их богатые
леса уменьшались в размерах, превращаясь в изо-
лированные островки.
Лишенные притока воды, которая была связана
в виде обширных ледниковых покровов, сокраща-
лись акватории океанов. Поверхность морей опуска-
лась по крайней мере на 328 футов (100 м) ниже их
современного уровня, обнажая сухопутные пере-
мычки, которые позволили людям заселить большие
острова Юго-Восточной Азии
В межледниковые периоды климат в некоторых
северных районах становился теплее, чем в настоя-
щее время. Теплолюбивые млекопитающие, такие
|
|
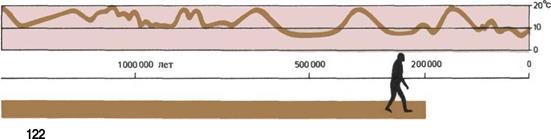
|
|
Мир в ледниковую эпоху.
как гиппопотамы и носороги Мерка, распростра-
нились на север вплоть до Южной Англии. В то же
время уровень моря поднялся на 180 футов (50 м)
выше современного, отделив от суши некоторые при-
брежные острова.
Любая популяция Homo erectus, подвергшаяся
изоляции в результате климатических изменений,
получала возможность эволюционировать слегка
отличным от других образом в зависимости от тех
условий, в которые она попадала.
Так выглядело Северное
полушарие во время оледенения.
а — Азия.
б — Европа.
в — Северная Америка.
1
— Территория, ставшая сушей
во время оледенения.
2 — Море.
3
— Территория, покрытая
ледниками.
4
— Летняя граница морского
льда.
|
|

Находки ископаемых остатков указывают на Аф-
рику как на континент, где смежные процессы, вли-
явшие друг на друга и связанные с усиленным ис-
пользованием рук, орудий и мезга, привели к увели-
чению размеров мозга у рода Homo, в результате чего
появился обладавший более крупным мозгом, более
развитым интеллектом и большей приспособляе-
мостью вид Homo erectus.
Первые известные нам ископаемые остатки челе-
века прямоходящего происходят из Восточной Аф-
рики и имеют возраст примерно 1,6 миллиона лет.
Один из скелетов представляет собой наиболее
хороню сохранившуюся из каких-либо гоминид,
дошедших до нас от периода, предшествовавшего
появлению преднамеренных захоронений, которые
начались около 70 тысяч лет назад.
Другие ископаемые — большей частью обломки
черепа или челюстей — говорят о том, что человек
прямостоящий в конечном счете распространился
из Восточной Африки в самые отдаленные уголки
континента. Но поскольку большинство ископаемых
находок состоят из отдельных мало связанных между
собой остатков и не дают нам ясного представления
о последовательно!! эволюции, а также потому, что
линия, отделяющая этот вид от нашего, выражена
нечетко, можно спорить о том, не относятся ли все
африканские и европейские остатки человека пря-
моходящего, которые моложе 400 тысяч лет, к виду
Homo sapiens.
Вот вкратце данные о некоторых наиболее значи-
тельных находках в Африке.
1. Тернифин; массивная нижняя челюсть с круп-
ными зубами, найденная рядом с двумя другими че-
люстями и фрагментам черепа. Возраст — возможно,
700 тысяч лет. Место — Тернифин, Алжир.
2. ![]()
![]()
![]() Кооби Фора; череп с тяжелыми надглазничными
Кооби Фора; череп с тяжелыми надглазничными
валиками; относится к числу наиболее полных и
самых ранних находок черепа Homo erectus. Воз
руст возможно, 1,6 милливна лет. Месте Кооби
Фора, к востоку от озера Туркана (Рудольф), Кения.
3. Сварткранс; фрагмент нижней челюсти с пятью
зубами, получивший сначала название "Telanfhro-
pus", а позже отнесенный к австралопитекам или
Homo habilis. Возраст возможно, 1 миллион лет.
Место — Сварткранс, Южная Африка.
вырасти до 6 футов (1,8 м).
что превышает рост большинства
современных .нолей. Скелет был
найден кенийским охотником
за ископаемыми остатками
Кимойя Кимеу в 1984 г. к западу
от озера Туркана (Рудольф).
Человек прямоходящий
в Африке.
Основные стоянки.
1. Тернифин.
2. Мелка Кунтуре.
3. Река Омо.
4. Нариокотоме.
5. Кооби Фора.
6. Чесованджа.
7. Олоргесайлие.
8. Олдувайское ущелье.
9. Сварткранс.
Три ископаемые находки
человека прямоходящего.
Нумерация соответствует
цифрам, которыми обозначены
в тексте находки остатков
Homo erectus в Африке.
1. Тернифин.
2. Кооби Фора.
3.
![]() Сварткранс.
Сварткранс.
|
|
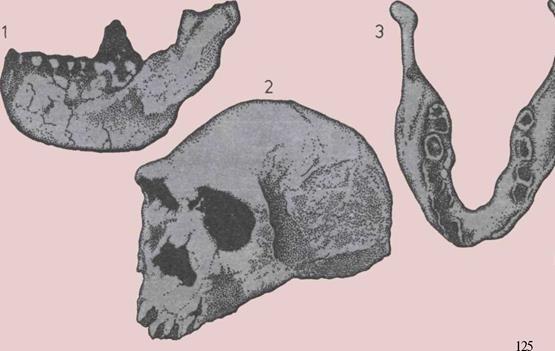
Человек прямоходящий
в Европе.
На карте отмечены некоторые
места находок костных остатков
или орудий, которые ученые
приписывают Homo ereclus.
Не исключено, однако, что все
они принадлежат архаичной
разновидности Homo sapiens.
1, 2. Амброна и Торральба.
3. Араго около Тотавеля.
4. Солейлак.
5. Терра Амата, Ницца.
6. Мауэр около Гейдельберга.
7. Бильцинглебен.
8. Пржезлетице.
9. Вертешсёллёш.
10.
![]()
![]() Петралона около
Петралона около
Салоники.
Древние орудия свидетельствуют о том, что Homo
erectus, возможно, появился в Европе около 1,5 мил-
лиона лет назад, но все найденные здесь костные
остатки, которые предположительно принадлежат
ему, как кажется, имеют возраст 500—200 тысяч лет
или датируются даже еще более поздним перио-
дом. Большинство из них представляют собой просто
фрагменты челюсти или черепа. Почти все они обла-
дают некоторыми особенностями, характерными для
Homo sapiens, что сбивает с толку исследователей.
Некоторые специалисты считают, что эти находки
принадлежат формам, являющимся переходными
между этими двумя видами. Возможно, их предки,
относящиеся к виду Homo erectus, пришли в Европу
в теплую фазу, когда ледниковые покровы отсту-
пили. Затем они эволюционировали в направлении
нашего вида, оказавшись в изоляции от других
человеческих популяций при похолодании в лед-
никовую эпоху.
Трудно решить вопрос о классификации этих ран-
них людей Европы. Некоторые ученые относят все
разновидности, о которых пойдет речь на следующих
двух страницах, к тем архаичным формам совре-
менного человека, которые будут описаны на с. 138—
139.
![]() 1. Гейдельбергский человек; массивная нижняя
1. Гейдельбергский человек; массивная нижняя
челюсть с зубами, без подбородка, соответствует
широкому выступающему вперед лицу. Возраст —
около 500 тысяч лет. Место — деревня Мауэр, близ
Гейдельберга, Германия.
2. Тотавельский череп; с большими надбровными
дугами, широким лицом и носовым отверстием, с
плоским лбом и длинной узкой мозговой коробкой.
Возраст — около 400 тысяч лет. Место — пещера
Араго около Тотавеля, Юго-Западная Франция.
3. Вертешселлешскии череп; фрагмент затылочной
кости — толстый, с костяным гребнем для прикреп-
ления шейных мышц. Объем мозга мог соответство-
вать нашему. Возраст — около 400 тысяч лет. Ме-
сто — село Вертешсёллёш, к западу от Будапешта,
Венгрия.
4. Петралонский череп; с широким основанием
и широкой лицевой частью, нависшими надбров-
ными дугами, скошенным лбом и угловатой заты-
лочной костью, но большого объема — около
1230 см3
. Возраст — около 300 тысяч лет. Место —
пещера Петралона около Салоники, Греция.
|
|
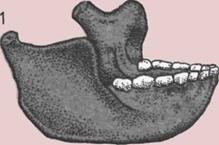
Четыре ископаемые находки.
Нумерация европейских
ископаемых костных остатков
соответствует принятой в тексте.
1.
Гейдельбергский человек
(нижняя челюсть из Мауэра).
2. Череп из Тотавеля.
3. Череп из села Вертешсёллёш.
4.
![]() Череп из пещеры Петралона.
Череп из пещеры Петралона.
|
|
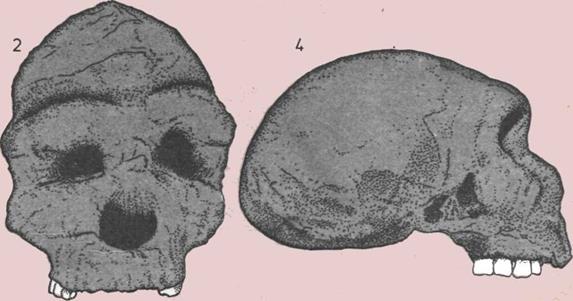
Большинство ископаемых остатков Homo erectus
происходит из Азии. Почти все они были найдены
на Яве или в Китае, а один череп, возможно принад-
лежащий человеку прямоходящему, был обнаружен
в Индии.
Наиболее ранние образцы «питекантропа» из так
называемых слоев Джетис на Центральной Яве
могут иметь возраст свыше 1,5 миллиона лет; в
яванских слоях Триниль найдены кости, возраст
которых, возможно, 700 тысяч лет. Ископаемый че-
ловек из Китая — синантроп (его называют также
пекинским человеком) — известен по остаткам более
40 особей, найденным около Пекина; все они исчезли
во время второй мировой войны, но слепки с них
сохранились. Мозг этой китайской разновидности
был крупнее, чем у более древних азиатских форм.
Синантроп существовал в условиях похолодания
около 360 тысяч лет назад.
![]() Все эти азиатские гоминиды жили недалеко от
Все эти азиатские гоминиды жили недалеко от
побережья Южно-Китайского моря, которое геоло-
ги сравнивают с гигантской ямой, то наполнявшейся
водой, то высыхавшей по мере таяния или наступле-
ния северных ледниковых покровов. Во время фаз
похолодания и понижения уровня воды Homo erectus,
вероятно, колонизировал опустившийся сейчас ниже
уровня моря Зондский шельф между Индонезией
и Китаем и мигрировал по образовавшейся суше
между двумя этими территориями.
Человек прямоходящий
в Азии.
На этой карте показаны места
некоторых наиболее важных
стоянок.
1. Нармада.
2. Юаньмоу.
3. Лук Йен.
4. Ланьтянь.
5. Юнцзи.
6. Наньчжао.
7. Пекин.
8. Хэцзян.
9. Саширан.
10. Пернинг, Моджокерто.
11.
![]() Трнниль.
Трнниль.
Вот примеры некоторых образцов из Азии.
1. «Питекантроп 4»; фрагмент крупного толсто-
стенного черепа и массивная верхняя челюсть с про-
межутком (диастемой) между клыком и резцом;
этому промежутку, вероятно, соответствовал боль-
шой клык на нижней челюсти. Возраст — около
1 миллиона лет. Место — Сангиран, остров Ява.
2. Череп из Ланьтяня; маленький (объем 780 см3
),
с толстыми стенками и с массивными, выгнутыми
дугой надглазничными валиками. У отдельно най-
денной в том же Ланьтяне нижней челюсти без
подбородочного выступа отсутствуют третьи корен-
ные зубы (эта врожденная особенность встреча-
ется у некоторых людей и сейчас). Возраст — около
600 тысяч лет. Место — округ Ланьтянь, провин-
ция Шэньси, Китай.
3. «Синантроп»; низкий широкий череп (объем
1075 см3
), относительно мелкие зубы без диастемы;
челюсть короче, чем у более древних азиатских
форм. Возраст — 360 тысяч лет. Место — селение
Чжоукоудянь близ Пекина, Китай.
Три ископаемые находки.
Нумерация рисунков
соответствует принятой в тексте.
1. «Питекантроп 4».
2. Череп из Ланьтяня.
3. «Синантроп».
|
|
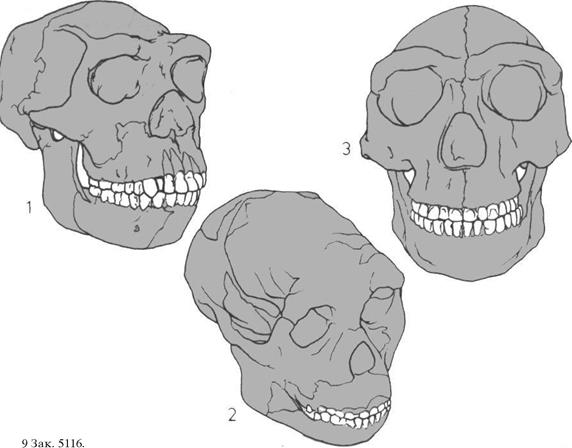
Около 1,6 миллиона лет назад в Восточной Африке
появился новый своеобразный тип каменных орудий.
Это так называемое ручное рубило состояло из куска
камня размером с кулак, которому была придана
форма, напоминающая ладонь или сплющенную
грушу; острые кромки камня были образованы пу-
тем скалывания отщепов с обеих сторон. Экспери-
менты показывают, что это орудие использовалось
главным образом при разделке туш, которые были
предварительно освежеваны при помощи острых
каменных отщепов, имевших форму топоровидных
лезвий (кливеров).
Самые первые ручные рубила появились примерно
в то же время, что и Homo erectus. Поскольку для
изготовления подобных однотипных орудий требо-
валась немалая сообразительность, можно заклю-
чить, что их вероятным изобретателем был именно
этот высокоразвитый гоминид.
К числу орудий древнего каменного века относятся
рубила, топоровидные кливеры, скрёбла и отщепы;
они называются ашельскими по находкам в Сент-
Ашёле на севере Франции; их возраст насчитывает
300 тысяч лет, Ашельская технология изготовления
орудий распространилась в Индии и Европе, где
она продолжала существовать примерно еще 100
тысяч лет назад, но, очевидно, так и не дошла до
Индонезии и Китая.
![]() Между тем культуры изготовления более грубых
Между тем культуры изготовления более грубых
рубящих орудий (чопперов), принадлежащие к тому
типу, который называется олдованским, распростра-
нились в Европе и Азии от Ближнего Востока до
Ашельские орудия.
Эти орудия из Анголы на
рисунках уменьшены примерно
в 2 раза.
1. Ручное рубило.
а — Тыльная сторона.
б — Режущий край.
в — Острие.
2. Кливер.
![]() г — Тыльная сторона.
г — Тыльная сторона.
д — Боковая сторона.
е — Режущий край.
|
|
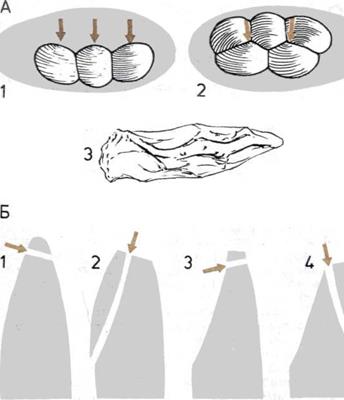
Явы, Филиппин и Чжоукоудяня в Северном Китае.
К местным вариантам относятся клэктонская куль-
тура из Клэктона-он-Си в Англии (где двухконус-
ные каменные ядрища, грубые рубила — чопперы,
толстые отщепы и зазубренные отщепы появились
еще до ашельской технологии) и тайякская культура
из Тайяка во французском департаменте Дордонь.
В одних местах ашельская технология и изготов-
ление чопперов из ядрищ существовали бок о бок,
а в других местах способы изготовления орудий, ве-
роятно, зависели от тех материалов, которые имелись
в наличии в том или ином месте, либо от того кон-
кретного вида работы, для которой они предназна-
чались.
К остаткам других орудий человека прямоходя-
щего относятся «наковальни» (рабочие плиты) и
ударники (отбойники). Некоторые из первых извест-
ных нам свёрл, лезвий и зубил и ранние образцы
костяных и деревянных орудий происходят из Амб-
роны и Торральбы в Испании; остатки деревянной
чаши найдены в Ницце во Франции.
Примитивная техника
обработки камня (А,
1—3).
1.
Ударами каменного молотка
по одной из сторон заготовки
получали ряд глубоких,
примыкающих друг к другу
сколов.
2.
Заготовку переворачивали
и наносили удары по
выступающим гребням, отчего
появлялся еще один ряд сколов.
3.
В итоге обработки
получалось ручное рубило
![]()
![]() с грубым волнистым режущим
с грубым волнистым режущим
краем, образованным
многочисленными наложенными
друг на друга сколами.
Усовершенствованная техника
обработки камня (Б,
1—4).
1.
Отколов верхнюю часть
заготовки, получали плоскую
ударную поверхность.
2.
Откалывали длинный тонкий
отщеп с одной из боковых сторон
заготовки.
3.
Следующим ударом готовили
новую верхнюю площадку.
4.
Ударом по этой площадке
откалывали длинный тонкий
отщеп с противоположной
боковой стороны заготовки.
В результате получался узкий
и более прямой режущий край,
нежели при примитивной технике
обработки.
Обработка камня с помощью
палки (В).
Рубила, изготовленные
с помощью каменных ударников
и техники, изображенной в серии
рисунков Б, подвергались
дальнейшей обработке ударами,
которые наносились упругой
костью, рогом или деревянной
палкой. С использованием
этой техники можно было,
не повредив заготовки, отделить
от ее поверхности мелкие отщепы
и придать изделию необходимую
форму.
а — Elephas antiquus, вымерший
слон; на юго-западе Европы
таких слонов убивали,
предварительно загнав их в
ловушку.
б — Sirnopithecus, вымерший
павиан; на него охотились
в Восточной Африке.
Остатки древнего пиршества.
На этом плане раскопок
в Амброне (Испания) мы видим:
а — Кости ископаемых слонов
и других животных.
б — Каменные орудия
и отброшенные за ненадобностью
отщепы.
![]() в — Обгоревшие поленья.
в — Обгоревшие поленья.
г — Камни, из которых,
возможно, был сложен очаг.
На ряде стоянок человека прямоходящего най-
дены серьезные доказательства того, что эти пред-
приимчивые гоминиды не просто собирали растения
и срезали мясо с туш убитых хищниками животных,
но и сами активно охотились на крупную дичь, объе-
диняясь для этого в группы с тем, чтобы спланиро-
вать и осуществить совместное преследование или
засаду. Находки, обнаруженные на трех континентах,
дают представление о приемах охоты и о животных,
которые были ее объектом. Все три следующих при-
мера, вероятно, датируются 400 тысяч лет до нашего
времени.
В Олоргесэйли (Кения) имеется одна стоянка с
остатками 50 обезьян — симопитеков. Древние люди,
вероятно, забили дубинками целое спящее стадо
этих крупных вымерших к настоящему времени ба-
буинов точно так же, как это еще делают и по сей
день некоторые танзанийские племена, охотясь на
современных потомков этих обезьян.
В Торральбе (северная часть Центральной Испа-
нии) охотники, видимо, использовали огонь, чтобы
загнать десятки бродячих слонов, диких быков, ло-
шадей, оленей и носорогов в естественную ловушку—
болотистый овраг в долине с крутыми склонами.
Здесь погибло по крайней мере 30 слонов, относя-
щихся к ныне вымершему виду с прямыми клыками;
эти слоны были более крупными, чем современный
|
|

африканский слон. Много диких зверей было, вероят-
но, забито в Торральбе и соседней Амброне.
Самые впечатляющие данные из Азии относятся
к пещере Чжоукоудянь близ Пекина. Здесь, судя по
пещерным отложениям, человек прямоходящий уби-
вал и поедал кабанов, бизонов, оленей, газелей,
лошадей и носорогов. Разбитые кости человеческих
конечностей и человеческие черепа с выломанным
основанием свидетельствуют о том, что эти охотники
были каннибалами, которые любили головной и
костный мозг особей, принадлежащих к их же соб-
ственному виду.
Не совсем ясным остается ответ на вопрос о том,
как древние люди убивали крупную дичь. Некото-
рые данные позволяют думать, что при этом они
пользовались деревянными копьями с каменными
наконечниками. Но какие бы способы ни применя-
лись, охота была связана с риском, и этим можно
объяснить то, что большинство черепов Homo erectus
имеют следы старых заживших повреждений.
Занятия охотников.
На рисунках изображены четыре
охотничьих орудия из Испании
(они пронумерованы) и
различные возможные виды их
применения (обозначены
буквами).
1. Деревянное копье.
а — Охота с копьями на крупных
млекопитающих.
2.
Каменное орудие с зубцами
по режущему краю (так
называемый «дентикюле»).
б — Заточка острия копья.
3.
Кливер из кварцита; его
длина 10 дюймов (25 см).
в — Разделка туши крупного
млекопитающего.
4.
Двустороннее скребло из
яшмы.
г — Удаление жира и мяса со
шкуры для ее очистки.
|
1 33 |
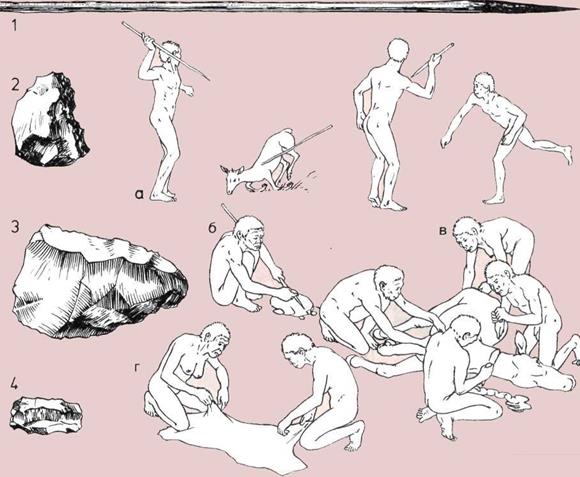
Распространение огня.
На карте показаны стоянки,
где более 100 тысяч лет назад
люди, очевидно, разводили
костры. Один из ученых
оспаривает доказательства
применения человеком огня
на стоянке 11. Стоянки 5, 6, 8
и 9, возможно, имеют возраст
1 миллион лет или древнее.
1. Торральба.
2. Эскале.
Терра Амата.
Вертешсёллёш.
Чесованджа.
![]()
![]() Каламбо Фоллз.
Каламбо Фоллз.
Скопления костей и каменных орудий находят на
тех местах, где семейные группы или более крупные
объединения Homo erectus разбивали свои стоянки.
Большинство из стоянок служило пристанищем
лишь в течение нескольких дней, пока люди плани-
ровали охоту, свежевали туши убитых животных и
делили между собой их мясо, собирали съедобные
растения, пили воду из соседнего родника, потока
или озера, обновляли запас своих орудий из камня,
дерева и кости, а также отдыхали и спали. В условиях
теплого климата тропиков было достаточно просто
найти для этого сухое место.
Но человек прямоходящий нередко сооружал и ук-
рытия, которые известны по находкам в прохладных
северных районах. В Торральбе и Амброне (Испа-
ния) обнаружены выложенные по кругу камни.
Подобные каменные круги остаются и сейчас на ме-
стах установки эскимосских жилищ — сшитых из
шкур шатров, средняя часть которых поддерживает-
ся центральным шестом, а края придавливаются
к земле тяжелыми камнями. На стоянке Терра Ама-
та близ французского города Ниццы на площади
90 кв. миль (235 км2
) сохранились старые жилые
площадки, на месте которых, возможно, были соору-
жены овальные хижины из переплетенных веток,
закрепленных камнями. Внутри хижин горели кост-
ры в очагах, защищенных от ветра каменными ук-
рытиями. Самые крупные хижины этой сезонной сто-
янки на морском берегу могли вмещать до 20 чело-
век.
![]() Близ Тотавеля во Французских Пиренеях и Чжо-
Близ Тотавеля во Французских Пиренеях и Чжо-
укоудяне около Пекина охотники жили в пещерах.
По-видимому, в пещеру Араго (Франция) они при-
ходили в те или иные сезоны вслед за мигрирующей
дичью. Но обнаруженный в Чжоукоудяне слой
пепла толщиной 19,7 футов (6 м), может быть,
является следствием длительного пребывания людей.
|
|
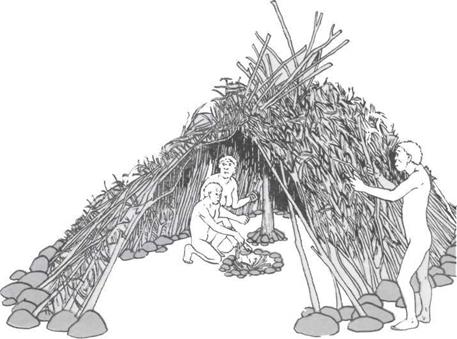
| Огонь был, по-видимому, знаком людям еще до В последний ледниковый период сооружение жи- Жилише на Ривьере. В таких овальных шалашах, |
![]()
![]()
![]() 300 тысяч лет назад неко-
300 тысяч лет назад неко-
торые архаичные формы ви-
да
Homo sapiens
начали при-
обретать те основные призна-
ки, которые характерны для
неандертальского человека.
Лучше всего он известен нам
по костным остаткам и оруди-
ям, возраст которых колеб-
лется от
70 до
40 тысяч лет.
Этот преимущественно евро-
пейский подвид явился охот-
ником-собирателем, который
принес с собой много ново-
введений. Он жил в среднюю
пору древнего каменного ве-
ка и был способен переносить
довольно сильные похолода-
ния климата. Погребения
умерших свидетельствуют,
что у древнейших извест-
ных нам неандертальцев
проявлялись некоторые чув-
ства, характерные для чело-
века. И все же неандерталь-
цы были, вероятно, лишь бо-
ковой ветвью того эволюци-
онного древа, от которого
произошел человек вполне
современного типа. По всей
видимости, эта ветвь вымерла
30 тысяч лет назад.
![]()
300 тысяч лет назад Homo erectus уже начал пре-
вращаться в человека вполне современного типа,
который изображен на с. 160. Но темпы эволюции
были разными в различных местах, что зависело
от таких факторов, как мутации, все более широкое
использование орудий для тяжелых работ, выпол-
нявшихся раньше при помощи зубов или мышц,
миграция и изоляция некоторых популяций. Даже
эволюция различных частей человеческого тела про-
ходила неравномерно, что приводило, например, к
появлению черепов, сочетавших в себе характерные
современные и архаичные черты. Несмотря на раз-
личия, палеоантропологи относят большинство че-
ловеческих остатков позднего ледникового периода
к одному виду, считая их архаичными формами Homo
sapiens. Сюда причисляют также ответвившийся
специфический подвид — неандертальцев, о которых
пойдет речь на с. 142-143. Здесь мы даем примеры
архаичных форм Homo sapiens, включая некоторых
так называемых «протонеандертальцев». (Не исклю-
чено, что к виду Homo sapiens относится также рас-
смотренный выше европейский Homo erectus).
1. Человек из Сванскомба; фрагменты женского
черепа с толстыми стенками, округлым затылком и
большим объемом мозга (1300 см ). Возраст — воз-
можно, 250 тысяч лет. Место — Сванскомб около
Лондона, Англия.
![]()
|
2. Человек из Штейнгейма; низкий череп с над-
глазничными валиками и объемом мозга 1100 см3
,
но с округлым затылком, относительно маленькой
выпрямленной лицевой частью и мелкими зубами.
Возраст — более 300 тысяч лет. Место — Штейн-
гейм около Штутгарта, Германия.
3. Человек из Брокен-Хилла («родезийский чело-
век» или «череп из Кабве»); череп с покатым лбом,
мощными надглазничными валиками и угловатым
затылком, но с крутыми, «с3
овременного типа», ску-
лами; объем мозга 1300 см3
. Возраст — возможно,
200 тысяч лет. Место — Кабве, Замбия.
4. Череп из бухты Салданья; похож на череп
человека из Брокен-Хилла. Возраст — по крайней
мере 200 тысяч лет. Место — Хоупфилд, бухта Сал-
данья, Южная Африка.
5. Человек с реки Соло; черепная коробка с тол-
стыми стенками, покатым лбом, сплющенной лоб-
ной костью и развитым в задней части гребнем;
объем мозговой полости 1035—1255 см3
; очень
похож на череп более раннего пекинского синантро-
па. Возраст — возможно, более 100 тысяч лет. Мес-
то — Нгандонг, река Соло, остров Ява.
6. Человек из Чжуцияо; один из группы массив-
ных черепов, найденных в Китае; похож на череп
Homo erectus, но округлен сзади. Возраст всей груп-
пы черепов 250—100 тысяч лет. Место — Датун, про-
винция Шаньси, Китай.
Шесть черепов древних
Homo
sapiens.
Нумерация соответствует
принятой в тексте.
1.
Человек из Сванскомба
(вид сзади).
2. Человек из Штейнгейма.
3.
Человек из
Брокен-Хилла.
4. Череп из бухты Салданья.
5. Человек с реки Соло.
6.
Человек из Чжуцзияо
(затылочная кость из задней
части черепа).
|
|
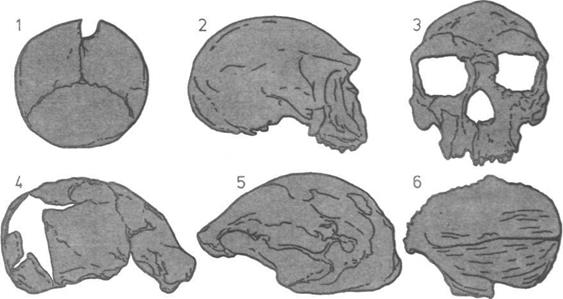
От 200 до 40 тысяч лет назад климат северных
регионов определялся волнами похолоданий, кото-
рые чередовались с более теплыми промежутками.
Именно к этому периоду времени относятся неан-
дертальцы и их непосредственные предшественники.
По традиции геологи делят эту часть плейстоцено-
вой эпохи на две стадии оледенения (в Европе их
часто называют рисс и вюрм), разделенных межлед-
никовьем, хотя в результате исследований установ-
лено большее число ледниковых и межледниковых
периодов. Связанные с ними изменения климата
влияли на уровень моря.
![]()
![]() Во время наиболее сильного похолодания в Север-
Во время наиболее сильного похолодания в Север-
ной Америке мамонты ушли на юг, а в Европе неко-
торые их виды оказались в ловушке между надви-
гающимися с севера ледниковыми покровами и аль-
пийскими ледниками. В Западной и Центральной
Европе леса полосы умеренного климата уступили
место степям или тундре. Здесь могли выжить толь-
ко приспособившиеся к холоду животные, к которым
относились такие новые виды, как покрытый шерстью
мамонт, шерстистый носорог и мускусный бык (ов-
цебык). Когда ледниковые покровы сокращались,
все эти виды мигрировали на север по мере отступ-
ления степей и тундры. С юга на север продвига-
лись и леса, в которых обитали гиппопотамы, слоны
с прямыми бивнями, гигантские лоси, львы и леопар-
ды. Когда похолодание возвращалось снова, мигра-
ция проходила в обратном направлении.
Европа ледникового периода.
На этой карте показаны суровые
климатические условия,
в которых жили европейские
неандертальцы во время
вюрмской стадии оледенения.
На карте видны древняя
и современная береговые линии.
1
— Ледяные покровы и ледники
во время максимального
оледенения.
2
— Вечная мерзлота (участки
земли с постоянно замерзшей
подпочвой).
3
![]() — Древняя береговая линия.
— Древняя береговая линия.
Здесь мы расскажем о четырех приспособившихся
к холоду млекопитающих.
1. Mammuthus primigenius, волосатый мамонт с
длинной черной шерстью, густым подшерстком, ма-
ленькими ушами и огромными загнутыми вверх бив-
нями. Рост 9 футов 6 дюймов (2,9 м). Время — сред-
ний и поздний плейстоцен. Место — Евразия и Се-
верная Африка.
2. Coelodonta antiquitatis, шерстистый носорог;
имел косматую шерсть и два рога, передний из ко-
торых был более длинным. Рост 6 футов б дюймов
(2 м). Время — средний и поздний плейстоцен. Ме-
сто — Евразия и Северная Африка.
3. Ursus spelaeus, пещерный медведь; имел боль-
шую голову с мощными челюстями, был всеяден.
Существовали как крупные, так и карликовые раз-
новидности. Длина от кончика носа до хвоста дости-
гала 9 футов (2,7 м). Время — середина позднего
плейстоцена. Место — Евразия.
4. Ovibos moschatus, мускусный бык (овцебык);
у него большая голова с рогами и массивное туло-
вище, покрытое подшерстком и длинными темными
волосами, свисавшими почти до самой земли. Рост —
до 5 футов (1,5 м). Время — от среднего плейсто-
цена до наших дней. Место — Северная Евразия и
Северная Америка.
Четыре млекопитающих
ледникового периода.
Нумерация рисунков
соответствует принятой в тексте.
1. Mammuthusprimigenius.
2. Coelodonta antiquitatis.
3. Ursus spelaeus.
4. Ovibos moschatus.
|
|

Homo sapiens neanderthalensis получил свое имя
по ископаемым остаткам, найденным в долине Неан-
дерталь близ Дюссельдорфа, Германия. Так называ-
емый классический неандерталец из Европы имел
крупную удлиненную голову; мозг его был больше на-
шего, а стенки черепа толще, чем у нас, но тоньше,
чем у Homo erectus. На человека прямоходящего
неандерталец был еще несколько похож своими мощ-
ными надглазничными валиками и покатым лбом.
У неандертальца был четко выраженный, похожий на
шишку затылочный бугор с большим основанием, к
которому прикреплялись шейные мышцы. Широкая
лицевая часть сильно выдвинута вперед и скошена
назад по бокам, что придавало скуловым костям «об-
текаемую» форму. Большой нос мог быть плоским
или выступающим. Резцы мошной нижней челюсти,
которая не имела подбородочного выступа, были
крупнее, чем у нас, а коренные зубы имели, как пра-
вило, крупную полость пульпы.
Классические неандертальцы были невысоки, чрез-
вычайно мускулисты и коренасты, с большими су-
ставами ног и рук. Пропорциями тела они похожи
на эскимосов, чье плотное сложение помогает им
сохранять тепло в условиях холодного климата. Но,
как будет видно из последующего изложения, отдель-
ные особи и популяции имели свои особенности.
Неандертальский подвид произошел от архаич-
ной формы Homo sapiens, возможно, еще 200 тысяч
лет назад. Физические данные и передовые техниче-
ские приемы неандертальцев сделали некоторых из
этих людей среднего палеолита, возможно, первыми
гоминидами, способными выносить тяготы зимы в ус-
ловиях холодного климата. Кроме того, ритуальное
поведение неандертальцев свидетельствует о дости-
жении ими более высокого уровня развития чело-
веческих эмоций и самосознания.
![]()
![]()
![]() Несмотря на все это, около 30 тысяч лет назад
Несмотря на все это, около 30 тысяч лет назад
эта группа, очевидно, вымерла. Некоторые ученые
| 1 шштш |
|
| а |
^6 |
| 2 мм |
..... |
| а |
|
| 3 а |
I: |
Сравнение черепов.
А — На этих двух изображениях
черепа неандертальца (сбоку
и спереди) заметны следующие
особенности.
а — Большой объем черепа.
б — Шишковидный затылочный
бугор.
в — Длинная низкая форма
черепа.
г — Покатый лоб.
д — Надглазничные валики.
е — Обтекаемые скулы.
ж — Широкое продолговатое
носовое отверстие.
з — Выступающая вперед
средняя часть лица.
и — Крупные (но более мелкие,
чем у Homo erectus) зубы.
к — Массивная нижняя челюсть,
обычно без подбородочного
выступа.
![]()
![]()
![]() Б — Для сравнения показан
Б — Для сравнения показан
череп современного человека
(вид сбоку и спереди).
| \ |
\ |
\ |
||
| а |
6 |
|||
| \ |
\ |
\ |
||
Сравнение бедренных костей.
1.
Бедренная кость
неандертальца — толстая,
массивная, с заметным изгибом.
2.
Бедренная кость современного
человека; она тоньше, слабее
и прямее, чем у неандертальца.
![]()
![]() Объем мозга (слева).
Объем мозга (слева).
а — Неандерталец, 1500 см3
.
б — Современный человек,
1400 см3
.
Места обитания неандертальцев.
Показаны стоянки европейских
неандертальцев
и «прото неандертальцев»;
район, выделенный
прямоугольником, приведен
в увеличенном масштабе.
1. Гибралтар.
2. Понтневидд.
3. Спи-сюр-ль'Орно.
4. Долина реки Неандер.
5. Саккопасторе.
6. Монте-Чирчео.
7. Крапина.
8. Кулна.
9. Субалюк.
10. Староселье.
11. Киик-Коба.
12. Джрухула.
13. Уртиага.
14. Мас д'Азиль.
15. Баньолас.
16. Ортю.
17. Ла-Шапель-о-Сен.
18. Ле-Мустье.
19. Ла-Ферраси.
20. Ла-Кина.
21. Ла-Шэз.
22. Фонтешевад.
23. Сен-Сезэр.
К восьмидесятым годам XX века ученые нашли
в Европе остатки примерно 200 неандертальцев или
«протонеандертальцев»; находки были сделаны боль-
шей частью в пещерах. Сюда относятся самые древ-
ние крупные остатки ископаемых людей из найден-
ных на каком бы то ни было континенте. По некото-
рым из них видно, что даже особи, проживавшие
совместно в одной группе, значительно отличались
друг от друга по строению черепа и челюстей.
Более половины всех найденных особей происхо-
дят из Франции — это фрагменты остатков 116 че-
ловек примерно с трех десятков стоянок. На две сто-
янки — Ортю и Ла-Кина — приходится большин-
ство остатков, найденных на территории Франции.
Черты, характерные для неандертальцев, присущи
также примерно двум десяткам особей со стоянки
Крапина (Югославия), одиннадцати из Италии, де-
сяти из Бельгии, восьми из Германии и еще несколь-
ким из других мест, включая Великобританию, Ис-
панию, Гибралтар, Чехословакию и Крым.
Возраст находок колеблется, возможно, от 250 до
30 тысяч лет, но наиболее «зрелые» неандертальцы
относятся к первой половине последнего (вюрмского)
оледенения: 70—30 тысяч лет назад. Среди наших
|
|
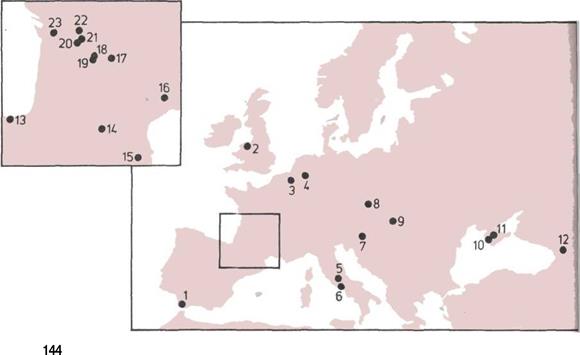
примеров представлены находки, относящиеся к раз-
личным периодам и регионам.
1. Череп из Эрингсдорфа. К этим остаткам отно-
сятся череп неандертальского типа, но с высоким
лбом и челюсть без подбородочного выступа, но с
маленькими зубами. Возраст — возможно, 200 тысяч
лет. Место — Эрингсдорф, Германия.
2. Понтневидд Зуб и фрагменты челюсти, най-
денные в 1980-х годах, имеют особенности, прису-
щие неандертальцам. Возраст — около 250 тысяч лет.
Место — пещера Понтневидд близ Рила, Северный
Уэльс.
3. Фонтешевад. На этой стоянке найдены фраг-
менты черепов с надглазничными валиками и без
них. Возраст — возможно, 150 тысяч лет. Место —
Фонтешевад, Западная Франция.
4. Ла-Шапель-о-Сен. Эта стоянка известна ске-
летом классического неандертальца — старика,
больного артритом. Возраст — возможно, 50 тысяч
лет. Место — Ла-Шапель-о-Сен, южная часть Цент-
ральной Франции.
5. Гибралтар. Здесь нашли череп фактически еще
раньше, чем в долине Неандерталь. Возраст 50 ты-
сяч лет.
6. Чирчео. Здесь найден искалеченный череп чело-
века. Возраст — около 45 тысяч лет. Место — Монте-
Чирчео около Рима, Италия.
7. Неандер. Первый из описанных в литературе
скелетов неандертальцев был найден в пещере на
берегу этой реки. Возраст — возможно, 50 тысяч
лет. Место — пещера Фельдхофер, Неандерталь
(«долина Неандер»), Дюссельдорф, Германия.
8. ![]()
![]() Крапина. Найдены широкие укороченные чере-
Крапина. Найдены широкие укороченные чере-
па с мощными надглазничными валиками; эти люди,
возможно, были жертвами пиршества каннибалов.
Возраст — около 100 тысяч лет. Место — Крапина,
Северная Югославия.
|
|
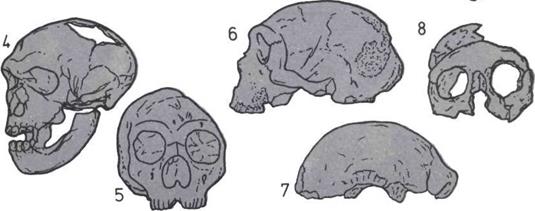
Череп неандертальца.
Бочкообразный череп
неандертальца (вид сзади)
со стоянки Шанидар-1; наиболее
широкая часть черепа
располагается выше, чем у
черепов австралопитека и Homo
erectus. Судя по отверстиям
в черепе, этот человек погиб
во время обвала.
Места обитания неандертальцев.
Нумерация соответствует
принятой в тексте и на рисунках
(с. 147).
1. Джебель-Ирхуд.
2. Табун.
3. Схул.
4. Амуд.
5. Кафзех.
6. Шанидар.
7.
![]()
![]() Тешик-Таш.
Тешик-Таш.
Неандертальцы за пределами
Европы
Неандертальцы обитали также в Юго-Западно
Азии и, возможно, в Африке, но у некоторых из ни
отсутствовали те грубоватые черты, которые был
характерны для классической европейской формы
Вероятно, у европейских неандертальцев такие чер
ты возникли в результате приспособления к сильным
холодам ледникового периода. Некоторые из азиат
ских и африканских неандертальцев имели боле
прямые и тонкие конечности, менее мощные над
глазничные валики и укороченные, менее массив
ные черепа. Наряду с надглазничными валикам!
и выдвинутым вперед лицевым отделом некоторые
черепа имели высокий лоб и высокую округлую че
репную коробку. За пределами Европы и Юго-За
падной Азии не было найдено типично неандерталь
ских остатков. Примерно 40 тысяч лет назад послед
ние неандертальцы Юго-Западной Азии, очевидно
существовали одновременно с людьми, имевшими
вполне современный облик. Некоторые из черепов
описанных ниже, тоже уже можно классифицировать
как почти современные.
1. Джебель-Ирхуд; удлиненный и невысокий че
реп с крупными надглазничными валиками, но с ли
цевым отделом современного типа и с небольшим
затылочным гребнем. Возраст — около 7 тысяч лет
Место — Джебель-Ирхуд, Марокко.
2. Табун; череп невысокий, с покатым лбом, над
глазничными валиками и утолщенными резцами
тем не менее лицевая часть и затылок имеют совре
менный вид. На нижней челюсти есть подбородочный
выступ. Изогнутые кости конечностей напоминают
кости европейских неандертальцев. Возраст 50 тысяч
лет. Место — пещера Табун, гора Кармель, Израиль
3. Схул-5; сочетаются большой объем мозговой по-
лости, надглазничные валики, довольно высокий
|
|
лоб. а также лицевой отдел и затылок современного
типа. Возраст 40 тысяч лет. Место — каменное
укрытие Схул, захоронение на горе Кармель, Из-
раиль.
4. Амуд-1; черен неандертальского типа объемом
1740 см (относится к числу крупнейших из ныне
известных) и удлиненные кости конечностей. Воз-
раст — возможно, 45 тысяч лет. Место — пещера
Амуд около Тивериадского озера, Израиль.
5. Кафзех; череп схож с предыдущим. Возраст -
возможно, 45 тысяч лет. Место — пещера Кафзех,
Израиль.
6. Шанидар; классический неандерталец с круп-
ным мозгом, но надглазничные валики не соеди-
нены, как у европейских неандертальцев. Возраст —
возможно, 70—45 тысяч лет. Место — пещера Ша-
нидар, Северный Ирак.
7. ![]() Тешик-Таш; череп мальчика с неразвитыми над-
Тешик-Таш; череп мальчика с неразвитыми над-
глазничными валиками и с другими классическими
признаками; лицевой отдел черепа и конечности со-
временного типа. Возраст — возможно, 45 тысяч лет.
Место пещера Тешик-Таш, Узбекистан, СССР.
|
|
|
|

![]()
![]()
10 * 147
Успехи в технике обработки
камня.
![]() А — Чтобы изготовить это
А — Чтобы изготовить это
ашельское топоровидное
рубило, прямоходящий человек
нанес по нему 65 ударов.
Б — Неандертальцу
понадобилось совершить
111 ударов для изготовления
этого ножевндного
остроконечника, характерного
для мустьерской культуры.
Увеличение числа ударов
и использование заранее
подготовленных ядрищ
позволяло экономить материал
и изготовлять большее число
различных тщательно
отделанных специализированных
орудий.
1. Остроконечник; отщеп, отделанный в виде длин-
ного острого треугольного наконечника, который,
возможно, привязывали к деревянному древку или
вставляли в расщеп, изготовляя таким образом стре-
лу или копье.
2. Скребло; выпуклое скребло с утолщенной ра-
бочей кромкой, применявшееся, возможно, для выде-
лывания шкур.
3. Нож; длинный отщеп с острым лезвием и тупой
(оббитой) тыльной частью, на которую можно было
нажимать рукой; применялся для свежевания туш,
разделки мяса, обработки дерева.
4. Зазубренная пилка («дантикюле»); отщеп с от-
деланным пилообразным режущим краем, пригод-
ным для обработки дерева.
5. ![]() Орудие с выемкой; отщеп с выемкой, пригод-
Орудие с выемкой; отщеп с выемкой, пригод-
ный для шлифования палок, которые, возможно,
использовались как копья.
Длина получаемого режущего
края.
А — Длина режущего края,
которую получал на 1 фунт камня
человек прямоходящий.
Б — Длина режущего края,
которую получал на 1 фунт
камня неандерталец.
|
|
Смертоносная ловушка.
![]() При попытке схватить мясную
При попытке схватить мясную
приманку эта лисица должна
сместить тяжелый камень,
который упадет и задавит ее.
Такие ловушки, еще и сейчас
применяемые эскимосами,
вошожно, помогали
неандертальцам обеспечивать
себя лисьими шкурами для
изготовления одежды.
Неандертальцы были, вероятно, самыми умелыми
охотниками своего времени. Иначе они не смогли бы
выжить в условиях сурового холодного климата
ледникового периода. Летом можно было питаться
корнями и ягодами. Но зимой группы неандерталь-
цев могли рассчитывать только на добычу мяса во
время совместной охоты на млекопитающих, обита-
ющих в тундре и лесах холодного пояса.
По костям, найденным в пещерах и на открытых
стоянках, можно сказать, что основную добычу евро-
пейских неандертальцев составляли такие крупные
животные, как бизоны, пещерные медведи, лошади,
северные олени, дикие быки, волосатые мамонты и
шерстистые носороги. Так, одни охотники специали-
зировались по бизонам (стоянка Ильская на Се-
верном Кавказе), а другие по северным оленям
(Зальцгиттер-Лебенштадт, Германия). Мелкая
добыча включала лис, зайцев, птиц и рыб. Только на
одной венгерской стоянке было найдено свыше 50
тысяч костей 45 видов крупных и мелких животных.
(Однако кажется, что многие кости медведей и
мамонтов на некоторых стоянках были взяты от туш
животных, погибших естественной смертью).
Без сомнения, у некоторых животных ценилось
не только мясо, но и шкуры, кости и сухожилия, кото-
рые использовались для изготовления одежды, жи-
лищ и ловушек.
![]() У нас мало данных о том, как охотились эти люди.
У нас мало данных о том, как охотились эти люди.
Возможно, они метали копья или кидали связанные
шары (бола), сбрасывали валуны с утесов или ста-
вили силки и рыли ловчие ямы. Добычей охотников
становились больные, старые, совсем молодые или
ослабленные животные и даже пребывающие в зим-
ней спячке медведи. С помощью огня люди могли
загонять целые стада испуганных лошадей на утесы
или в каньоны, заканчивающиеся тупиками, и устра-
ивать там массовую бойню.
Предполагается, что для таких охотничьих хитро-
стей надо хорошо знать привычки животных: как
они едят, где пьют, в каких местах кочуют. Необ-
ходимо было также тщательно планировать охоту
и обеспечивать взаимодействие ее участников.
![]()
![]() В Юго-Восточной Азии, вне зон периодических
В Юго-Восточной Азии, вне зон периодических
похолоданий, группы мужчин, без сомнения, про-
ходили большие расстояния, преследуя и убивая
крупных животных, таких как дикие быки, овцы,
козлы. Тем временем женщины и дети прочесывали
местность вокруг стоянки в поисках грызунов, пре-
смыкающихся, насекомых, ягод, смолы, меда, корней
и клубней.
Процентный состав добычи
охотников.
Эти данные основаны на
подсчете костей животных,
убитых 55 тысяч лет назад
и найденных на летней
лагерной стоянке
Зальцгиттер-Лебенштадт в
Германии.
а— Северный олень, 72%.
б — Волосатый мамонт, 14%.
в — Бизон, 5,4%.
г — Лошадь, 4,6%.
д — Шерстистый носорог, 2%.
е — Другие животные, 2%.
Промысловые животные.
Все эти 15 видов животных из
различных сред обитания (одни
из них были приспособлены
к холоду, другие —
теплолюбивые), вероятно,
являлись добычей охотников-
неандертальцев.
1. Дикий бык.
2. Пещерный медведь.
3. Бурый медведь.
4. Горный альпийский козел.
5. Шерстистый носорог.
6. Северный олень.
7. Волосатый мамонт.
8. Бизон.
9. Лошадь.
10. Окунь.
11. Шука.
12. Песец.
13. Полярный заяц.
14. Черепаха.
15. Аист.
Шалаш в пещере?
Судя но результатам раскопок
пещеры Ле-Лазаре около Ниццы
во Франции, можно
предположить, что 150 тысяч лет
назад древние Homo sapiens
для защиты от холода
соорудили внутри пещеры
жилище из шкур, разделенное
на две част».
![]() А — Заднее отделение.
А — Заднее отделение.
Б — Переднее отделение.
а — Стена каменного укрытия,
которая, возможно, была
укреплена кольями.
б — Места, где, вероятно,
располагались выходы.
в — Предполагаемая
перегородка.
г — Мусор, возможно
включающий остатки подстилок.
д — Очаги.
Европейские неандертальцы смогли пережить су-
ровые зимы ледникового периода только благодаря
тому, что создавали для себя теплый микроклимат
с помощью одежды и обогреваемых жилищ.
В некоторых местах естественными укрытиями
от холода являлись пещеры. Группы людей числен-
ностью до 40 человек жили во многих десятках из-
вестняковых пещер в Дордони на Юго-Заиаде Фран-
ции; по крайней мере два десятка пещер известно
на территории Европейской России. Ямка от столба
у входа в пещеру Комб-Греналь во Франции говорит
о том, что здесь имелся занавес из шкур, который
защищал от ветра, снега и дождя. Внутри пещеры
Куэва Морин в Северной Испании сохранилась даже
каменная стенка.
Там, где не было пещер, группы охотников стро-
или укрытия в открытых местах. Среди этих укры-
тий от стихий наиболее внушительное впечатление
производят большие шалаши или хижины, построен-
ные в долинах украинских рек, где было очень много
крупной дичи. Здесь люди возводили из веток соору-
жения длиной до 30 футов (9 м), шириной до 23 фу-
тов (7 м) и высотой до 10 футов (3 м). Очевидно,
эти каркасы покрывали шкурами, которые закреп-
ляли тяжелыми костями мамонта (они были най-
дены вокруг хижин).
Следы старых очагов свидетельствуют о том, что
неандертальцы обогревали свои шалашы и пещеры,
сжигая дрова или кости. Они уже знали, как разво-
дить огонь, высекая искры из кусков колчедана и
применяя в качестве трута сухие грибы-трутовики.
![]()
![]()
![]() Об одежде можно судить главным образом по кос-
Об одежде можно судить главным образом по кос-
венным данным. Люди, очевидно, пользовались
каменными ножами для раскройки меховых шкур,
прокалывали дырочки в шкурах каменным или ко-
стяным шилом, а затем сшивали эти шкуры сухо-
жилиями. Снимая шкуры с лис, зайцев, волков, они,
очевидно, иногда сохраняли лапки для связывания
этих шкур. В результате, вероятно, получались гру-
бые штаны, рубашки, плащи, колпаки и обмотки
для ног.
Изготовление одежды.
1.
Очистка шкуры — удаление
с нее мяса и жира.
2.
![]() Скрепление шкур кожаным
Скрепление шкур кожаным
шнурком, который продевали
через отверстия, проколотые
шилом.
Погребения, ритуалы, искусство
Захоронения, ритуалы и зачатки искусства гово-
рят о том, что неандертальцы в большей степени
обладали самосознанием, общественными интереса-
ми и в целом были более способны к абстрактному
мышлению, чем их предок Homo erectus.
Неандертальцы явились первыми людьми, которые
систематически хоронили своих мертвых. К началу
второй половины XX века ученые раскопали 68 захо-
ронений, главным образом в Европе, и почти все в
пещерах, где было найдено более 150 тел.
Имеются несомненные доказательства того, что
погребение умерших было обрядом. Скелеты лежали
в ямах, вырытых в полу пещер. Многие были уло-
жены в позу спящего человека и снабжены пред-
метами, которые, как считалось, должны были со-
путствовать умершему в его могиле: от каменных
орудий и кусков жареного мяса до каменных поду-
шек, подстилки из лесного хвоща и разбросанных
вокруг поздних весенних цветов (о чем можно су-
дить по оставшейся от них пыльце).
![]()
![]()
![]() Все это говорит о том, что неандертальцы прида-
Все это говорит о том, что неандертальцы прида-
вали значение жизни и смерти отдельного человека
Погребения в Шанвдаре
(вверху).
На атой блок диаграмме
нанесены неандертальские
погребения (некоторые из них
были завалены камнями)
в большей пещере Шанидар
на севере Ирака. Глубина
раскопок 45 футов (13,7 м).
А — Поздний палеолит.
28,7—35 тысяч лет назад.
Б — Мустьерекая культура.
35—100 тысяч лет назад.
1. Старик-калека.
2—4. Жертвы обвала.
5. Мужчина, погребенный
вместе с цветами.
6—8. Одно мужское и два
женских погребения.
9. Ребенок.
Погребение неандертальца
(справа).
На этой сложной реконструкции
можно видеть:
а — Тело умершего в позе
спящего.
![]() б — Тело ориентировано
б — Тело ориентировано
в направлении восток запад.
в — Голова повернута в южном
направлении.
г — Каменная подушка.
д — Сожженные кости.
е — Орудия из камня.
ж — Подстилка из лесного
хвоща.
з — Цветы.
Пиршество каннибалов?
![]() У этого человеческого черепа
У этого человеческого черепа
расширено затылочное отверстие
и выломано основание. Черед
образовавшуюся большую
дыру кто-то, вероятно, выскреб
и съел головной мозг, а затем
поместил череп внутрь круга
из камней, которые найдены
в пещере Гуаттари на стоянке
Чирчео в Италии.
и, возможно, задумывались о загробном существо-
вании. Может быть, они проявляли также и состра-
дание друг к другу. На стоянке Шанидар в Ираке
только заботливые соплеменники могли долго уха-
живать за полуслепым, больным артритом стари-
ком с высохшей рукой, пока он не умер.
Но все же, подобно своим предшественникам, эти
люди, по-видимому, иногда ели друг друга, раска-
лывая черепа и кости конечностей, чтобы извлечь
мозг. Возможно, они надеялись унаследовать силу
тех, кого поедали.
Кроме каннибализма, очевидно, существовали и
другие ритуалы. В одной пещере неандертальцы рас-
положили кольцом рога горных козлов, в другой
сложили груду черепов пещерного медведя. Может
быть, они хотели умилостивить духов тех животных,
которых убивали.
Ритуальный характер, возможно, имело и искус-
ство неандертальцев, о котором мы знаем только по
таким предметам, как костяной амулет, испещ-
ренная царапинами галька, куски красной окиси
железа и растертый в порошок марганец, очевидно
применявшиеся для раскрашивания тела. (Возмож-
но, красный цвет символизировал кровь и, следова-
тельно, — жизнь.)
Культ медведя?
![]() Череп этого медведя лежал
Череп этого медведя лежал
в расщелине, отгороженной
от остальной части пещеры
камнями. Это позволяет сделать
предположение о наличии
зачатков религиозного ритуала.
Кости были найдены около реки
Ветерница в Югославии.
![]() Первыми свидетельствами о
Первыми свидетельствами о
появлении вполне современ-
ного человека, относящего-
ся к подвиду
Homo sapiens
sapiens,
были кости, найден-
ные в гроте Кро-Маньон на
юго-западе Франции. От
кроманьонцев остались ске-
леты и предметы материаль-
ной культуры во многих ча-
стях Европы. Здесь примерно
от
40 до
10 тысяч лет назад
жили люди, которые, вероят-
но, были предками совре-
менных людей европеоидной,
или кавказской расы. Охот-
ники позднего палеолита
смогли пережить суровые
невзгоды ледникового перио-
да, изготовляя новые, более
совершенные орудия из кам-
ня и кости и пользуясь ими
для охоты на крупных, даю-
щих много мяса животных,
которые бродили по конти-
ненту.
На смену этой эпохе пришел
мезолит
— среднекаменный
век, который длился от
10
до
5 тысяч лет назад.
На этом доисторическом
наскальном рисунке изображены
лучники, стреляющие
в бегущих оленей. Эта сцена
времен позднего палеолита или
эпипалеолита запечатлена на
стене пещеры Куэва-де-лос-
Кабаллос около Кастеллона
в Восточной Испании.
(Иллюстрация взята из книги
«История техники», изданной
под редакцией Сингера,
Холмйарда, Холла и Уильямса.)
![]()
Телосложение кроманьонца. О раннем современном человеке
Кроманьонский человек был,
![]()
![]()
![]()
![]()
|
|
|
|
|
|
|
Новые приемы обработки
камня и кости
С появлением первых вполне современных людей
палеолит (древний каменный век) вступил в позд-
нюю, или новую, стадию развития — период своего
наивысшего расцвета. В Европе он продолжался от
35 до 10 тысяч лет назад и совпал со временем по-
следнего плейстоценового похолодания.
Как и в мустьерских культурах неандертальцев,
в культурах кроманьонцев позднего палеолита пре-
обладали каменные инструменты и оружие, пред-
назначенные для охоты. Но имелись также и удиви-
тельные новшества, вероятно, заимствованные у
культур Ближнего Востока.
Во-первых, кроманьонцы разработали новый очень
эффективный способ изготовления каменных лезвий.
Прижав к краю удлиненной каменной заготовки
долото, изготовленное из отростка оленьего рога,
мастер ударял по нему молотком, сделанным из
камня, кости, дерева или рога. В результате этого
направленного под углом удара от заготовки отка-
лывался узкий плоский отщеп с острой кромкой. На-
давливая на этот отщеп в надлежащих местах ост-
рыми каменными клиньями, можно было получить
другие, более мелкие, отщепы. С помощью этих спо-
собов из одного камня удавалось изготовить значи-
тельно большее количество тщательно отделанных
орудий, чем это позволяли ранее применявшиеся
технологии.
По сравнению со своими предшественниками —
неандертальцами, кроманьонцы производили значи-
тельно более широкий круг более тщательно изго-
товленных ножей, скребков, пил, наконечников,
сверл и других каменных орудий. Около половины
всех инструментов кроманьонцев было сделано из
кости, которая прочнее и долговечнее дерева. Ка-
менные резцы применялись также для изготовления
изделий из рога, дерева и слоновой кости.
Из этих материалов кроманьонцы делали и такие
новые орудия, как иглы с ушками, крючки для ловли
рыбы, гарпуны и копьеметалки. Вместе взятые все
эти кажущиеся простыми орудия в огромной степени
усилили власть человека над природой.
Обработка камня методом
отжима.
Прижимая заостренное орудие
к внешнему краю кремневой
заготовки, откалывали мелкие
отщепы с ее нижней стороны.
а — Заостренная палка или
кость.
![]()
![]() б — Обрабатываемое изделие.
б — Обрабатываемое изделие.
в — Прокладка из коры на
каменной рабочей плите
(наковальне).
Орудия позднего палеолита.
![]() Справа показаны четыре
Справа показаны четыре
кремневых и одно костяное
орудие (масштабы не
выдержаны). Изображения
других орудий можно увидеть
на следующих страницах.
а — Кремневый нож, тыльная
сторона которого обработана
методом отжима.
б — Кремневое скребло,
округленное с одной стороны
методом отжима.
в — Долотообразный резец
для обработки рога, кости или
дерева.
г — Маленькое сверло для
прокалывания отверстий в коже,
дереве, кости или роге.
д — Костяная игла с ушком,
проколотым маленьким сверлом.
|
|
Экономия камня.
![]() А — Длина режущего края на
А — Длина режущего края на
1 фунт камня, которую можно
было получить при обработке
орудий с помощью технологии,
применяемой неандертальцами.
Б — Длина режущего края на
I фунт камня при обработке
орудий приемами кроманьонцев.
Успехи в технике обработки
камня.
![]() Более 250 ударов (включая
Более 250 ударов (включая
и отжимы) требовалось
кроманьонцам для изготовления
тщательно отделанного
кремневого ножа. Сравните это
с тем числом ударов, которые
затрачивали на производство
орудий неандерталец и Homo
erectus.
11 Зак. 5116.
Культурные комплексы.
Четырьмя отрезками линий
показано время существования
четырех культурных
комплексов эпохи позднего
палеолита в Западной Европе.
(Мы опускаем краткосрочную
культуру азиль, которая
началась после оледенения
около 10 тысяч лет до н. э.;
некоторые считают ее
послепалеолитической культурой
мезолита.)
1.
Перигор; мы включаем сюда
и поздний перигор, или
граветтийскую культуру,
которая началась по крайней
мере 29 тысяч лет назад.
2. Ориньяк.
3. Солютре.
4. Мадлен.
Набор орудий, применявшихся европейскими
кроманьонцами, менялся в зависимости от времени
и места. Некоторые археологи думают, что теми
или иными орудиями пользовались разные группы
людей. Другие специалисты доказывают, что эти на-
боры орудий просто представляют различные школы
мастерства.
На основании орудий и других предметов выде-
ляют пять основных культур позднего палеолита
для Западной Европы; их остатками особенно бо-
гата Юго-Западная Франция. (В прочих регионах
мира имеется много других культур.)
Для перигорской культуры (Франции) возрастом
примерно 35—32 тысячи лет характерны скребла,
пилы и ножи с округлой тыльной стороной, кото-
рые, возможно, берут начало еще в мустьерских тра-
дициях и продолжают изготовляться во времена
шательпетронской культуры неандертальцев.
Более поздняя — граветтийская — культура с
длинными сужающимися остроконечниками типа
граветт, с многочисленными резцами и костяными
орудиями имела широкое географическое распро-
странение — от Франции до России, где она суще-
ствовала еще 10 тысяч лет назад.
От ориньякской культуры (примерно 35—29 тысяч
лет назад) сохранилось много скребел, сверл и до-
лот, но она была сравнительно бедна резцами типа
ножей. На Ближнем Востоке встречались анало-
гичные технологии даже в более древнюю эпоху.
В недолговечной культуре солютре (около 22—
18,5 тысячи лет назад) особое внимание уделялось
высокому мастерству в изготовлении таких пред-
метов, как тонкие листовидные остроконечники, за-
остренные с обоих концов и тщательно обработанные
с обеих сторон.
Культура мадлен (около 18,5—11 тысяч лет назад)
известна стержнями с крюком, предназначенными
для метания копий, и орудиями рыбной ловли от
зазубренных крючков до гарпунов.
|
|
Орудия кроманьонцев.
Показаны орудия пяти культур
кроманьонцев (масштабы
не соблюдены).
1. Перигор.
а — Скребло.
б — Резец.
в — Нож с округлой тыльной
стороной.
2.
Граветтийская культура.
г — Сужающийся
остроконечник.
д — Резец.
е — Нож с тупой тыльной
стороной.
3. Ориньяк.
ж — Скребло с выступом.
з — Заостренное лезвие.
и — Резец.
4. Солютре.
к — Наконечник стрелы.
л — Листовидный
остроконечник,
м — Проколка.
5. Мадлен.
н — Гарпун из оленьего рога.
о — Костяной рыболовный
крючок.
![]() п — Наконечник из оленьего
п — Наконечник из оленьего
рога.
|
|
| to,*,#/™*.'/»*'»"* |
| Последовател ьность Схематический разрез, последовательность культурных слоев, установленных во Франции. а — Пустые слои. б — Мадлен. в — Солютре. г — Ориньяк. д — Мустьерская культура |
Археологические находки говорят о том, что ору-
жие и способы охоты кроманьонцев были значительно
совершеннее, чем у неандертальцев; это имело ог-
ромное значение для увеличения пищевых ресур-
сов и роста населения.
![]() Копьеметалки, напоминающие по внешнему вицу
Копьеметалки, напоминающие по внешнему вицу
огромные крючки для вязания тамбуром, давали
человеческой руке выигрыш в силе, удваивая рас-
стояние, на которое охотник мог метнуть свое копье.
Теперь он был способен поразить добычу на боль-
шом расстоянии еще до того, как она успеет испу-
гаться и убежать. В числе зазубренных наконечни-
ков был изобретен гарпун, которым можно было
добывать лосося, идущего из моря вверх по реке ме-
тать икру. Рыба впервые стала важным продуктом
питания, особенно в Юго-Западной Франции. Ка-
навки, выдолбленные в наконечниках копий, уве-
личивали сток крови у раненого животного, ускоряя
его гибель. Кроманьонцы ловили птиц в силки; поч-
ти нет сомнения, что именно они придумали смер-
тельные ловушки для птиц, волков, лис и значи-
тельно более крупных животных. Некоторые спе-
циалисты считают, что именно в такую западню по-
пала та сотня мамонтов, чьи остатки были найдены
около Павлова в Чехословакии.
| а ----------------- |
^ й. |
| 6 — |
Охота с копьеметалкой.
1.
Так использовал копьеметалку
кроманьонский охотник
(справа внизу).
а — Копьеметалка.
б — Копье.
2.
На верхнем рисунке видно,
насколько это увеличивало
дальность метания копья.
![]()
![]() а — Обычно охотник может
а — Обычно охотник может
метнуть длинное копье на 70
ярдов (64 м); фактически
расстояние, на котором можно
поразить добычу, составляет
15 ярдов (13,7 м).
б — Копьеметалка помогает
метнуть копье на 150 ярдов
(137 м); с ее помощью можно
поразить жертву на расстоянии
30 ярдов (27,4 м).
Раненый бизон.
На этом позднепалеолитическом
пещерном рисунке из пещеры
Нио в Юго-Западной Франции
изображен бизон, пораженный
стрелами. Но кроме таких
изображений и нескольких
найденных наконечников стрел,
имеется мало доказательств
того, что до окончания
ледникового периода охотники
широко пользовались луком
и стрелами.
Кроманьонцы были, возможно, самыми искус-
ными из когда-либо существовавших охотников на
крупного зверя. Сезонное проживание на избранных
участках говорит о том, что они преследовали север-
ного оленя и каменного козла (козерога) во время
их сезонных миграций на новые пастбища и обратно.
Огромные груды костей свидетельствуют о том, что
охотники научились загонять стада на те участки, где
животных было легко забить. Этим можно объяснить
находку тысячи бизонов, очевидно, убитых в одном
из оврагов Южной России. Еще большее впечатле-
ние производят скелеты 10 тысяч диких лошадей,
которые были найдены под скалой около Солютре
на востоке Центральной Франции. Около 17 тысяч
лет назад охотники, очевидно, загоняли целые стада
на вершину этой скалы, ловили и убивали их у се под-
ножия.
Используя подобные приемы и орудия, кромань-
онцы получили почти неиссякаемый источник высо-
копитательной пищи. Это, очевидно, способствовало
их размножению и помогло им заселить даже суро-
вые холодные районы Сибири.
Миграция оленьих стад (внизу).
Стрелками показаны возможные
пути весенней миграции
северных оленей из районов
с мягкой зимой на прибрежные
или горные пастбища.
а — Перигор.
б — Побережье Атлантического
океана (в настоящее время этот
район затоплен морем).
в — Кантабрийские горы.
г — Пиренеи.
![]() д — Центральный массив.
д — Центральный массив.
е — Средиземноморское
побережье (сейчас этот район
затоплен морем).
ж — Альпы.
|
|
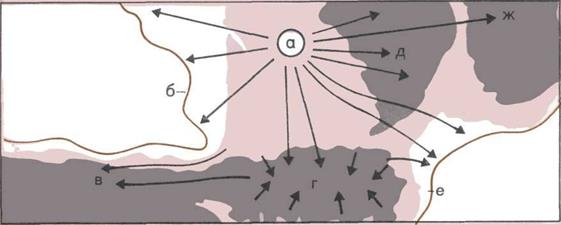
Колебания численности видов
животных.
Диаграммы показывают, как
возрастала и уменьшалась
численность некоторых видов
животных и растений на юго-
западе Франции по мере
колебания температуры в конце
плейстоцена.
а — Изменение температуры
за период 30—10 тысяч лет до
н. э.
б — Благородный олень.
в — Песец.
г — Лиственные деревья.
д — Хвойные деревья.
Европа позднего каменного века кишела крупными
дикими млекопитающими, от которых можно было
получить много мяса и мехов. После этого никогда
уже их количество и разнообразие не были столь ве-
лики. Многие виды животных были, вероятно, из-
вестны уже неандертальцам. Обильные, широко рас-
пространенные следы охоты кроманьонцев проли-
вают новый свет на то, какая была в то время дичь
и где она водилась.
Дикие лошади, северные олени, бизоны, мамон-
ты, сайгаки и шерстистые носороги паслись в откры-
той степи. На лесистых склонах и в долинах укрыва-
лись благородные олени, дикие вепри, пещерные
медведи и пещерные волки. Серны, каменные козлы
и дикие козы забирались высоко в горы на летние
пастбища. Дикие туры изобиловали в степях и в при-
мыкающих к ним лесистых местностях.
По находкам костей животных устанавливается,
что в начале позднего палеолита на территории Се-
верной Германии преобладали северный олень и ди-
кая лошадь, на юге Германии — мамонты и носоро-
ги, а далее к востоку, по берегам Черного моря, —
быки и лошади. Но состав животного мира не был
постоянен. Так, на одной из итальянских стоянок
основными объектами охотничьего промысла были
сначала дикие быки, лошади, благородные олени
и кабаны, затем среди добычи на первое место выхо-
дят лошади и дикие ослы, которых в свою очередь
сменяют кабаны, северные олени и быки. Не исклю-
чено, что на численность различных групп живот-
ных оказывали влияние периодические изменения
климата.
|
|
Вот пять видов животных, которые были основны-
ми источниками питания кроманьонцев.
1. Rangifer tarandus, северный олень; это един-
ственный вид оленя, у которого и самцы и самки
имеют рога. Высота в холке — до 4,6 футов (1,4 м).
Время — от плейстоцена до наших дней. Место —
Заполярье.
2. Cervus elephas, благородный олень. Высота
в холке — до 5 футов (1,5 м). Время — от плейсто-
цена до наших дней. Место — Евразия и Северная
Африка.
3. Bos prlmigenlus, тур — огромный бык. Высота
в холке 6 футов (1,8 м). Время — от плейстоцена до
1627 г. н. э. Место — Евразия и Северная Африка.
4. Equus caballus, лошадь; маленький дикий под-
вид современной домашней лошади. Высота в хол-
ке — до 4,6 футов (1,4 м). Время — от плейстоцена
до наших дней. Место — континенты Северного по-
лушария.
5. Capra pyrenaica, горный подвид пиренейского
каменного козла; у самцов длинные рога, загнутые
вперед и вверх. Высота в холке — до 30 дюймов
(76 см). Время — от плейстоцена до наших дней.
Место — Испания и одно время Юго-Западная Фран-
ция.
Пять объектов охоты.
Нумерация соответствует
принятой в тексте.
1. Северный олень.
2. Благородный олень.
3. Тур.
4. Лошадь.
5. Горный пиренейский козел.
|
|
Одежда для зашиты от холода.
![]() По вырезанным из кости
По вырезанным из кости
статуэткам, найденным в России,
можно судить о том, как люди
защищались от холода во время
зим ледникового периода.
а — Статуэтка человека,
одетого в плотно облегающий
меховой костюм со штанами
и капюшоном. Эта статуэтка,
вырезанная из бивня мамонта,
была найдена на стоянке
Буреть в Сибири.
б — Статуэтка без головы,
изображающая женщину,
одетую в нарядный костюм
с поясом. Найдена на стоянке
Костенки к югу от Москвы.
В строительстве кроманьонцы в основном сле-
довали старым неандертальским традициям, однако
в некоторых их жилищах и, возможно, во всей одеж-
де имелись существенные нововведения, которые
способствовали выживанию в условиях последнего
холодного тысячелетия плейстоцена.
Как и их предшественники — неандертальцы,
которые жили в пещерах, европейские кроманьонцы
пользовались известняковыми приречными утесами в
Юго-Западной Франции, особенно в Дордони, а так-
же и на северных склонах Пиренеев. Многие пещеры
имели выход на юг, обогревались солнцем и были
защищены от холодных северных ветров. Пещеры
находились рядом с обильными источниками воды,
и оттуда открывался хороший вид на пастбища, где
паслись тучные стада травоядных животных. В тех
местах, где пища имелась в изобилии круглый год,
в одной большой пещере могло постоянно жить до
нескольких десятков человек. Но в некоторых пеще-
рах мы видим только сезонные следы пребывания
человека.
И опять же подобно неандертальцам, представи-
тели граветтийской культуры в России строили зим-
ние жилища в речных долинах. Одни жилища были
сложены из камня, другие вырыты в земле. У многих
стены и крыша были сделаны в виде шатра из шкур,
подпертых бедренными костями мамонта и закреп-
ленных внизу по краям другими тяжелыми костями
и бивнями. К крупнейшим сооружениям относится
продолговатое жилище длиной 89 футов (27 м) со
стоянки Костенки в долине реки Дон; ряд очагов,
идущих по центру, говорит о том, что здесь зимовали
несколько семей под одной крышей, состоявшей из
нескольких покрытых шкурами шатров.
По находкам и старым рисункам во Французских
пещерах видно, что кочующие охотники возводили
также легкие летние шалаши, подобные тем, кото-
рые и сейчас строят некоторые современные люди,
ведущие примитивный образ жизни.
Возможностью жить и трудиться в условиях лед-
никового периода кроманьонцы были обязаны но-
вым видам одежды не в меньшей степени, чем но-
вым методам строительства и использованию огня.
Костяные иглы и изображения одетых в мех людей
говорят нам о том, что они носили сохранявшие теп-
ло тела тесно прилегающие штаны, парки с капюшо-
нами, обувь и рукавицы с хорошо прошитыми шва-
ми.
История одной пешеры.
1.
На плоскогорье при
просачивании воды растворился
слой известняка и образовалась
подземная пещера с выходом
в реку.
2.
После понижения уровня
воды в реке пещера осушилась,
и в ней поселились кроманьонды.
3.
В настоящее время вход
в пещеру закрыт обвалом,
карстовые воронки в ее кровле
заросли деревьями, и остатки
кроманьонцев оказались
захороненными в слоях осадков,
покрывающих днище пещеры.
Длинное жилище.
![]() Это реконструкция длинного
Это реконструкция длинного
коммунального жилища, пол
которого был слегка погружен
в землю; крышей служили
шкуры, подпертые шестами;
низ стен был укреплен костями
и бивнями мамонта. Стоянка
обнаружена близ села Пушкари
к северо-востоку от Киева. Это
сооружение могло состоять из
трех выстроенных в один ряд
и соединенных между собой
шатров.
|
|

Фазы Луны? (внизу).
а — Костяная пластинка из
Сержака во Франции (размеры
несколько уменьшены).
б — Символы,
![]() выгравированные на пластинке,
выгравированные на пластинке,
могут изображать
последовательные фазы Луны.
В эпоху с 35 до 10 тысяч лет назад Европа пере-
жила великий период своего доисторического ис-
кусства. Круг произведений был широк: гравюры
животных и людей, сделанные на небольших кусоч-
ках камня, костей, слоновой кости и оленьих рогов;
глиняные и каменные скульптуры и рельефы; рисун-
ки охрой, марганцем и древесным углем, а также
изображения, выложенные на стенах пещер мхом
или нанесенные краской, выдутой через соломинку.
Исследование более 100 пещер и скальных укры-
тий привело некоторых ученых к выводу, что искус-
ство кроманьонцев прошло четыре стадии развития.
Для первого • периода (32—25 тысяч лет назад)
характерны изображения животных и других объек-
тов, большей частью плохо нарисованные на малень-
ких предметах, которые люди носили с собой. Ко
второму периоду (25—19 тысяч лет назад) относится
ранее пещерное искусство, включая отпечатки рук,
а также гравированные и нарисованные силуэты
животных с дугообразно изогнутыми спинами. Тре-
тий период (19—15 тысяч лет назад) явился вер-
шиной пещерного искусства, что можно видеть по
прекрасно выполненным, динамичным рисункам
лошадей и туров в пещере Ляско на юго-западе
Франции и по другим образцам рельефной скульп-
туры. Для четвертого периода (15—10 тысяч лет
назад) особенно характерны изображения на мел-
ких предметах, а также символические знаки и вели-
колепно выполненные в реалистической манере
изображения животных в пещерах Альтамира в Се-
верной Испании и Фон-де-Гом во Франции.
Большинство изображений находятся глубоко в
пещерах, где художники, очевидно, работали при
свете горящих поленьев или ламп с фитилями из
мха. Возможно, их искусство имело ритуальное зна-
чение. Некоторые рисунки изображают фигуры по-
лулюдей — полуживотных, предположительно кол-
дунов или шаманов. Многие сцены с изображением
тучной охотничьей добычи, возможно, объясняются
охотничьей магией или сексуальным символизмом.
Символика плодородия, вероятно, воплощена в че-
ловеческих фигурках с утрированными женскими
признаками. Геометрические узоры могут быть услов-
ными системами обозначений, одна из которых, воз-
можно, изображает фазы Луны. Но все эти интерпре-
тации носят дискуссионный характер.
|
|

![]()
СГоянки Западной Европы,
на которых были обнаружены
предметы палеолитического
искусства.
На этой карте Западной
Европы эпохи позднего
палеолита показаны ледниковые
покровы, а также древняя
и современная береговые линии.
Точками обозначены находки
образцов пещерного искусства,
сосредоточенные главным
образом на севере Испании
и на юго-западе Франции. Это
наскальные гравюры, статуэтки
и другие предметы,
встречающиеся также в более
северных и восточных районах.
К стоянкам, на которых были
найдены предметы искусства,
представляющие особую
ценность, относятся:
1. Альтамира.
2. Нио.
3. Пещ-Мерль.
4. Ляско.
5. Монгодье.
Шедевры искусства
кроманьонцев.
Три находки из Франции,
1.
Вырезанное из рога северного
оленя изображение бизона,
лижущего свою спину.
2.
Наскальный рисунок лошади,
окруженный точками
и отпечатками ладоней рук,
из пещеры Пеш-Мерль.
3.
Тюлени, выгравированные
на жезле из рога, обнаруженном
в Монгодье. Такие жезлы
![]()
![]() с отверстием, возможно, служили
с отверстием, возможно, служили
для обстругивания копий.
Два ожерелья эпохи позднего
палеолита, погребенные вместе
со своими владельцами,
свидетельствуют о
существовании личной
собственности.
1.
Ожерелье из костяных бусин
и подвесок с отверстиями;
погребено вместе с ребенком
в Сибири.
2.
Фрагмент трехрядного
ожерелья: два ряда состоят
из рыбьих позвонков
с отверстиями, а один — из
просверленных раковин, причем
все три ряда соединены зубами
оленя, в которых тоже
проделаны отверстия. Ожерелье
было погребено вместе с юношей
на юго-востоке Франции.
Так называемые «негры»
из Гримальди.
Эти скелеты были найдены
в отдельном, залегающем
на небольшой глубине
погребении внутри «Грота
детей» — пещеры в Италии.
Деформация костей черепа
после смерти вызвала
прогнатизм (выступание
челюстей вперед); это привело
некоторых специалистов
к неверному выводу о том, что
здесь найдены непосредственные
предки современных негров.
А — Юноша ростом 5 футов
1,5 дюйма (1,56 м).
![]() Б — Скелет старой женщины
Б — Скелет старой женщины
ростом 5 футов 3 дюйма
(1,6 м).
Захоронения, находящиеся в столь удаленных
друг от друга местах, как Франция и Россия, дают
ключ к пониманию жизни кроманьонцев.
Изучение скелетов говорит о том, что две трети
кроманьонцев достигали 20-летнего возраста, тогда
как у их предшественников — неандертальцев число
таких людей не составляло и половины; один из
десяти кроманьонцев доживал до 40 лет по срав-
нению с одним человеком на двадцать у неандер-
тальцев. Хотя эти цифры основаны на небольшом
количестве образцов, исходя из них можно думать,
что продолжительность жизни возрастала.
С большей уверенностью по погребениям кромань-
онцев можно судить о их символических ритуалах
и росте богатства и социального статуса. Хоронив-
шие часто посыпали мертвых красной охрой, кото-
рая, как предполагают, символизировала кровь и
жизнь, что, может быть, свидетельствует о наличии у
кроманьонцев веры в загробное существование.
Некоторые трупы были захоронены с богатыми ук-
|
|
рашениями; это ранние признаки того, что в сооб-
ществах охотников-собирателей начали появляться
богатые и уважаемые люди.
Вот всего лишь несколько примеров захоро-
нений.
В кроманьонском укрытии под'скалой около Лез-
Эйзи в Юго-Западной Франций находились пять
взрослых и один ребенок. Их останки были окрашены
красной охрой и погребены вместе с ориньякскими
орудиями из кремня и с морскими раковинами, в
которых проделаны отверстия.
В Гримальди на Итальянской Ривьере найдено
16 человек, похороненных в различных пещерах око-
ло 30 тысяч лет назад. В пещере Грот-дю-Кавийон
был найден мужчина с короной из оленьих зубов
и с головным убором, украшенным сотнями рако-
вин моллюсков. В пещере «Грот детей» были по-
гребены два ребенка с украшениями из раковин
моллюсков-трубачей. В Барме-Гранде найдены
останки людей с ожерельями из позвонков живот-
ных.
В Пржедмости в Чехословакии было захоро-
нено 29 человек вместе с глиняными фигурками и
орудиями из рога и кости.
![]() Возможно, самые удивительные вещи найдены в
Возможно, самые удивительные вещи найдены в
захоронении охотников, сделанном 23 тысячи лет
назад в Сунгири к востоку от Москвы. Здесь лежал
старик в меховых одеждах, искусно украшенных
бусами. Поблизости были захоронены два мальчика,
одетые в украшенные бусинами меха, с кольцами и
браслетами из слоновой кости; около них лежали
длинные копья из бивней мамонта и два странных,
вырезанных из кости и похожих на скипетры стержня
того типа, который называют batons de comman-
dement (жезл командующего).
Скелет, покрытый украшениями
(слева).
![]() Более тысячи бусин и других
Более тысячи бусин и других
украшений покрывали меховые
одежды старика, скелет которого
был раскопан в 1955 г.
в погребении Сунгири. Меха
истлели много веков назад.
Добыча охотников периода
мезолита.
На схеме показано, на скольких
из 165 европейских стоянок были
найдены кости этих
мл скоп итаю ш их, являвшихся
основной добычей охотников
периода мезолита.
1. Благородный олень.
2. Кабан.
3. Дикий бык.
4. Косуля.
5. Лиса.
6. Барсук.
7. Дикая кошка.
8.
![]() Бобер.
Бобер.
10 тысяч лет назад холодная эпоха плейстоцена
уступила место голоцену, или «совершенно новой»
эпохе. Это время мягкого климата, в которое мы
живем и сейчас. По мере потепления климата Евро-
пы территория, занятая лесами, расширялась. Леса
наступали, занимая огромные участки бывшей тунд-
ры, а море, уровень которого повышался, затопляло
низкие побережья и долины рек. Климатические
изменения и усиленная охота привели к исчезнове-
нию огромных диких стад, за счет которых корми-
лись кроманьонцы. Но на суше оставались в обилии
лесные млекопитающие, а в воде — рыба и водо-
плавающая дичь.
Инструменты и оружие, изображенные на следую-
щих двух страницах, позволили северным европей-
цам использовать указанные источники пищи. Эти
специфические группы охотников и собирателей и
создали культуру мезолита, или «среднего камен-
ного века». Он был так назван, потому что последо-
вал за древним каменным веком, который характе-
ризовался охотой на стада животных и предшество-
вал появлению в Северной Европе земледелия, свой-
ственного для нового каменного века. Продолжав-
шийся только от 10 до 5 тысяч лет назад мезолит
явился лишь кратким мгновением доисторического
периода. По костям, найденным на мезолитических
стоянках, видно, что добычей охотников мезолита
были благородный олень, косуля, кабан, дикие быки,
бобры, лисы, утки, гуси и щуки. Огромные кучи ра-
ковин моллюсков говорят о том, что ими питались на
побережье Атлантики и Северного моря. Люди мезо-
лита занимались также сбором корней, плодов и
орехов. Группы людей, очевидно, мигрировали с
места на место, следуя за сезонными изменениями
источников пищи.
|
|
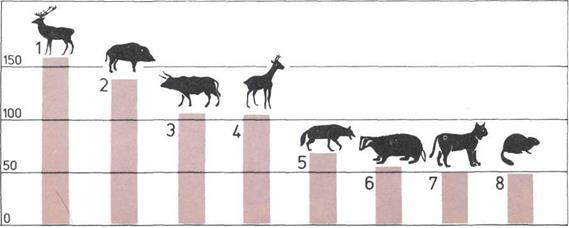
|
|
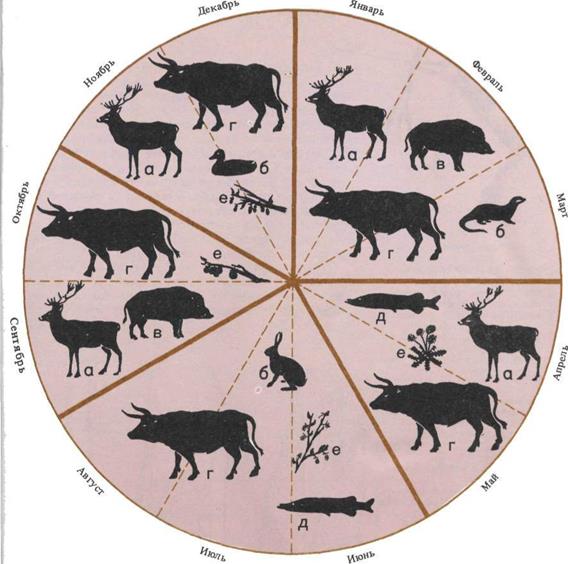
Археологи считают, что люди эпохи мезолита жили
меньшими группами, чем их возможные предки —
кроманьонские охотники на крупного зверя. Но до-
быча пищи держалась теперь на более устойчивом
уровне в течение круглого года, в результате чего
количество стоянок и, следовательно, численность
населения возрастали. Продолжительность жизни
тоже, по-видимому, увеличилась. По нескольким
десяткам скелетов взрослых людей, найденных на
реке Дунай в Югославии, можно судить о том, что
если удавалось выжить в детстве, женщины в этом
благодатном месте достигали в среднем 35 лет, а
мужчины — 50 лет. Немногие из людей палеолита
жили столь долго.
Годовой цикл питания.
Годовой цикл смены продуктов
питания у населения северо-
запада Европы 8 тысяч лет
назад делился на пять основных
сезонов. Мясом оленя и быка
питались круглый год, но
количество этих животных
изменялось в зависимости
от сезона.
а — Благородный олень.
б — Мелкая дичь.
в — Кабан.
г — Дикий бык.
д — Рыба.
е — Растения.
![]()
![]()
|
|
|
|
|
|
|
|
|
или трапеции. Микролиты приклеивались смолой к
древку из оленьей кости, и их можно было легко
заменить, если они отваливались или притуплялись.
Кроме того, изготовление микролитов сберегало
материал: опытный мастер мог создать 300 футов
(92,5 м) режущей поверхности всего лишь из 2 фун-
тов (987 г) кремня.
![]()
![]() Новые образцы крупных каменных орудий помо-
Новые образцы крупных каменных орудий помо-
гали людям эпохи мезолита изготовлять челноки,
весла, лыжи и сани. Все это вместе взятое позволило
освоить огромные водные участки для ловли рыбы
и облегчило передвижение по снегу и заболоченной
местности.
Изготовление микролитов
(внизу слева).
1. Лезвие из кремня.
2. Лезвие с выемками
на противоположных сторонах.
3.
Лезвие, расколотое на три
части.
4.
Центральная часть лезвия,
которой придана форма
ромбовидного микролита.
5.
Лезвие, имеющее выемки
с одной стороны.
6.
Лезвие, расколотое на три
части.
7.
Центральная часть лезвия,
отделанная в форме трапеции
(а), полумесяца (б),
треугольника (в).
Т ипы микролитов.
Эти орудия представляют
четыре различные культуры
периода мезолита.
1—3. Азильская культура.
названная по пещере Мас
д'Азиль во Французских
Пиренеях.
4—6. Маглемозианская, или
лесная, культура, названная по
стоянке Маглемоз в
голландской провинции
Зеландия;
7—9. Совтеррианская культура,
названная по стоянке
Совтерр-ля-Леманс во Франции,
но широко распространенная
и в других районах Западной
Европы.
![]() 10—12. Тарденуазская
10—12. Тарденуазская
культура, названная по стоянке
близ городка Фер-ан-Тарденуа
в Северной Франции.
![]() СОВРЕМЕННЫЙ
человек
СОВРЕМЕННЫЙ
человек
ОСВАИВАЕТ ЗЕМЛЮ
![]() Появление нашего подвида
Появление нашего подвида
людей в Европе совпало с его
быстрым распространением в
Африке, Азии и Австра-
лии. А произошел современ-
ный человек, вероятно, от
одной из архаичных форм
Homo sapiens,
жившей в Аф-
рике
100 тысяч лет назад.
В этой главе дан краткий
обзор ранних находок остан-
ков современного человека
во всем мире и показано, как
местные различия в климате
способствовали той диф-
ференциации рас, которую
мы наблюдаем в настоящее
время.
Некоторые из аборигенов
Австралии охотятся, пользуясь
копьями точно так же, как это
делали наши предки в эпоху
палеолита. Человек появился
в Австралии, вероятно, более
40 тысяч лет назад, когда люди
современного типа колонизовали
все континенты Старого Света.
(Фотография предоставлена
Австралийским бюро новостей
и информации.)
![]()
Одна из теорий происхождения
человеческих рас.
![]() Выше дана схема происхождения
Выше дана схема происхождения
человеческих рас, полученная
на основе исследования белков
и молекул ДНК, содержащихся
в хромосомах.
а — Австралоиды.
б — Монголоиды.
в — Негроиды.
г — Европеоиды.
Другая теория происхождения
рас.
На основе исследования ДНК
митохондрий была создана
другая теория происхождения
рас.
а — Большинство австралийских
аборигенов.
![]()
![]() б — Большинство монголоидов.
б — Большинство монголоидов.
в — Негроиды.
г — Европеоиды.
д — Некоторые из монголоидов.
е — Некоторые из
австралийских аборигенов.
Происхождение современного
человека
К 80-м годам нашего века стало казаться все бо-
лее маловероятным, что европейские кроманьонцы
произошли от неандертальцев или были первыми
вполне современными людьми в мире. Скептики ут-
верждают, что любой эволюционный скачок от не-
андертальца к кроманьонцу должен был бы совер-
шиться слишком быстро, судя по тем ископаемым
находкам, которыми мы располагаем. Большинство
из них полагают, что наш подвид произошел от ар-
хаичной формы Homo sapiens в Африке около 100
тысяч лет назад.
«Современные» ископаемые черепа действительно
позволяют нам думать, что наши предки вышли имен-
но из Африки (см. с. 182—183). Другие свидетель-
ства этого можно найти в нас самих в виде генов,
унаследованных различными расовыми группами.
Когда-то ученые классифицировали эти группы по
таким ненадежным внешним признакам, как тело-
сложение и цвет кожи. Позже биологи начали срав-
нивать уже такие более устойчивые особенности,
как группы крови и связанные с ними протеины. Но
различные последовательности нуклеотидов в ДНК
могут создавать гены, состоящие из одних и тех же
протеиновых веществ. Поэтому самый надежный спо-
соб, позволяющий проследить долгосрочные унасле-
дованные связи групп людей, заключается в сравне-
нии самих последовательностей нуклеотидов в ДНК.
Такое сравнение и выполнили биологи.
В 1986 г. английские ученые Джеймс Уэйнскот
и Адриан Хилл сообщили о возможности использо-
вания биологических веществ, называемых энзи-
мами, для отделения пяти фрагментов ДНК от гена,
отвечающего за получение части молекулы гемогло-
бина красных кровяных телец. Они установили раз-
личные сочетания этих фрагментов у 600 человек,
в число которых входили африканцы, англичане,
индусы, меланезийцы и таиландцы. Оказалось, что
не-африканцы имеют ограниченное число общих
сочетаний, тогда как наиболее характерным для
африканцев является сочетание, отсутствующее у
других групп. Ученые сделали вывод, что все наро-
ды мира происходят от одной маленькой группы до-
исторических африканцев, связанных между собой
кровными родственными узами.
Многие сторонники этой теории считают, что че-
ловек появился в Африке 100 тысяч лет назад, не-
сколько позднее распространился в Азии и 30 тысяч
лет назад вступил на все континенты, за исключе-
нием Антарктиды.
Американские ученые Ребекка Канн и Марк Стоун-
кинг пришли к аналогичным выводам, изучая гео-
![]()
|
|
|
|
|
|
|
|
|
|
|
|
|
|
графическое распределение генов митохондрий,
передаваемых только по женской линии. Их выводы
состоят в том, что наш подвид происходит от одной
женщины, которая жила в Африке 200 тысяч лет
назад — значительно раньше, чем об этом можно
судить по ископаемым остаткам.
![]() Дальнейшие генетические исследования должны
Дальнейшие генетические исследования должны
пролить новый свет на наше не вполне ясное про-
исхождение.
Данные получены на основе
изучения фрагментов ДНК
у 600 человек, о которых идет
речь в нашем тексте. Чем меньше
общих символов, относящихся
к каким-либо двум группам
населения, тем меньше их
генетическая удаленность и тем
ближе предполагаемое родство.
Б — Здесь представлено
эволюционное древо этих восьми
групп населения.
Распределение серповидных
клеток.
На этой карте показано
современное распространение
серповидных красных кровяных
телец и малярии среди
различных человеческих
популяций. Серповидные клетки
появились в результате мутации
у одной особи. Ген, отвечающий
за образование серповидных
клеток, повышает
сопротивляемость человека
к заболеванию малярией, но
вызывает анемию в том случае,
если он унаследован от обоих
родителей.
1
— Распространение
серповидных клеток.
2 — Распространение малярии.
Стоянки и центры обитания
человека современного типа
в Африке.
1. Фиш Хек.
2. Устье реки Клазис.
3.
Флорисбад
(местообитание архаичной
формы).
4. Пещера Бордер.
5.
Местообитание современных
бушменов.
6. Мумбва.
7. Лукение-Хилл.
8. Элментейта.
9. Ишанго.
10. Омо.
11.
Предполагаемый центр
происхождения негров.
12.
![]()
![]() Афалу-бу-Руммель.
Афалу-бу-Руммель.
В Африке были найдены более древние ископае-
мые остатки, чем на каком-либо из других конти-
нентов. Так, в Южной Эфиопии нашли Омо-1 —
неполный череп со многими современными призна-
ками, возраст которого, вероятно, более 60 тысяч лет.
В устье южноафриканской реки Клазис обнару-
жены «современные» остатки, возраст которых 100
тысяч лет, а в пещере Бордер найдена «современ-
ная» нижняя челюсть возрастом 90 тысяч лет.
Некоторые другие древние остатки можно увя-
зать с современными неграми и бушменами. Пред-
полагается, что наследственные особенности, ха-
рактерные для этих групп, обнаруживаются в чере-
пе из Флорисбада (Южная Африка), которому более
100 тысяч лет. Но специфические, похожие на дет-
ские черепа бушменского типа имеют большую древ-
ность, чем первые известные нам остатки негров
давности 17 тысяч лет. По всей видимости, бушме-
Современные охотники-
собиратели.
Подобно своим предкам эпохи
палеолита, некоторые бушмены
до сих пор охотятся с помощью
лука и стрел и питаются в
основном дикими растениями.
но-готтентотская группа когда-то занимала всю
Южную Африку. Группы бушменов охотников-со-
бирателей и сейчас еще ведут почти палеолитиче-
ский образ жизни в пустыне Калахари на территории
Ботсваны.
На рисунках приведены изображения древних
и ныне живущих африканцев.
1. Омо-1; тонкокостный неполный череп, с выпук-
лой верхней частью, округлым затылком, с неболь-
шими надглазничными валиками, с хорошо разви-
тым подбородком; объем мозга 1400 см . Некоторые
специалисты относят его к архаичной разновидно-
сти sapiens. Возраст — свыше 60 тысяч лет. Место —
река Омо, Эфиопия.
2. Череп из Флорисбада; крупный череп с низ-
ким сводом, надбровными валиками, с выступаю-
щими вперед челюстями и низко посаженными пря-
моугольными глазными впадинами. Возраст — свы-
ше 100 тысяч лет. Место — Флорисбад, Южная Аф-
рика.
3. Бушмен; раса современного человека. Невы-
сокие стройные жилистые люди с небольшим лицом
и высокими скулами, плоским широким носом и вы-
ступающим вперед лбом. Время — возможно, начи-
ная с 40 тысяч лет назад до нашего времени. Место —
Южная Африка.
4. Негр; современная форма. Темнокожие люди
разного роста, с характерными широкими ноздрями,
отогнутыми наружу губами и выступающей вперед
нижней частью лица. Место — Африка к югу от
Сахары (и эмигранты по всему миру).
Древние и современные
обитатели Африки.
Нумерация рисунков
соответствует принятой в тексте.
1.
Омо-1; реконструкция
основана на неполном черепе.
2.
Череп из Флорисбада; на
реконструкции видны
архаичные черты, что позволяет
отнести этот череп к одной из
древних разновидностей Homo
sapiens.
3. Современный бушмен.
4. Современный негр.
|
|
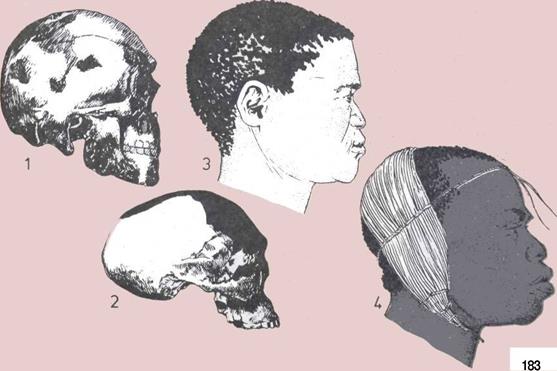
Места обитания человека
современного типа в Азии.
На этой карте показано 15
стоянок человека вполне
современного типа, возраст
которых составляет 50—100
тысяч лет.
1. Кафзех.
2. Ксар Акил.
3. Махадаха.
4. Аттирампаккам.
5. Батадомбалена.
6. Новоселове
7. Мальта.
8. Ордос (находки орудий).
9. Чжоукоудянь.
10. Цзиян.
11. Люцзян.
12. Минатогаа.
13. Пещера Табон.
14. Пещера Ниах.
15. Вадьяк.
Ископаемые черепа 40-тысячелетней давности,
которые относятся к вполне современному типу,
встречены в самых различных районах Азии — от
Израиля до Явы. У всех них есть подбородочный
выступ или другие явно «современные» признаки.
Но на западе и юго-востоке Азии попадаются также
формы с архаичными особенностями, характерными
для неандертальцев (см. с. 146—147). Некоторые
дальневосточные формы приводят в замешательство
исследователей, будучи похожими на еще более ран-
него человека Homo erectus. Специалисты объясняют
это по-разному.
Судя по некоторым черепам из Китая, можно ду-
мать, что 20 тысяч лет назад там жили предки со-
временных азиатских монголоидов — невысокие ко-
ренастые люди с белой или коричневой кожей, пря-
мыми черными волосами, слабо развитым волосяным
покровом на теле, с особой складкой кожи на верхнем
|
|
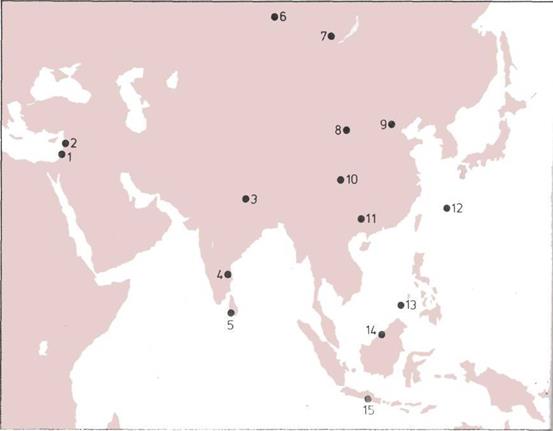
Древние и современные
обитатели Азии.
Нумерация рисунков
соответствует принятой в тексте.
1. Череп из Кафзеха.
2. Человек из пещеры Ниах.
3. Человек из Вадьяка.
4. Человек из Люцзяна.
5.
![]()
![]() Азиатский монголоид.
Азиатский монголоид.
|
Человек из Дель-Мар. Этот череп «древнейшего |
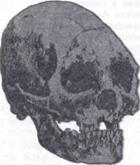
Ранние орудия обитателей
Америки.
1.
Наконечник культуры кловис
(10 тысяч лет до н. э.) с выемкой
для прикрепления к древку.
2.
Костяной скребок для
очистки кожи, зазубренный с
одного конца (25 тысяч лет
до н. э.).
|
|

Впервые люди появились в Северной Америке,
вероятно, между 70 и 12 тысячами лет назад. В пе-
риоды! наибольших похолоданий в это время море
отступало и образовывалась широкая сухопутная
перемычка Берингия, которая сейчас затоплена Бе-
ринговым проливом. Охотники-собиратели, вероятно,
прошли по ней не замочив ног из Сибири на Аляску.
Продвинувшись на юг по одному из свободных ото
льда путей, они распространились по Великим рав-
нинам и проникли в Южную Америку, достигнув
Патагонии.
По мнению антропологов, у этих так называемых
палеоиндейцев бвии прямые чернвю волосы, мед-
ного оттенка кожа, темные глаза, широкие скулы
и лопатовидные передние зубы-резцы. Перечислен-
ные монголоидные черты сохранились у американ-
ских индейцев и сейчас.
Но все еще вызывает споры вопрос о том, когда
именно пришли в Америку их предки. Сторонники
более поздней датировки указывали на то, что наибо-
лее ранние ископаемые остатки имеют возраст всего
лишь 12 тысяч лет. Примерно в то время на террито-
рии от Аляски до Мексики появились специфиче-
ские каменные наконечники для копий с искусно
сделаннвши выемками у основания. Распростране-
ние этих наконечников так называемой культуры
кловис, возможно, было связано с резким увеличе-
нием численности населения в то время, когда палео-
индейцы достигли богатых зверем и дичью земель
к югу от северных ледниковых покровов.
Сторонники более ранней даты утверждают, что
человек пришел в Америку более 40 тысяч лет назад.
Но после того, как была уточнена датировка черепа
человека из Дель-Мар в Калифорнии, которому,
как предполагали раньше, бвио 48 тысяч лет, его
возраст бвт уменьшен до 11 тысяч лет.
Таким образом, мы приходим к средней дате —
человек появился в Америке около 30 тысяч лет на-
зад.. Эта дата подкрепляется находками из бассейна
Старого Ворона на Юконе; там был найден, в част-
ности, костяной скребок для очистки шкур, кото-
рому, очевидно, 27 тысяч лет. К другим достовер-
ным находкам, которым 20 тысяч лет или более, от-
носятся очаги и кости 22-тысячелетнего возраста
из Мексики.
Палеоантропологи все еще спорят о том, когда че-
ловек впервые вступил на землю Северной и Южной
Америки и происходила ли эта миграция в виде не-
скольких следовавших одна за другой волн (которые
предшествовали наиболее поздним пришельцам —
эскимосам). Не подлежит сомнению лишь тот факт,
что 11 тысяч лет назад эти два последних незасе-
ленных, но пригодных для обитания человека кон-
тинента были уже населены людьми от одного конца
до другого.
![]()
![]()
![]() W"
W"
О Стадия 1
• Стадия 2
С Стадия 3
![]()
![]()
![]() Ш Стадия 4
Ш Стадия 4
Когда исчезли эти животные.
Все эти крупные млекопитающие
Северной Америки вымерли в
сравнительно недавние времена.
Некоторые ученые относят
их гибель к более поздним
временам, чем те, которые
указаны на приведенной выше
карте. На схеме цифрами
обозначено, сколько
тысячелетий назад исчезла,
согласно подсчетам одного
из специалистов, каждая
из этих групп животных.
а — Наземные1
ленивцы.
б — Глиптодонты.
в — Мастодонты.
г — Мамонты.
д — Лошади.
![]() е — Доисторические верблюды.
е — Доисторические верблюды.
ж — Доисторические
вилорогие антилопы.
з — Доисторические мускусные
быки (овцебыки).
и — Длиннорогие бизоны.
В период от 12 до 5 тысяч лет назад словно огром-
ной волной вымирания было сметено много крупных
животных Северной и Южной Америки. Ее жертвами
стали семь доисторических разновидностей слонов
(четыре вида мамонтов и три вида мастодонтов),
три вида верблюдов, длиннорогий бизон, лошадь,
некоторые вилорогие антилопы и мускусные быки,
гигантские наземные ленивцы (мегатерии), гигант-
ские броненосцы (глиптодонты) и саблезубые кошки.
Пик вымираний приходится на период от 12 до
10 тысяч лет назад — на то время, когда палеоин-
дейцы завладели обоими континентами. Поэтому
некоторые ученые считают, что эти виды животных
были истреблены человеком. Согласно этой теории
массового уничтожения, избиение животных было
совершено копьями с каменными наконечниками,
которые метались с помощью специальных приспо-
соблений. Один специалист утверждает, что 100
человек, продвигаясь из Канады на юг со скоростью
10 миль (16 км) в год, могли размножиться за 300
лет до 300 тысяч человек, достигнуть Мексиканского
залива и уничтожить по дороге за это время 100 мил-
лионов крупных млекопитающих.
Но это вымирание животных совпало по времени
и с радикальными климатическими изменениями,
связанными с таянием северных ледяных щитов.
Колебания температуры и количества дождевых
осадков резко влияли на растительность, лишая
пищи некоторых травоядных; таким образом, изме-
нения климата явились косвенной причиной голода
хищников, которые питались травоядными живот-
ными.
Согласно третьей теории, в вымирании виноваты
как человек, так и климат. По этой теории, изменения
| 20 тыс. лет |
15 |
10 |
5 1 |
| А 4*б |
|||
| Я1 |
2 |
||
| т |
|||
| ТП |
|||
| \т |
ТКе |
||
| Рж |
|||
| и |
климата, повлиявшие на растительность, привели к
сокращению площади питания определенных видов
животных. Сосредоточенные в этих районах и ослаб-
ленные нехваткой пищи целые стада становились
жертвой охотников.
![]() По мере исчезновения мамонтов и увеличения
По мере исчезновения мамонтов и увеличения
численности людей следовавшие друг за другом куль-
туры охотников на мамонтов — кловис и фолсом —
около 10 тысяч лет назад сменились культурой плано,
в которой сочетались охота на бизонов и собирание
дикорастущих растений. Охотничья стадия культуры
североамериканских индейцев сменилась культурой
с экономикой смешанного типа, относящейся к так
называемой архаической стадии, которая заверши-
лась около 2300 лет назад.
Наконечники копий и добыча
охотников.
На рисунках показаны оружие
и животные, на которых
охотились представители шести
доисторических
Наконечники копий.
1.
Наконечник типа фолсом
с выемкой.
2.
Наконечник типа иден —
узкий.
3.
![]() Наконечник типа ева —
Наконечник типа ева —
с зазубринами.
| А 1 i |
А — Культура плано, около 8 В— —5 Кут^^,л фолсо"и ,э . |
| 5 1 |
9—7 тысяч лет до н. э. |
| В Ц |
метательных орудий, 36(?)-- |
| & г * it |
д — Доисторический верблюд. Г |
| д 1 |
г% /Л В 1Н |
| Е €$> |
ЯШ 3 " ЧНК |
Четыре обитателя Австралии.
1.
Массивный череп, возраст
около 9,5 тысячи лет до н. э.
2.
Тонкостенный череп
(грациального типа), 28 тысяч
лет до н. э.
3.
Массивный череп,
удлиненный вследствие того,
что он был подвергнут сжатию
в младенчестве.
4.
Современный австралийский
абориген; австралоид с мощными
надглазничными валиками,
выступающими вперед
челюстями, крупными зубами,
хрупким телосложением, темной
кожей и волнистыми
волосами.
Следы очагов и ископаемые остатки, возраст ко-
торых установлен, свидетельствуют, что современный
человек жил в Австралии по крайней мере уже 40
тысяч лет назад. Некоторые специалисты счита-
ют, что люди пришли туда 120 тысяч лет назад, когда
на территории Нового Южного Уэльса резко увели-
чилось возгорание кустарников и широко распро-
странились огнестойкие растения современных видов.
Но в Австралии не найдено остатков человеческих
скелетов или орудий, относящихся к этому периоду.
Вероятнее всего, люди впервые появились здесь
в период от 55 до 45 тысяч лет назад, когда уровень
океана был на 160 футов (50 м) ниже, чем сейчас.
и многие острова составляли единое целое. Жителям
Юго-Восточной Азии достаточно было пересечь на
каноэ или плотах всего лишь 40 миль (65 км) моря,
чтобы попасть в Меганезию — материк, частью ко-
торого являлась Австралия.
Первые австралийцы отнюдь не были похожи друг
на друга. Около .30 тысяч лет назад на берегах озера
Мунго на юго-востоке материка жили люди совре-
менного типа, но хрупкого сложения, которые по
строению скелета несколько напоминали современ-
ных китайцев. Но 13 тысяч лет назад в районе Кау-
Суомп, расположенном неподалеку, жили люди более
|
|

плотного телосложения, с массивным черепом, вы-
ступающими вперед челюстями, с надглазничными
валиками и покатым лбом, т. е. они обладали теми
признаками, которые были присущи человеку из
Соло — архаичному Homo sapiens с острова Ява.
Поэтому одни ученые полагают, что в Австралии
имели место две волны переселения. Другие же спе-
циалисты утверждают, что разнообразные формы
доисторического человека представляли собой лишь
крайние варианты предков современных абориге-
нов, надглазничные валики которых были наследи-
ем глубокой древности.
![]() Перед тем как в Австралии два века назад выса-
Перед тем как в Австралии два века назад выса-
дились европейцы, все аборигены вели жизнь, харак-
терную для позднего палеолита. Небольшие полу-
кочевые группы почти голых людей бродили каж-
дая по своей собственной постоянной территории.
Мужчины, вооруженные копьями и бумерангами,
охотились на дичь. Женщины собирали растения и
насекомых. Мифы и ритуалы играли большую роль
в их жизни, что нашло отражение в танцах, живопи-
си, песнях и легендах, передаваемых изустно. Все
это сохранилось и в настоящее время, поэтому изу-
чение обычаев австралийских аборигенов позволяет
нам лучше понять образ жизни доисторических лю-
дей и в других частях света.
языковой группы расселились
по Тихому океану от Гавайских
островов до Новой Зеландии.
1.
Люди крупного
телосложения, 45 тысяч лет до
н. э.
2.
Люди более хрупкого
(грациального) телосложения,
45 тысяч лет до н. э.
3.
![]()
![]()
![]()
![]() Австронезийцы (главным
Австронезийцы (главным
образом монголоиды), 5000 лет
до н. э. — 1000 лет н. э.
Цвет кожи.
![]()
![]() Оттенки цвета кожи аборигенов
Оттенки цвета кожи аборигенов
связаны с количеством
солнечного излучения, за
исключением тех районов, где
в результате недавних миграций
естественный отбор длился
в течение слишком краткого
периода, чтобы он мог оказать
существенное влияние на цвет
кожи.
По мере того как наш подвид распространялся по
всему миру, разные группы людей оказывались в
различных климатических зонах. В ходе естествен-
ного отбора люди физически приспособились к раз-
личным природным условиям. Вследствие этого по-
явились люди с черной, белой и желтой кожей. Цвет
кожи отнюдь не означает принадлежности к той или
иной расе (т. е. к биологической группе людей, свя-
занных общим происхождением). И у негров, и у
меланезийцев черная кожа, по они столь же мало
связаны родством друг с другом, как и с людьми,
имеющими белую или желтую кожу. И разумеется,
все люди принадлежат к одному и тому же подвиду.
Рассмотрим вкратце влияние, которое оказывал
жаркий или холодный климат на кожу, развитие
телосложения людей черной, белой и желтой кожи
и на появление существенных различий между ними.
![]() Возьмем для начала цвет кожи. У людей с белым
Возьмем для начала цвет кожи. У людей с белым
цветом кожи от сильного тропического солнечного
излучения она начинает шелушиться и покрываться
волдырями, а интенсивное ультрафиолетовое излу-
чение может вызвать даже рак кожи. У чернокожих
людей кожа, окрашенная темным пигментом мела-
нином, защищена от этих вредных воздействий. Но
в расположенных за пределами тропиков странах,
где небо чаще покрыто облаками, более светлый
цвет кожи является преимуществом, так как он по-
зволяет поглощать достаточное количество слабого
ультрафиолетового излучения, необходимого для
выработки организмом витамина D, который спо-
собствует здоровому росту костей. В прохладных,
туманных странах чернокожие дети часто болеют
рахитом.
|
|
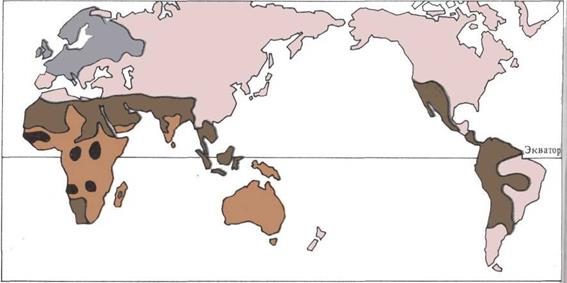
Солнечное излучение и рак
кожи.
На диаграмме показана частота
заболевания раком кожи
европейского и темнокожего
населения Южной Африки.
1. Разъедающая эпителиома.
2. Рак губы.
а — Мужчины европейцы.
б — Женщины европеянки.
в — Темнокожие мужчины.
г — Темнокожие женщины.
Имеется также ряд важных факторов, влияющих
на телосложение — на форму и размеры тела. У вы-
сокого худощавого негра-нилота с длинными ко-
нечностями очень велика поверхность тела сравни-
тельно с его объемом, а такое соотношение идеально
для излучения избыточного тепла, чтобы не страдать
от жары в тропиках. В тех же условиях эскимос
(монголоид) с его невысоким коренастым телом,
имеющим небольшую поверхность, страдал бы от пе-
регрева. Не такое телосложение с изолирующим
подкожным слоем жира позволяет сохранять тепло,
важное для жизнедеятельности в условиях аркти-
ческих зим, когда человек худощавого телосложе-
ния страдал бы от переохлаждения, гибельного для
его организма.
Конечно, мы несколько упрощаем общую картину.
Так, отнюдь не все монголоиды имеют то же тело-
сложение, что и эскимосы. Но биологические при-
способления, подобные вышеописанным, без сом-
нения, помогли человеку эпохи палеолита населить
все пригодные для обитания континенты.
Ключ к карте распределения
цвета кожи
(слева).
ЦЦ Очень бледный
□ Довольно бледный
Промежуточный
Довольно темный
ЦНЦ Очень темный
Ключ к карте распределения
формы волос
(справа).
Курчавые и густые
| I Прямые
Волнистые и прямые
13 Зак. 5116.
Форма волос.
![]()
![]()
![]()
![]() Форма волос связана с
Форма волос связана с
интенсивностью солнечного
излучения. Курчавые волосы
некоторых тропических пародов
защищают голову от теплового
излучения Солнца.
![]() Со времени окончания оле-
Со времени окончания оле-
денений, за последние
10 ты-
сяч лет человек мало изме-
нился физически. Но куль-
турная эволюция преобра-
зила человечество. В этой
главе пойдет речь об основ-
ных доисторических нововве-
дениях
— от добывания пи-
щи и развития металлургии
до возникновения городов и
изобретения письма. Мы уви-
дим, насколько культурные
новшества изменили наш
внешний облик и способ-
ствовали огромному росту
численности нашего вида, и
закончим эту главу смелым
взглядом в будущее.
Человек, пашущий на упряжке
волов, изображен на этом
наскальном рисунке бронзового
века из Швеции. Около
5 тысяч лет назад на большей
части Европы охота сменилась
земледелием. (Иллюстрация
из книги Джеффри Бибби
«Открытия, сделанные
лопатой».)
|
|

![]() 195
195
Расширение площади суши.
На этих картах показаны
изменения территории
Скандинавии начиная с 8300 г.
до н. э.
1.
8300 г. до н.э.; большая часть
Скандинавии покрыта льдом.
2.
7000 г. до н.э.; лед растаял,
но пониженные участки суши
покрыты водой.
3.
![]()
![]()
![]() 5000 г. до н. э.;
5000 г. до н. э.;
произошло значительное
поднятие поверхности суши.
Голоцен, или «новая» геологическая эпоха, кото-
рая началась около 10 тысяч лет назад, ознаменова-
лась огромным культурным прогрессом, которому
суждено было привести в конечном результате к
господству человека на Земле. Эти культурные заво-
евания совпали по времени с потеплением климата
после последней длительной волны похолодания
ледникового периода, которой завершился плей-
стоцен. (Возможно, что в действительности голоцен
является всего лишь кратким промежутком потепле-
ния перед наступлением очередного периода похо-
лодания.)
Значительные изменения на земном шаре про-
изошли вскоре после начала таяния северных лед-
никовых покровов 15 тысяч лет назад. Стекающая
в океан талая вода постепенно подняла его уровень
на 430 футов (130 м). Море затопило сухопутные
перемычки, которые соединяли Азию и Северную
Америку, Индонезию и Малайю, Британские острова
и Европу. Одна двадцатая часть всей поверхности
земной суши была покрыта морем, по в то же время
но мере сокращения ледниковых покровов обнажа-
лась та суша, которую раньше покрывал лед. В не-
которых районах Канады и Скандинавия старая
береговая линия теперь поднята на 980 футов (300 м)
над современным уровнем моря.
Согласно исследованиям ученых, за эпоху голо-
цена климат Европы прошел через несколько стадий
изменений, каждая из которых оказала влияние как
на сушу, так и на фауну, ее населявшую. В пред-
бореальное и бореальное время (10 000-7750 лет
назад) ледниковые покровы и горные ледники сокра-
тились в размерах и количество дождевых осадков
в средних широтах и в тропиках значительно воз-
росло. С повышением уровня воды в озерах и реках
Сахара и некоторые другие засушливые районы
мира покрылись влажными лугами и превратились
в пастбища. Дикие евразийские быки, козы и овцы
мигрировали на север, сменив там лосей и северных
оленей. В Юго-Западной Азии появились первые
земледельческие поселения.
В северных странах климат атлантического вре-
мени (7750 -5100 лет назад) был теплее, чем в на-
стоящее время. Растительность распространилась
по всему миру, за исключением высочайших горных
вершин; значительная часть северной тундры была
вытеснена лесом. Охотники индейцы населили когда-
то покрытые ледниками части Северной Америки,
и земледелие распространилось по всей Евразии.
В суббореальное время (5100—2200 лет назад)
зимы в центральных областях континентов на сред-
них широтах стали прохладнее и суше. Увеличилась
численность степных и луговых животных. В Цент-
ральной Америке начало развиваться земледелие, и
на некоторых континентах возникла городская
жизнь. В некоторых местах начала появляться пись-
менная хроника событий -- человечество порывало
со своим доисторическим прошлым.
Около 2200 лет назад началось то субатлантиче-
ское время с его климатическими колебаниями, в
которое мы живем и сейчас.
Затопленные участки суши.
Площадь суши на юго-востоке
Северной Америки сократилась,
когда уровень моря повысился
в результате таяния льда.
к 1
X 6
|
а — Современная береговая б — Береговая линия во время |
Культурные достижения в эпоху
голоцена.
Над шкалой времени и графиком.
отражающим колебания
температуры за последние
10 тысяч лет со времени
значительного потепления
в конце плейстоцена,
изображены символы
культурных достижений
человека.
а — Появление земледелия.
б — Появление городов.
в — Керамика, изготовленная
па гончарном круге (а также
плавка металлов и ткачество).
г — Колесо (а также изобретение
письма и постройка парусных
кораблей).
д — Государственность.
е — Классические цивилизации
ж — Век открытий.
з — Промышленная революция.
|
|

10 тысяч лет назад началось преобразование жиз-
ни человечества, связанное с открытием земледе-
лия. Определенный участок плодородной земли
может прокормить значительно больше земледель-
цев, чем охотников-собирателей; поэтому там, где
распространялось земледелие, население возрастало.
Кроме того, уход за посевами способствовал стрем-
лению к оседлому .образу жизни в поселениях, где
имелись мастера, которые занимались изготовлением
и обменом таких предметов, как полированные ка-
менные орудия и гончарные изделия.
Эти культуры эпохи неолита (нового каменного
века), связанные с земледелием и/или скотовод-
ством, сменили культуры палеолита и мезолита и
предшествовали культурам бронзового и железного
веков.
Различные неолитические культуры возникали,
по-видимому, независимо друг от друга в различных
местах и в разное время. Земледелие впервые широко
распространилось около 10 тысяч лет назад внутри
и поблизости от так называемого «Плодородного
полумесяца», который охватывает Египет, Юго-
Западную Азию и побережье Персидского залива.
Здесь первые земледельцы занимались возделыва-
нием пшеницы, ячменя, чечевицы и гороха. Пример-
но 7 тысяч лет назад китайцы начали выращивать
такие культуры, как просо, рис, соя, таро и ямс.
5 тысяч лет назад. Мезоамерика, или Центральная
Америка (Южная Мексика, Гватемала и Гондурас),
превратилась в третий великий центр земледелия,
где занимались возделыванием кукурузы, фасоли,
тыквы и хлопка.
Ученые спорят о том, когда и почему люди камен-
ного века перешли к оседлому образу жизни и к зем-
леделию. Многие собиратели-охотники уже добывали
достаточно пиши для своего пропитания и имели
много досуга. Ранние земледельцы работали зна-
чительно больше, получая менее разнообразные
продукты питания и рискуя оказаться голодными в
случае неурожая.
Однако в некоторых районах рост населения обго-
нял увеличение охотничьей добычи, и это, возможно,
вынуждало охотников искать дополнительные источ-
ники питания в виде растений. Другим мощным фак-
тором, вероятно, было распространение трав с круп-
ными съедобными зернами.
Сбор урожая дикорастущих
культур.
В урожайный сезон один
человек мог собрать за час
6 ![]()
![]()
![]() фунтов (почти 3 кг) зерен
фунтов (почти 3 кг) зерен
дикой полбы, а семья могла
заготовить за три недели запас
пиши на целый год.
|
|

От колоса до муки.
На верхних рисунках показано,
как древние земледельцы
обрабатывали спелую пшеницу.
1.
Жатва; уборка колосьев
пшеницы серпом. Срезать было
легко только твердые колосья,
которые оставались при этом
целыми.
2.
Молотьба; колосья молотили,
пока они не распадались на
отдельные зерна.
3.
Веяние; зерно
подбрасывали вверх, чтобы
ветер унес ненужные частицы
соломы.
4.
![]()
![]()
![]() Измельчение зерен пшеницы
Измельчение зерен пшеницы
и освобождение их от мякинной
оболочки.
Из зерен можно было сварить
кашу или, перемолов их в муку,
испечь хлеб.
13 тысяч лет назад собирание зерен дикорасту-
щих злаков стало основным источником питания,
по крайней мере одной группы населения в Израиле.
Люди натуфийской культуры пользовались серпами
с кремневыми лезвиями, чтобы срезать спелые ко-
лосья диких родственников современной пшеницы
и ячменя. Со временем такие группы людей, систе-
матически занимавшиеся собирательством, начали
сеять семена, отбирая те сорта пшеницы и ячменя,
которые было легче всего возделывать. Селекция
такого типа должна была превратить дикорасту-
щие растения с маленькими семенами, плодами,
корнями и листьями в те крупные домашние сорта,
которые служат нам пищей в настоящее время.
Места обитания диких предков
домашних животных.
На этих четырех каргах покачано
распространение диких
млекопитающих, бывших
предками одомашненных видов
животных. Все чти животные,
кроме зубров, и сейчас еще
обитают в основном в тех же
районах.
1. Ovis, овца. Только вили (а)
и (б) оказали значительное
влияние на формирование
современных пород.
а—Ovis orienlali.s,
малоазиатский муфлон.
б Ovis vignei, уриал (степной
баран).
в Ovis ammon, архар
(горный баран).
2. — Capra aegagrus, дикий
козел, назвшаемый также
безоароиыч козлом.
3. Sits scroja, кабан.
4. Bos primigenius. зубр,
вымерший дикий бык
I la карте показано его
распространение в эпоху
плейстоцена.
Точно так же, как собирательство пищи привело
к появлению земледелия, охота привела к одомаш-
ниванию животныи. И опять же основным центром
здесь быта Юго-Западная Азия. 13 тысяч лет назад
собаки, произошедшие от прирученных волков, воз-
можно, уже помогали охотникам Юго-Западной
Азии загонять стада и отдельные группы диких ко-
пытных млекопитающих в ущелья, чтобы их можно
быгло там забить. 8,5 тысячи .чет назад некоторые
охотники уже превратились в пастухов, разводив-
ших овен, коз и крупный рогатый скот, которые слу-
жили им в качестве живых источников мяса и одеж-
ды. Удобным быгло то, что каждая из -лих групп
животныи питалась растениями, несъедобными для
человека: козы обгрызали деревья и кустарники,
овцы ели травы, росшие на склонах холмов, а круп-
ный рогатый скот пасся на богатых лугах долин.
Тем временем свиньи, произошедшие от диких каба-
нов, стали полезными «уборщиками» мусора в де-
ревне.
Селекция всех этих пород привела к появлению
послушных животных, которые имели более короткие
рога или клыки, чем их дикие предки, и давали боль-
ше мяса, молока, шерсти. Об этом косвенно свиде-
тельствуют кости, которые находят на древних скот-
ных дворах.
|
|
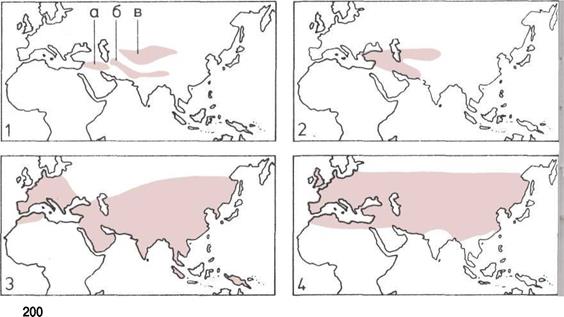
5 тысяч лет назад одомашненные быки, верблюды,
ослы, лошади, а в Южной Америке также и ламы
уже перевозили грузы, что в корне преобразовало
путешествия по суше.
Вот некоторые сведения о диких предках четырех
животных, которые сейчас распространены во всем
мире и мясо которых мы употребляем в пищу.
1. Ovis orientalis, дикая овцу, с более минными
рогами, шерстью и конечностями, чем у домашней
овцы. Время приручения около 11 тысяч лег на-
зад. Место возможно, Турция, Иран или Ирак
2. Capra aegagrus, дикая коза, с рогами, загибаю-
щимися сразу назад без тех спиральных завитков,
которые можно видеть у домашних коз. Время при-
ручения около 10 тысяч лет назад. Место воз-
можно, Иран.
3. Bos primigenius, тур, имел более крупные раз-
меры, большую длину, более длинные рога и был
агрессивнее, чем ранний одомашненный крупный
рогатый скот. Время приручения около 8,5 тысячи
лет назад. Место возможно, Турция.
4. Sus scroja, дикий кабан, с более длинным рылом
и более густой щетиной, чем у домашних свиней. Вре-
мя приручения около 9 тысяч лет назад. Место
возможно, Турция.
Дикие предки домашних
животных.
Ну мерация рисунков
соответствует принятой в тексте.
1. Ovis oripn tails.
2. С.ирги aegagrus.
3. Bos prlmigenius.
4.
![]() Sus
scrota
Sus
scrota
|
|
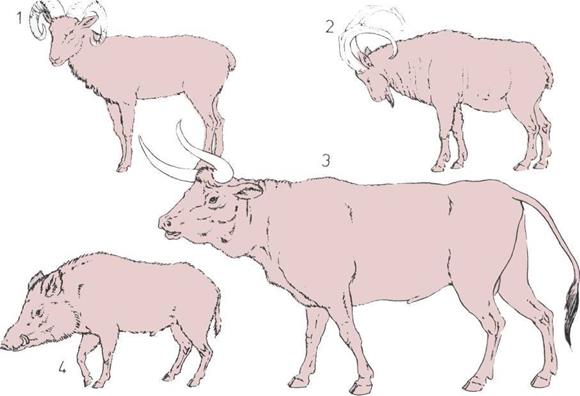
Торговые маршруты эпохи
неолита.
На карте показаны пути
торговли обсидианом
в Средиземноморье во времена
неолита. Существовали два
основных района торговли —
в западной и восточной частях
Средиземного моря, причем
большая часть экспорта шла
с одних островов на другие или
на европейский материк.
К основным центрам экспорта
относились:
1. Сардиния.
2. Понцианские острова.
3. Липарские острова.
4. Остров Пантеллерия.
5. Остров Милос.
6. Джали.
7. Чифтлик.
8. Ачигёль.
9. Озеро Ван.
Сейчас археологи уже отказались от старого
представления о том, что люди эпохи неолита имели
стандартный набор орудий с полированными камен-
ными топорами, гончарными изделиями и камнями
для помола зерна. Теперь мы знаем, что, например,
в Греции земледелие появилось раньше, чем гон-
чарное дело, а в Испании и Скандинавии, наоборот,
земледелие было освоено позже. Развитие земле-
делия сопровождалось распространением орудий
для расчистки полей, подготовки почвы к посевам,
сбора урожая, складирования запасов зерна, выпеч-
ки хлеба, ткачества и строительства помещений для
живых людей, покойников и изображений богов.
Жители поселений эпохи неолита изготовляли
большинство своих орудий сами, но часть предме-
тов выменивали у жителей других местностей, на-
пример, такие товары, как соль, гематит (для румян
и покраски гончарных изделий) и обсидиан — стек-
ловатую горную породу, применявшуюся для из-
готовления орудий и украшений. Некоторые рудники
|
|
снабжали районы, удаленные от них на несколько
сотен миль.
Приведем шесть примеров орудий тех типов, кото-
рые были характерны для ранних земледельческих
культур.
1. Топор с лезвием из полированного камня; ис-
пользовался для расчистки зарослей леса. Этот об-
разец происходит из Дании.
2. Деревянная мотыга; использовалась для рас-
чистки земли под посевы. Этот образец происходит
из Египта.
3. Серп с лезвием из кремня; использовался для
жатвы пшеницы или ячменя. Этот образец происхо-
дит из Египта.
4. Глиняный горшок; использовался для хране-
ния и варки пищи или воды. На этом образце из
Кении насечен узор в виде плетеной корзинки. Такое
обожженное прочное гончарное изделие, возмож-
но, получилось при случайном сгорании корзинки,
обмазанной изнутри глиной, более 9 тысяч лет на-
зад.
5. Жернов и мельничный камень; применялись
для помола зерна. Такие каменные изделия были
широко распространены.
6. ![]() Ткацкий станок; изображен на египетском гон-
Ткацкий станок; изображен на египетском гон-
чарном блюде. Ткацкое и прядильное искусство воз-
никли по крайней мере 7 тысяч лет назад.
|
|
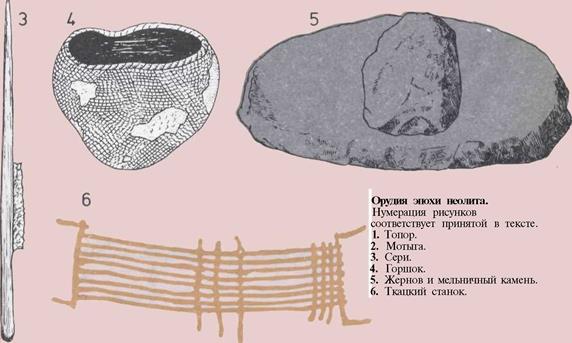
Поселение эпохи неолита.
Раскопки столбов и углублений
из-под них позволили
художникам создать
реконструкцию этой деревни
времен неолита, расположенной
на берегу озера в Юго-Западной
части Германии.
А — На плане показаны
группировка и расположение
хижин.
![]() Б — Так выглядели хижины
Б — Так выглядели хижины
со стороны озера. Деревянные
каркасы придавали им
прочность.
Неолитические культуры Старого Света никогда
не достигали уровня замечательного искусства на-
скальных изображений позднего палеолита, хотя в
неолите и изготовлялись статуэтки и искусно укра-
шенная многочисленными узорами керамика. Воз-
можно, наибольших успехов создатели неолитиче-
ских культур добились в строительстве, более раз-
нообразном и замечательном, чем когда-либо
прежде.
Мастера строили деревни и поселения из материа-
лов, имевшихся в данной местности. Так, в Иерихо-
не в Палестине были дома из глиняных кирпичей
со штукатурными стенами и полами, защищенные
наружными каменными стенами. Дома с глиняными
стенами были распространены в Юго-Восточной
Европе и Юго-Западной Азии с их сухим климатом.
Но в дождливых районах Западной Европы много
строили из леса. Здесь дома имели деревянные стол-
бы и коньковые брусы, поддерживающие наклон-
ные крыши; вода стекала с карнизов, выступающих
над сплетенными из прутьев стенами, которые были
обмазаны для защиты от воды глиной или навозом.
В Швейцарии озерные жилища стояли на деревян-
ных сваях, которые предохраняли их от риска за-
топления. Дома в Скара-Брэй — неолитическом по-
селении на безлесных, продуваемых ветрами Орк-
нейских островах, имели каменные стены; крыши
были из плавника, шкур и торфа.
Но наиболее внушительное впечатление произво-
дили неолитические сооружения, выполнявшие ре-
лигиозные функции. Более 8 тысяч лет назад десятки
храмов стояли в раннем городском поселении Чатал-
Хююк, которое сейчас находится на территории Тур-
ции. 6 тысяч лет назад вырубленные из камня гроб-
$ ш Ля ,
Л F i W4
' а,
ницы появились в Италии и на Мальте, где из огром-
ных каменных глыб, называемых мегалитами, соору-
жались неолитические заупокойные храмы. 5 тысяч
лет назад строители мегалитов на Британских остро-
вах и в Бретани независимо друг от друга сооружа-
ли дольмены, состоящие из отдельных камер; это
были гробницы с галереями или крытыми ходами.
По мере перехода от неолита к энеолиту, т. с. мед-
ному веку, который предшествовал бронзовому, мно-
жество людей посвящали миллионы человеко-часов
возведению рядов и кругов из вертикально стоящих
камней в таких местах, как Карнак в Бретани и
Стоунхендж в Южной Англии. В планировке этих
каменных сооружений проявлялось некоторое зна-
комство с астрономией.
К 2700 г. до н. э. общие захоронения в виде длинных
могильных курганов уступили место единичным
гробницам в круглых могильных холмах. Золотые и
другие ценные предметы в могильниках говорят о
высоком статусе некоторых из похороненных там
людей. Возможно, это были крупные вожди, настоль-
ко могущественные, что они могли сооружать для
себя каменные монументы. Происходило социальное
расслоение общества.
Время возведения
мегалитических сооружений.
Полосками показаны периоды
создания различных типов
мегалитических построек
в Европе.
1. Храмовые комплексы.
2. Гробницы с крьными ходами.
3. Гробницы типа галерей.
4. Круги из камней (кромлехи).
5.
![]() Ряды камней (менгиры).
Ряды камней (менгиры).
| 5000 лет до н.э: ГййЗ; I |
4000 |
3000 |
2000 |
| 1 i>" !„# 2 |
|||
| v_ |
____ 3 |
||
| L \ г |
|||
| I / |
5 |
|
|
Центры добычи золота.
Металлурги доколумбовой
Америки придавали особое
значение изготовлению золотых
украшений. На карте
показаны некоторые центры
обработки золота и названы
народы, которые этим
занимались.
1. Ацтеки.
2. Микстеки.
3. Майя.
4. Верагуас.
5. Тайрона.
6. Муиска.
7. Чиму.
8. Мочика.
9. Инки.
Древние центры обработки
металлов.
На этой карте показан ряд
древних стоянок, где были
найдены следы обработки
металлов. Большинство из них
находилось в районах горных
хребтов, богатых рудами
металлов. Несмотря на очень
древний возраст некоторых
центров ковки меди в Юго-
Западной Азии, плавка
металлов на юго-востоке Европы
была, вероятно, открыта
независимо от них.
1.
Тишапольгар, 4500 лет
до н. э.
2. Гюльменица, 4400 лет до н. э.
3. Чатал-Хююк, 6000 лет до н. э.
4. Тимна, 4000 лет до н. э.
5.
Кайёню-Тепези, 7200 лет до
н-. э.
6. Шанидар, 9500 лет до н. э.
7. Тепе-Сиалк, 5000 лет до н. а.
8.
Таль-и-Иблис, 4000 лет
до н. э.
На протяжении более 2 миллионов лет самые твер-
дые и острые орудия и оружие изготовлялись из об-
битого камня. Но за последние несколько тысяч лет
металл в корне преобразовал технологию изготовле-
ния орудий. Подобно земледелию, центры металлур-
гии, очевидно, появились независимо друг от друга
в разное время в разных частях Европы, Азии и Аме-
рики.
Около 9 тысяч лет назад люди, жившие на юго-
востоке Турции, начали ковать вхолодную из чистой
меди булавки, бусы и сверла. Медь более прочная
и менее хрупкая, чем камень, и ей можно придавать
ту или иную форму и перезатачивать сделанные из
нее орудия. Но чистая медь встречается значительно
реже, чем ее руда — химические соединения этого
металла с различными примесями. Настоящая ме-
таллургия появилась только около 6600 лет назад,
когда кузнецы на юго-востоке Европы и юго-западе
Азии начали извлекать медь из руды при помощи
плавки.
Появлению плавильной печи, вероятно, предшест-
вовало строительство печей для обжига гончарных
изделий при нагреве до 1470° по Фаренгейту (800°С);
это достаточно высокая температура для плавки ме-
ди. Кузнецы научились разливать медь в каменные
формы. 5 тысяч лет назад они смешивали одну часть
олова и девять частей меди, изготовляя бронзовый
сплав. Этот сплав легче поддается отливке, чем медь,
и пригоден для изготовления более твердых орудий
и оружия.
![]() Примерно к 1500 г. до н. э. хетты в Турции начали
Примерно к 1500 г. до н. э. хетты в Турции начали
производить железо. В природе железо встречается
в значительно больших количествах, чем медь, но
труднее поддается плавке. Температура в хеттских
плавильных печах не достигала 2880° по Фаренгейту
(1535°С), т. е. точки плавления железа. Поэтому в
результате плавки получалась крица — прочная мас-
са, состоящая из железа и шлака. Кузнецы доводили
эту массу до красного каления и ковали ее, пока не
отделялся шлак. Чтобы сделать металл более проч-
ным, к нему примешивали древесный уголь, а изго-
товленные из раскаленного железа инструменты и
оружие закаляли в воде.
Ковка оставалась единственным способом обра-
ботки железа до тех пор, пока около 2500 лет назад
в китайских плавильных печах с высокой температу-
рой не стали изготовлять предметы из чугуна.
К тому времени на большей части Европы, Азии
и Северной Африки культура бронзового века сме-
нилась культурой железного века. Во всех этих трех
частях света массовое производство орудий из же-
леза способствовало развитию земледелия и тор-
говли, а также войнам и уничтожению лесов. В дру-
гих местах производство металлов имело меньшее В
значение. Африка южнее Сахары переходила от кам-
ня непосредственно к железу, а доколумбовая Аме-
рика так и не поднялась выше уровня бронзового
века.
![]()
![]()
![]()
![]()
![]()
Бронзовое литье (вверху).
На сценках из жизни Древнего
Египта можно видеть
следующие технологические
процессы.
Колыбели цивилизации.
На карте показаны места
возникновения городской жизни
на четырех великих речных
равнинах Старого Света.
1. Долина Нила.
2. Месопотамия.
3. Долина Инда.
4.
![]() Долина Желтой реки
Долина Желтой реки
(Хуанхэ).
5 тысяч лет назад производство продуктов пита-
ния и металлургия способствовали превращению
некоторых поселений в первые в мире крупные горо-
да. Цивилизация — наш образ жизни, связанный
с городами, впервые развилась у шумеров бронзово-
го века в Месопотамии — стране, расположенной
в междуречье Тигра и Евфрата на территории со-
временного Ирака. Затем первые города появились в
долине Нила в Египте, в долине Инда в Пакистане,
по берегам Жёлтой реки (Хуанхэ) в Китае, а также
в наиболее благодатных районах Перу и Мексики.
Во всех этих местах ирригация плодородных зе-
мель помогала крестьянам производить достаточное
количество пищи для пропитания больших концент-
раций городского населения, которое состояло из
специалистов, полностью посвятивших себя ремес-
лам, не связанным непосредственно с получением
продуктов питания. Горшечники, плотники, работ-
ники по металлу, ювелиры, писцы, купцы предлагали
свои услуги и меняли изготовленные ими товары на
пищевые продукты, производимые местными кре-
стьянами, и на такое сырье, как камень, дерево,
асфальтовый битум и металлы, которые доставля-
лись с гораздо большего расстояния.
По мере этих перемен человеческие сообщества
превратились из племенных групп в родовые кланы,
а потом в государства, расслоенные на обществен-
ные классы. Большая часть богатства и власти были
сосредоточены в руках царей или священников, пра-
вивших с помощью чиновников, которые, очевидно,
|
|
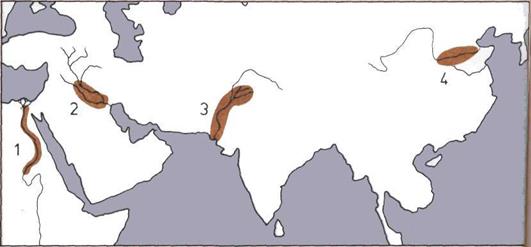
занимались организацией орошения, сбора налогов
и запасов продуктов питания для общих нужд.
Эта общественная структура нашла свое отраже-
ние в новых внушительных типах построек. Над ран-
ними городами Месопотамии возвышались зиккура-
ты — огромные ступенчатые кирпичные пирамиды,
увенчанные храмами. Храмовые чиновники, обладав-
' шие знанием письма, математики и астрономии,
по-видимому, пользовались календарями, чтобы оп-
ределять для крестьян время посева; замеряли поля,
а также собирали и хранили зерно, поступавшее
в виде налогов. К 3000 г. до н. э. строительство круп-
ных дворцов знаменует переход власти от жрецов к
царькам, которые правили в небольших городах-
государствах. Высокие стены защищали богатства
их городов от набегов кочевников и от нападения
соперничающих городов-государств.
![]()
![]()
![]()
Со временем в результате войн между соперни-
чающими городами-государствами население неко-
торых из них слилось в первые небольшие народы.
К 2279 г. до н. э. честолюбивый аккадский правитель
Саргон покорил страны от Турции до Персидского
залива, создав государство, которое можно назвать
первой империей в мире.
|
|
|
|
5 тысяч лет назад перевозки по,воде также под-
верглись коренным изменениям. Парусники перево-
зили теперь тяжелые грузы вверх по рекам. Поставив
себе на службу ветер и животных, люди смогли ис-
пользовать значительно большее количество энергии,
чем это позволяла мускульная сила человека.
Тем временем изобретение письма революциони-
зировало сферу человеческого общения. 5 тысяч лет
назад шумеры начали использовать тростниковые
перья, чтобы на мягких глиняных табличках рисовать
значки-картинки, называемые пиктограммами; при
засыхании и затвердевании эти таблички станови-
лись долговечными. Позже писцы заменили 2000 пик-
тограмм значительно меньшим количеством фоно-
грамм — словесно-звуковых символов, основанных
на пиктограммах. В результате упрощения они прев-
ратились в рукописный шрифт, воспроизводивший
разговорный язык. Писцы использовали свою шумер-
скую клинопись (письмо в виде клинышков) для за-
писи сводов законов, календарей, налогов и различ-
ных значительных исторических событий.
![]()
Изобретение письменности означало, что новые от-
крытия в науке, математике и астрономии могли быть
точно записаны и сохранены для потомства. Таким
образом, письменность способствовала развитию
культуры. А там, где началась запись исторических
событий, кончился доисторический период.
Клинописное письмо.
1.
Похожие на клинышки
знаки наносили тростниковой
палочкой на мягкую глиняную
табличку.
2.
В горизонтальных рядах
(слева направо) показано, как
два клинописных знака
превратились из пиктограммы
в фонограмму.
А — Рыба.
Б — Колос.
| А :=> $ |
||
| Б | т*— |
ш |
у |
![]()
Преграды в виде морей и пустынь изолировали
зарождающиеся цивилизации Евразии от Австралии,
Африки южнее Сахары, от Северной и Южной Аме-
рики. Но в этих странах, особенно в Америке, также
происходило развитие культуры. На следующих че-
тырех страницах мы дадим краткий обзор тенденций
развития доколумбовой Северной и Южной Аме-
рики.
К северу от Мексики к 5000 г. до н. э. индейцы все
еще занимались охотой на крупных животных на Ве-
ликих равнинах. На северо-западе значительную
роль играла также ловля рыбы в море и реках. В вос-
точных лесах основной упор делался на охоту, рыб-
ную ловлю и собирательство. В пустынях юго-запада
традиционно занимались сбором семян и ловлей
мелкой дичи. Эти виды занятий сохранились, но к
первым векам новой эры сельское хозяйство, гон-
чарное дело, изготовление тканей из растительных
волокон, погребальные курганы и центры религиоз-
ных церемоний распространились к северу от Мек-
сики, повлияв на культуры анасази и гогокам на
юго-западе и на культуры адена и хоупвелл в бас-
сейне Огайо. После 700 г. н. э. культура хоупвелл
уступила место культуре Миссисипи, а в зоне восточ-
нвк лесов появились крупные сообщества, занятые
возделыванием кукурузы.
Места обитания жителей
Северной Америки.
На этой карте показаны
культурные районы, на которые
разделилась Северная Америка
к 1500 г. н. э.
1.
Арктический; эскимосы-
охотники.
2. Субарктический; охотники.
3.
Северо-западное побережье;
рыбаки и охотники.
4.
Внутреннее плато; рыбаки
и охотники-собиратели.
5.
Великие равнины; охотники
на бизонов и земледельцы.
6.
Район восточных лесов;
охотники, рыбаки, собиратели
и земледельцы.
7.
Долины Огайо и Миссисипи;
земледельцы.
8.
Калифорния; охотники,
рыбаки и собиратели.
9.
Большой Бассейн;
собиратели, обитавшие
в пустыне.
10.
Юго-запад; земледельцы
и собиратели.
11.
Пустыни юго-запада;
собиратели.
12.
Центральная Америка
(Мезоамерика); земледельцы,
а также жители поселков
![]() и городов.
и городов.
Шесть местных культурных традиций представле-
ны следующими декоративными предметами.
1. Кувшин; предназначен для хранения пищевых
продуктов; принадлежит культуре гогокам в Ари-
зоне (около 300 г. до н. э. — 1400 г. н. э.), которая
известна своими оросительными каналами.
2. Ткань; относится к культуре хоупвелл (около
200 г. до н. э. — 700 г. н.э.), центр которой находился
в Огайо; эта культура славилась трубками из рез-
ного камня, медными изделиями и ритуальными
земельными курганами.
3. Разновидность повозки; на таких спаренных
шестах, которые волокла собака, перевозили свои
грузы жившие в вигвамах охотники на бизонов Ве-
ликих равнин.
4. Ручка ложки; этим стилизованный «бобром»
тлингитской культуры представлена искусная резь-
ба по дереву индейцев северо-западного побережья.
5. Коробочка из бересты; индейцы восточных ле-
сов использовали бересту для изготовления коробок,
крыш жилищ и челноков (каноэ).
6. ![]()
![]()
![]() Голова, вырезанная из бивня моржа; эскимо-
Голова, вырезанная из бивня моржа; эскимо-
сы Северо-Американского Заполярья занимались
резьбой по моржовой кости еще 2 тысячи лет назад.
Доколумбовы произведения
искусства.
Нумерация соответствует
принятой в тексте.
1. Голова, высеченная из камня.
2. Бог маиса.
3. Нож из обсидиана.
4. Каменная статуэтка пумы.
5.
Ваза в форме головы
человека.
6. Золотая маска.
7.
![]() Золотая статуэтка ламы.
Золотая статуэтка ламы.
К северу от Мексики земледельческие деревни и
поселки оставались крупнейшими поселениями
вплоть до прихода европейцев. Но к югу от Мексики
индейцы создавали города, государства и империи,
основанные на сельском хозяйстве, торговле и завое-
ваниях. Это происходило в двух районах: в Цент-
ральной Америке, или Мезоамерике (от Мексики до
Г ондураса), а также в Центральных Андах и на близ-
лежащих территориях. Независимо от ранних куль-
турных центров Старого Света здесь тоже появи-
лись социальные классы со своими монархами, жре-
цами, солдатами, ремесленниками и крестьянами.
Самостоятельно они создали замечательную кера-
мику, скульптуру, живопись, открыли технологию
производства бронзы, возвели пирамиды и холмы
типа платформ из кирпича или камня. Обитатели
Центральной Америки даже придумали пиктогра-
фическое письмо. Но все же американские индейцы
никогда не овладели технологией производства же-
леза и применяли колеса только в детских игрушках.
На следующих иллюстрациях представлены основ-
ные стадии культурного развития после 2000 г. до н. э.
|
|

|
|

| 1. Высеченная из камня голова; этот монолит вы- 2. Бог маиса, из религиозной книги индейцев майя. 3. Нож из обсидиана; ацтеки пользовались им 4. Пума из камня, культура шавин (900—200 г. 5. Портретная ваза; богатая культура мочика, су- 6. Золотая маска; работа индейцев чиму из Перу. 7. Золотая лама; изготовлена мастерами империи |
Места обитания пигмеев.
На этой карте показаны места
распространения пигмеев.
1. Бассейн Конго (Заир).
2. Штат Керала в Индии.
3. Шри-Ланка.
4. Андаманские острова.
5. Малайзия.
6. Остров Палаван.
7. Остров Лусон.
8. Остров Минданао.
9. Остров Сумбава.
10.
надо было потреблять больше пищи, чем они могут
вырастить. Эти люди не настолько мелкие, чтобы они
не смогли выполнять необходимую работу.
Улучшение же питания приводит к увеличению
роста тех групп, где он не ограничен генетически.
Одно исследование, проведенное в 60-х годах нашего
века, показало, что дети итальянских иммигрантов
в США часто имеют больший рост, чем их родители.
Потребление вареной пищи и применение орудий
изменили человеческое лицо. Питание мягкой варе-
ной пищей и использование орудий для выполнения
тех видов работ, которые когда-то осуществлялись
зубами, привело к уменьшению большинства лице-
вых структур, предназначенных в основном для под-
держания жевательного аппарата. Зубы уменьши-
лись, в особенности среди тех популяций, которые
длительное время ели вареную пищу. Самые малень-
кие зубы у населения Европы, Ближнего Востока и
Китая, а самые крупные — у австралийских абори-
генов. Начиная с бронзового века нижние челюсти
имели тенденцию уменьшаться в размерах до тех
пор, пока зубы верхней челюсти не начали перекры-
вать нижние зубы и правильный прикус не уступил
место неправильному.
![]() Со времен позднего палеолита все возрастаю-
Со времен позднего палеолита все возрастаю-
щее применение орудий и приспособлений в общем
уменьшило потребность в мощных мышцах и проч-
ных костях. Соответственно этому и скелеты стано-
вились все менее крепкими.
![]()
![]()
|
3 миллиона лет назад в мире обитало, возмож-
но, всего лишь несколько десятков тысяч гоминид,
являвшихся нашими предками. К 1980 г. н. э. на Зем-
ле проживало 4 миллиарда 500 миллионов человек.
Но на протяжении большей части доисторического
времени численность населения возрастала мед-
ленно. Рост происходил поэтапно по мере эволюции
наших предков, колонизации новых земель и увели-
чения пищевых ресурсов.
Согласно одному из подсчетов, 2 миллиона лет
назад 1 миллион травоядных австралопитеков оби-
тал только в нескольких местах на одном континенте.
500 тысяч лет назад занимавшийся охотой и соби-
рательством Homo erectus населял уже самые раз-
ные ландшафты Африки и значительной части Евра-
зии. Но ограниченный участок земли может пропи-
тать собирателей-охотников даже меньше, чем тра-
воядных существ. Поэтому некоторые ученые счи-
тают, что в мире в то время проживало всего лишь
1,7 миллиона существ вида Homo erectus.
10 тысяч лет назад Homo sapiens уже колонизо-
вал все континенты и климатические зоны, за исклю-
чением Антарктиды. И все же большинство людей
продолжали вести кочевой образ жизни охотников-
собирателей, неспособных регулировать свои пище-
вые ресурсы. В мире в то время насчитывалось са-
мое большее 10 миллионов человек.
Население различных
континентов.
На этих полосах, разделенных
на части, показана
приблизительная дола
населения Земли, которая
приходилась на тот или иной
континент в одну из семи
произвольно выбранных дат
начиная с 10 000 г. до н. э.
а — Океания (включая
Австралию).
б — Северная и Южная
Америка.
в — Африка.
г — Азия.
д — Европа.
| а б в г д
|
|||||||||||||||||
Количество населения резко возросло всего лишь
около 7 тысяч лет назад, когда разведение земле-
дельческих культур привело к увеличению производ-
ства пищи, и те люди, которые раньше погибли бы
от голода, теперь оставались в живых. Новые ору-
дия и технологии, описанные вкратце в этой главе,
способствовали дальнейшему росту пищевых ресур-
сов и численности человечества. К 500 г. до н. э. (а по
мнению некоторых ученых, к 2500 г. до н. э.) населе-
ние Земли превысило 100 миллионов. Позже обмен
земледельческими культурами и домашними живот-
ными в глобальных масштабах, промышленная тех-
нология и научная медицина еще более ускорили
рост населения. К 2000 г. на этой перенаселенной
планете, возможно, будут тесниться бок о бок 6 мил-
лиардов человек.
Население Земли.
На этих двух диаграммах
показан предполагаемый рост
населения Земли.
А — От 15 000 лет до н. э. до
1 г. н. э.
Б — От 1700 до 2000 г. н. э.
![]() =
2,2 миллиона
=
2,2 миллиона
![]()
![]() 10000
10000
134
|
|
|
|
ЫЖЖЖЖЖЖЖЖЖЖЖЖЖЖ '500 6000
Исчезновение охотников.
На этой карте мира показано
сокращение числа сообществ
охотников-собирателей.
Исчезновение людей, ведущих
такой образ жизни, означает
конец влияния факторов
естественного отбора, которые
способствовали эволюции
нашего вида. Показанные более
темным цветом районы еще
200 лет назад были заселены
группами охотников-
собирателей. Точками отмечены
районы, где в последние
десятилетия еще сохранились
охотники-собиратели.
1. Индейцы варау.
2. Индейцы шириана.
3. Индейцы бороро.
4. Негриллы.
5. Хадза.
6. Бушмены.
7. Бирхор.
8. Сакай.
9. Семанг.
10. Пунан.
11. Тасадай.
12. Австралийские аборигены.
Из группы приматов, слабо защищенных от воз-
действий внешней среды, эволюция отобрала наш
плодовитый вид, обладающий уникальной способ-
ностью к размножению, миграции и преобразованию
нашей планеты.
Продолжится ли эволюция человека? В настоящее
время мы должны ответить: «нет». Культурная эво-
люция защитила нас от биологических перегрузок,
которые устраняли слабых, медлительных и плохо
соображающих особей. Теперь применение машин,
компьютеров, одежды, очков и современная меди-
цина обесценили прежние, наследовавшиеся людьми
преимущества, связанные с мощным телосложением,
интеллектуальными способностями, пигментацией,
остротой зрения и устойчивостью к таким заболе-
ваниям, как, скажем, малярия. В каждом обществе
имеется высокий процент физически слабых или
плохо сложенных людей, а также людей со слабым
зрением или таким цветом кожи и сопротивляемо-
стью заболеваниям, которые не соответствуют кли-
матическим условиям той местности, в которой они
живут. Физически несовершенные люди, которые
100 лет назад умерли бы в детстве, теперь выживают
и дают потомство, передавая будущим поколениям
свои генетические дефекты.
Приостановке эволюции человека способствовала
и миграция. Сейчас уже ни одна из групп населения
|
|
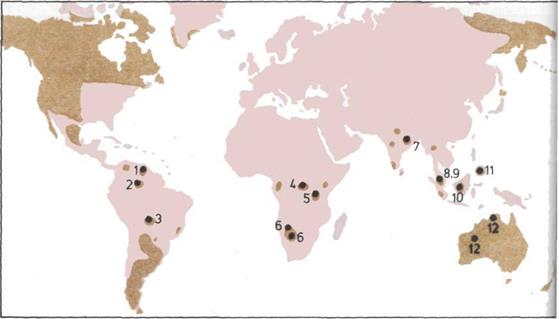
Земли не живет в условиях изоляции достаточно
длительное время, необходимое для ее превращения
в новый вид, как это происходило в эпоху плейсто-
цена. А расовые различия будут сглаживаться по
мере роста числа смешанных браков между пред-
ставителями народов Европы, Африки, Америки, Ин-
дии и Китая.
В сущности, вымирание человечества представля-
ется более вероятным, чем его дальнейшая эволюция.
Резко возрастающее население находится в зависи-
мости от увеличения производства пищи и энергии.
Но и то и другое требует чрезмерной эксплуатации
природных ресурсов нашей планеты. Усиленная об-
работка почвы приводит к ее эрозии, что снижает
плодородие, а истощение запасов ископаемого топ-
лива представляет угрозу для энергоснабжения.
Климатические изменения могут усугубить обе эти
проблемы. Чрезмерно многочисленный, испытываю-
щий нехватку в продуктах питания и топливе вид
Homo sapiens может вымереть от голода, войн или
эпидемий.
![]()
Продолжительность существования большинства
видов животных составляет менее 3 миллионов лет.
Нашему виду — самому разумному из всех — может
потребоваться вся его изобретательность, чтобы про-
держаться хотя бы половину этого срока.
♦♦t щ
Смешение рас (вверху).
Скрещивание расовых групп
(А, Б, В) увеличивает
разнообразие генов (а)
в популяции. Гены,
унаследованные отдельными
людьми, определяют их
характерные особенности.
![]()
![]() Таким образом, смешение
Таким образом, смешение
рас может привести к появлению
белокурых негров или
рыжеволосых китайцев.
![]() Глава
11 ПОЗНАНИЕ ИСТОРИИ ЧЕЛОВЕЧЕСТВА
Глава
11 ПОЗНАНИЕ ИСТОРИИ ЧЕЛОВЕЧЕСТВА
На следующих страницах
дается краткий обзор сведе-
ний о древних людях, а так-
же описание техники и ме-
тодов, которые помогают уче-
ным интерпретировать эти
данные. Книга заканчивается
краткими биографиями неко-
торых из тех многих людей,
которые значительно спо-
собствовали разгадке тайн
прошлого; приводится также
путеводитель по нескольким
мировым музейным коллек-
циям остатков ископаемых
приматов и предметов д о
-
исторической материальной
культуры. Не все из них от-
крыты для регулярного посе-
щения широкой публикой, и
поэтому прежде, чем отпра-
виться туда, надо уточнить
возможность открытого дос-
тупа.
Этот поперечный разрез
доисторических погребений был
приведен в книге английского
генерала XIX века Огастаса
Питт-Риверса — одного из
основателей современной
научной археологии.
|
|

Образование ископаемых остатков
|
|
|
|
лотных вод, которые приводят к растрескиванию,
расслаиванию (шелушению) и растворению костных
остатков. Захоронение также замедляет процессы
гниения.
Со временем белок костных остатков исчезает, воз-
можно замещаясь растворенными минералами,
содержащими такие вещества, как кальций или крем-
ний. Минералы делают кости более твердыми, тяже-
лыми и прочными, и кости лучше выдерживают вес
подводных слоев ила или песка, осаждающихся
поверх них. Значительное давление и естественные
цементирующие вещества медленно уплотняют эти
осадки, превращая их в слоистые известняковые,
глинистые или песчанистые породы — массивные
гробницы, в которых замурованы ископаемые ос-
татки.
![]() Ископаемые кости и зубы — это обычно единствен-
Ископаемые кости и зубы — это обычно единствен-
ное, что непосредственно осталось от древнего чело-
века. Однако в некоторых специфических условиях
кожа и даже мясо человека сохранялись 2 тысячи лет
и больше. Мы встречаем иссохшие мумии в древ-
них захоронениях Египта и Перу, где сухой воздух
пустыни защитил от разложения и сохранил их кожу
и волосы. Еще более замечательны выглядящие
почти так же, как при жизни, трупы удавленных
жертв, которые были брошены в болота около 2 ты-
сяч лет назад в таких районах, как Скандинавия
и Ирландия. Влажная кислая почва, не дающая до-
ступа воздуху, способствовала сохранению гибкости
и мягкости кожи этих трупов и предохранила их от
разложения. Почти столь же удивительны трупы,
возраст которых исчисляется в 2400 лет, из долины
Пазырык в советской Центральной Азии (на Алтае.—
Прим. пер.). Здесь лёд, образовавшийся в могилах
древних вождей, предохранил от разложения чело-
веческую кожу, покрытую татуированными изобра-
жениями самых разных животных.
|
|
| Они сохранились во льду А — В заполненной льдом |
| обнаружены остатки человека, |
| Б — Человек из Толлунда |
| 15 За к. 5110 |
Судьба ископаемых остатков.
![]() 1. Мертвый австралопитек
1. Мертвый австралопитек
лежит на дне озера. Мягкие
части тела разлагаются, кости
остаются в сохранности.
Палеонтологи, как правило, находят ископаемые
остатки только в тех случаях, когда они встречаются
на земной поверхности или недалеко от нее. Большин-
ство ископаемых обнажается в результате измене-
ний, происходящих в земной коре. Поднятие горных
пород, содержащих ископаемые остатки, или опуска-
ние расположенных по соседству глыб приводит к
эрозии покрывающих слоев осадков. Жар и холод,
ветер и дождь разрушают горные породы, находя-
щиеся на земной поверхности, а затем сметают или
смывают их, обнажая более глубокие и обычно более
старые породы вместе с содержащимися в них иско-
паемыми остатками.
Например, в результате поднятия гор Сивалик в
Северном Пакистане обнажились остатки ископае-
мых человекообразных обезьян Ponginae возрастом
от 8 до 14 миллионов лет. Смещение блоков земной
коры в гигантской Рифтовой долине в Африке при-
вело к понижению уровня озера Туркана, что осу-
шило древнее «озеро Олдувай». В результате были
обнажены древние прибрежные отложения, содержа-
щие ископаемые костные остатки австралопитеков
и видов людей Homo habilis и Homo erectus, вместе
с костями тех животных, на которых люди охотились
или трупы которых они поедали, а также оббитые
каменные отщепы, которые использовались в каче-
стве орудий.
В поисках древнего человека палеонтологи изу-
чают отложения, сформировавшиеся на континентах
примерно за последние 5 миллионов лет. Кроме бе-
регов древних озер к возможным местам находок
относятся также пещеры, сухие овраги и обширные
волнистые полупустыни (например, в Эфиопии и Ке-
нии). Неожиданные ценные материалы можно иног-
да обнаружить в карьерах, шахтах и горных выра-
ботках.
Поиски ископаемых гоминид часто представляют
собой медленную, трудоемкую работу. Исследовате-
Поднятие слоев земной коры
по разломам
(справа).
На этом рисунке показано, как
в результате скольжения вдоль
разлома — линии наименьшего
сопротивления в земной
коре — на поверхности могут
оказаться захороненные
отложения, содержащие
ископаемые остатки.
А — Опущенный вниз блок
горной породы.
![]() Б — Взброшенный вверх блок
Б — Взброшенный вверх блок
горной породы.
а — Содержащий ископаемые
остатки слой горной породы,
который обнажился в результате
взброса.
лям иногда приходится тщательно просматривать
обширные склоны и большие груды пород в поисках
маленьких предметов необычной формы или цвета:
ископаемые кости человека иногда бывают серого
или белого, но часто и черного цвета и встречаются
обычно в виде изолированных фрагментов. Иногда
проходят годы, прежде чем начинающий специалист
обретет надлежащий опыт. Прочесывание же пустых
участков земли, где возможны находки, требует
терпеливого труда целых поисковых групп. В Север-
ной Кении, чтобы обнаружить остатки 200 гоминид,
группе из шести человек пришлось работать в тече-
ние 11 лет, просматривая по 300 квадратных миль
(800 км~) в год. Только девять из этих ископаемых
остатков являлись черепами; остальные представ-
ляли собой отдельные зубы или обломки костей. До
времени неандертальцев — первых людей, которые
начали хоронить своих мертвых, находки целых чело-
веческих скелетов неизвестны.
Стоянки, обнаруженные
в районах разломов
в Африке.
А — Район исследования
в Африке.
Б — Система африканской
Рифтовой долины, где
в результате гигантских
подвижек земной коры по
разломам было обнажено
много стоянок древних
ископаемых гоминин. Ниже
перечислены восемь
основных районов стоянок
с указанием возраста
ископаемых находок в
миллионах лет.
1.
Район Гадара в Афарском
треугольнике; 3,1—2,6.
2. Бассейн реки Омо; 2,9—1,0.
3. Нариокотоме; 1,6.
4.
Восточный берег озера
Туркана; 2,4/1,8—1,3.
5. Лотегем; 5,5.
6. Кананой; 4
7.
![]()
![]() Олдувайское ущелье; 1,85
Олдувайское ущелье; 1,85
и до настоящего времени.
8. Лаэтоли; 3,5 и ранее.
Найденные на поверхности земли, беспорядочно
разбросанные ископаемые костные остатки или ка-
менные орудия дают нам только отрывочные све-
дения о древнем человеке. Но случайные находки
на поверхности могут привести к обнаружению ко-
стей, предметов обихода и пыльцы, которые отно-
сятся к одному и тому же месту и времени и хорошо
сохранились в древних отложениях. Тщательные
раскопки таких участков могут дать нам пенные
сведения о наших древних предках, их образе жиз-
ни, возрасте и окружающей их среде.
Раскопки начинаются, как правило, с картирова-
ния участка, который предстоит исследовать. Па-
леоантропологи или археологи разбивают его сеткой,
ячейки которой имеют стороны длиной 1 м. Сначала
собирают находки, встречающиеся па поверхности.
Затем раскапывают квадраты в виде блоков или
траншей, вскрывая отложения по одному слою за
раз. Для удаления перекрывающих горных пород
могут быть использованы кирки и лопаты. Затем ра-
бота замедляется, так как искатели начинают дей-
ствовать совочками и зубилами. Зондами и кисточ-
ками удаляют вмещающую породу, приставшую к
предметам, а при помощи ситечек отделяют мелкие
зубы и фрагменты костей.
Рабочие тщательно нумеруют каждый ископае-
мый остаток, бытовой предмет или камень, отмечают
его положение на карте и записывают, в каком слое
он был найден. Хрупкие предметы покрывают пре-
дохраняющим веществом.
Объемы работы могут быть неимоверно велики.
Так, за 46 месяцев одна партия работников напита
32 тысячи ископаемых остатков и 37 тысяч предме-
тов материальной культуры, залегавших па 43 уров-
нях 13 участков в Олдувайском ущелье в Танзании.
Удаленной пустой горной породой можно быио бы
заполнить траншею длиной 1 миля (1,6 км), шириной
10 футов (3 м) и глубиной 10 футов (3 м).
При некоторый раскопках приходится прибегать
к необыиным методам. Так, например, в южноафри-
канских пещерах встречаются хрупкие кости австра-
лопитеков, залегающие в естественном цементирую-
щем веществе под названием брекчия, которую мож-
но разбивать только на отдельные монолитные куски;
на извлечение костей в сохранном виде уходят целые
недели. Исследование затопленный участков связано
с подводныши работами: сменяющие друг друга груп-
пы! археологов, обученных подводному плаванию,
просеивают ил или глину с помощью специальный
приспособлений, определяют местоположение нахо-
док и поднимают их на поверхность.
![]()
![]()
![]()
![]() Раскопки это только первый шаг. За ними сле-
Раскопки это только первый шаг. За ними сле-
дует работа по интерпретации данный и публикации
новый открытий.
|
|

| >.;i= Аз*- 3fasf #•■ '^, d«44V4 , ■■ **d£': \7"л '%, V !•"■ ■' " - * ^Л ^Тл S*r* *> ■•" "^ЛГЛ W v- u v »• » ( W. - ■ ■' r * iu |
| _ v' "■■ ; W- ■'\S- Дк( |
| -"( к ? / |
| *I« |
| *r^7 '_ .„ ' J 4Y |
| Раскопки карстовых пещер. Раскопки южноафриканских |
![]()
Реконструкция проконсула
(справа).
![]()
![]() На этой реконструкции
На этой реконструкции
ископаемого гоминоида
Proconsul окрашенными
участками показаны
найденные фрагменты костей.
Остальные участки были
восстановлены
предположительно.
Реконструкция ископаемых
остатков
Реконструкция ископаемых черепов и конечно-
стей и восстановление утраченных мягких частей
тела помогают нам представить себе внешний вид
доисторических предков современного человека.
Прежде всего музейные препараторы очищают
кости. С помощью сверления, вибрации и песко-
струйной очистки удаляют прилипшую к образцам
песчаную породу. Несколькими повторными промы-
ваниями и обработкой уксусной кислотой освобож-
дают ископаемые остатки, сцементированные брек-
чией. Влажные костные остатки можно высушить,
подвесив их в н-бутиловом спирте. Лабораторные
специалисты закрепляют хрупкие пористые кости с
помощью искусственных пластмасс, растворенных
в различных растворителях, или с помощью эмуль-
сий, разведенных в воде.
Затем искусные анатомы соединяют фрагменты
костей, подобно частям головоломки, и реконструи-
руют череп или скелет, заменяя недостающие части
пластмассовыми имитациями. Форма и размеры
определенных костей показывают, как ископаемое
существо держало голову и конечности. На изготов-
ление и монтирование одной реконструкции могут
уйти недели или даже месяцы старательного труда.
|
|
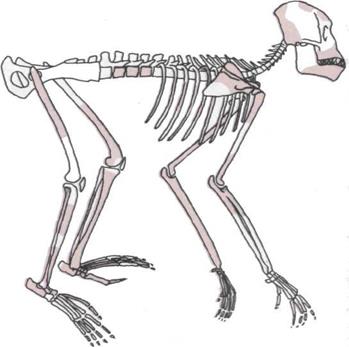
По черепам и скелетам художники и скульпторы,
специализирующиеся в анатомии, могут реконструи-
ровать голову и все тело. По прочности и размерам
костей конечностей можно судить о величине мышц,
приводивших их в движение. По шишкам, углубле-
ниям и другим особенностям поверхности костей
можно говорить о месте прикрепления мышц и об их
форме. Так, например, по костным буграм и греб-
ням на черепе видно, что крупные мышцы удержива-
ли голову с сильно выступающей вперед лицевой
частью или приводили в движение мощные челюсти.
Все это помогает художникам облечь плотью череп
или скелет, сначала добавляя мышцы и жировой
слой, а затем кожу и волосы.
Но все же воссоздание полного облика ископае-
мого существа связано с большей неопределен-
ностью, чем реконструкция скелета по его сохранив-
шимся частям, поскольку мягкие ткани не всегда
оставляют свои отметки на костях. Два скульптора,
работающие с черепом одного и того же гоминида,
могут в результате воссоздать две очень отличаю-
щиеся одна от другой головы, в разной степени похо-
жие на человеческую. Точно так же — лишь при по-
мощи проницательных догадок — можно восстано-
вить кожу, глаза, цвет волос и волосяной покров
тела.
Реконструкция черепа
(вверху).
![]() Восстановление черепа по
Восстановление черепа по
фрагментам его костей
напоминает решение
головоломки (воссоздание
трехмерной загадочной
картинки по отдельным ее
кусочкам). Руководствуясь
размером, формой и изгибом
фрагментов костей, анатом
прилаживает косточки
к сделанному из мягкой глины
макету головы, изменяя его
форму так, чтобы она
соответствовала изгибам
костей реконструируемого
черепа.
![]()
|
|
О чем говорит стертость зубов
(внизу).
Сравнивая сильно увеличенные
фотоснимки поверхности зубов
вымерших и современных
видов, ученые определяют, чем
питались предки нынешних
животных.
1.
Зубы травоядного —
с мелкими бороздками
(не встречающимися у гоминид).
2.
Зубы животных, питавшихся
плодами, — гладкие с точечками
(встречаются у орангутана
и австралопитеков).
3. Зубы плотоядных
![]() и всеядных — с глубокими
и всеядных — с глубокими
ямками н царапинами,
образовавшимися при
разгрызании костей и хрящей
(встречаются у гиен и человека
прямоходящего — Homo
erect us).
Кости и предметы материальной
культуры
Для искусных истолкователей костные остатки и
предметы материальной культуры являются энци-
клопедией древней жизни.
Сравнение размеров и строения десятков костей
и зубов позволяет ученым определить, что фрагмен-
ты черепа и костей конечностей принадлежали не-
андертальцам или, скажем, дриопитекам.
Анатомические данные говорят нам о физических
возможностях людей и их образе жизни. По длине
костей можно судить о телосложении и росте. По пле-
чевому поясу видно, какие из приматов были способ-
ны висеть на ветках деревьев. Положение отверстия
в черепе, через которое проходит спинной мозг, гово-
рит о том, держало ли данное существо голову прямо
или она была выдвинута вперед. По костям и суста-
вам большого и прочих пальцев можно судить о спо-
собности сжимать предметы рукой и точно их схва-
тывать или об отсутствии такой способности, как это
было у ранних приматов. По костям бедер и пальцев
ног можно отличить четвероногих, лазавших по де-
ревьям, от двуногих ходоков, и прямоходящих — от
тех, чьи тела кренились при ходьбе из стороны в сто-
рону.
По черепу можно оценить объем мозга, зрение и
обоняние. Челюсти и зубы, особенно их строение, раз-
меры, форма и степень истирания, убедительно сви-
детельствуют о том, чем питались их обладатели.
Но ископаемые костные остатки могут нам расска-
зать кое-что и сверх этого: специалисты могут делать
по ним выводы о некоторых видах ран и заболеваний,
а также о том, насколько успешным было выздоров-
ление жертвы. По зубам и длинным костям можно
определить возраст человека в момент смерти.
Дальнейшие сведения о доисторических гоминидах
можно получить, изучая предметы их материальной
культуры, вначале изготовляемые главным обра-
зом из камня, кости и древесного угля, а позже —
|
|

гончарные изделия; исследуются также углубления,
оставшиеся от столбов. По всем этим данным можно
судить о том, как развивалась техника, как древний
человек открывал все более эффективные способы
добывать пищу, согреваться и сооружать укрытия.
то, какими орудиями пользовался древний человек.
Экспериментаторы вновь открывают способы изго-
товления и применения каменных орудий нашими
предками. И наконец, ископаемые орудия и живот-
ные остатки со стоянок охотников «современного
палеолита», таких как эскимосы, позволяют соста-
вить представление об общественной жизни доисто-
рического человека. Сравнение остатков в лагерях
эскимосов и древнего человека помогает ученым
оценить численность групп доисторических охотни-
ков, определить те виды деятельности, которыми
занимались их члены, установить длительность их
пребывания на отдельных стоянках и узнать время
года, когда это происходило.
Царапины на кости.
![]() Короткие поперечные царапины
Короткие поперечные царапины
(а) на этой кости антилопы нз
Олдувайского ущелья
образовались в тех местах, где
древний гоминид срезал с нее
заостренным камнем кусочки
мяса. Царапины расположены
поверх следов зубов хищника,
и можно предположить, что эти
древние люди, скорее всего,
не занимались охотой сами,
а поедали мясо животных, уже
убитых хищниками.
Ключи к познанию окружающей
среды
О чем может рассказать пыльца
древних растений (вверху).
По пыльце из одного торфяного
болота в Дании можно судить
о том, что на этом месте
4300 лет назад местные
земледельцы временно
расчистили лес под пашню.
Высота закрашенного поля на
диаграмме изменяется
в зависимости от численности
в тот или иной период тех
растений, которые эта
диаграмма представляет.
А — Древний лес.
Б — Расчищенный участок.
В — Участок, вновь заросший
лесом.
а — Вяз, липа и ясень.
б — Дуб.
в — Береза.
г — Травянистая
растительность.
д — Дикие травы.
е — Хлебные злаки.
ж — Подорожник (сорняк среди
![]()
![]() посевов).
посевов).
По древним формам рельефа, ископаемым расте-
ниям и животным можно судить о климатических
условиях и характере местности, в которой обитали
древние люди. Вот всего лишь несколько примеров.
Тщательное исследование слоистых осадков поз-
воляет обнаружить древние озера, реки и скалы. Не-
нарушенные осадки мелких частиц накапливаются на
дне озер. Крупнозернистые осадки, связанные при-
родными цементирующими веществами, могут пред-
ставлять собой ложе древних рек. По песчаной ряби
на поверхности осадков древних рек можно делать
выводы о скорости и направлении их течения. Слои
затвердевшей почвы между корнями трав свидетель-
ствуют о дождливых периодах, когда происходил
нанос осадков вышедшими из берегов озерами или
реками, и о перемежающих их сухих периодах. По
пещерам в прибрежных морских утесах, располо-
женным выше современного уровня моря, можно
судить о положении древнего берега в те времена,
когда уровень моря был выше современного.
![]()
![]() Листья, стебли, корни и пыльца ископаемых рас-
Листья, стебли, корни и пыльца ископаемых рас-
тений могут рассказать о многом другом. Последо-
вательные остатки пальм, дубов и лиственниц, кото-
рые находят на одном и том же месте в покрываю-
щих друг друга слоях, говорят о похолодании кли-
мата. Ил ледникового озера, перекрытый последо-
вательно слоями, содержащими березу, сосну, дуб
и липу, свидетельствует о потеплении климата, кото-
рое наступило вслед за последним ледниковым пе-
риодом. Смена пыльцы деревьев пыльцой зерновых
и сорных трав является существенным признаком
культурного разведения растений после расчистки
лесов.
Речные террасы.
Парные террасы по обоим
берегам реки соответствуют
изменениям уровня дна долины
в древности. Река прорезала
дно все глубже каждый раз.
Остатки животных также дают ценную информа-
цию о характере исчезнувших ландшафтов. Полосы
раковин в слоистых осадках указывают на распо-
ложение берегов древних морей. В известковых поч-
вах, где редко встречаются хорошо сохранившиеся
зерна пыльцы, о древней растительности можно су-
дить по улиткам. В доисторических лесах, тундре и
лугах были распространены различные виды улиток,
и их ракушки сохранились в почве на различных
уровнях.
в различных слоях на некоторых стоянках, можно
проследить изменение условий их обитания. Напри-
мер, в Олдувае в определенный момент влаголюби-
вые грызуны сменились слепышами, которые роют
ходы в сухой почве, что говорит о наступлении за-
сушливого периода. В Европе ледникового периода
в результате климатических изменений виды фауны,
характерные для теплой лесной местности, подоб-
нвге слону с прямвши бивнями, сменились такими
животными, как овцебык, волосатый мамонт и дру-
гие, которые бвии лучше приспособлены к суще-
ствованию в холодной степи и тундре.
А — Ледниковые периоды:
деревья немногочисленны.
1.
Норвежский лемминг
(холодные степи).
2.
Лесная мышовка (березовые
леса).
3.
Лесная мышь (лесистая
местность).
4.
Садовая соня (лесистая
местность).
5.
Земляные белки (холодные
степи).
6.
Сурок (щебнистые склоны
холмов).
7. Пашенная полевка (луга).
8.
Северная водяная полевка
(болота).
9.
Арктический лемминг
(холодные степи).
Б — Межледниковые периоды:
леса.
10.
Обыкновенная белка
(лесистая местность).
11.
Соня-полчок (лесистая
местность).
12.
Лесная мышь (лесистая
местность).
13.
Садовая соня (лесистая
местность).
14.
Рыжая полевка
(лесистая местность; остальные
виды полевок обитали в открытой
местности).
15.
Обыкновенная соня
(лесистая местность).
Относительная датировка — это просто опреде-
ление того, старше, младше или одновременно дан-
ное ископаемое, бытовой предмет или слой осадков
относительно какого-либо другого ископаемого,
предмета или слоя. Это еще ничего не говорит нам
о возрасте данного объекта, но ученые часто могут
соотнести его с другим объектом, возраст которого
установлен более точно. При этом может быть при-
менено несколько методов датировки.
![]() Биостратиграфический метод основан на срав-
Биостратиграфический метод основан на срав-
нении различных типов ископаемых животных или
растений, сохранившихся в разных слоях пород;
обычно более древние формы залегают глубже,
чем более поздние. Изучение эволюционной после-
довательности поколений доисторических диких
свиней и слонов помогло ученым соотнести между
собой слои, образовавшиеся в одно и то же время
на значительно удаленных друг от друга участках
Восточной Африки. В свою очередь это помогает
специалистам увязать значительно более редкие
находки ископаемого человека.
|
|
|
Палеомагнитный метод связан с периодическими
изменениями направления магнитного поля Земли
в прошлом. Намагниченные частицы в минеральных
отложениях, особенно в лавах, расположены в соот-
ветствии с тем магнитным полем Земли, которое
существовало в момент их образования. По этим
отложениям можно косвенно судить об относитель-
ном возрасте костных остатков и предметов мате-
риальной культуры, которые были в них захоронены
во время их формирования.
Ученые часто могут примерно определить возраст
находок даже по их внешнему виду. Тщательное
изучение морфологии кости (ее формы и строения)
позволяет отнести эти остатки к той эволюционной
последовательности, возраст которой уже был уста-
новлен стратиграфически. Аналогичным образом
типологическое исследование — классификация
предметов материальной культуры по их форме —
способно помочь археологам датировать каменные
предметы исходя из того, как они изменялись во
времени.
Относительная датировка костных остатков осно-
вана на сравнении химических веществ, содержа-
щихся в данной ископаемой кости, с теми вещества-
ми, которые есть в других костях, возраст которых
установлен. Кости поглощают фтор и уран из грунто-
вых подземных вод. Чем более длительному захоро-
нению подверглась кость, тем большее количество
фтора или урана в ней накопилось. Поэтому измере-
ние содержания фтора или урана в кости либо уровня
ее радиоактивности дает представление об относи-
тельном возрасте кости. Но это верно лишь по отно-
шению к костям из одной и той же местности и нахо-
дившимся в одних и тех же условиях.
Пилтдаунская подделка.
![]() Найденные около Пилтдауна
Найденные около Пилтдауна
в Юго-Восточной Англии
остатки черепа: человеческая
черепная коробка
и обезьяноподобная челюсть
«эоантропа» — озадачили
специалистов. Позже,
определив количество фтора
в этих костях и подвергнув их
другим исследованиям, ученые
установили, что эта находка
представляет собой
мистификацию: она была
составлена из костей
современного человека
и орангутана.
С помощью методов хронометрической датировки
устанавливается приблизительный абсолютный воз-
раст доисторических предметов. Большинство из
перечисленных ниже методов связано с измерением
тех изменений, которые происходят с постоянной ско-
ростью в строении атомов вещества.
1. Дендрохронология основана на подсчете еже-
годных колец в древесине тех деревьев, периоды
существования которых частично перекрывали друг
друга. Применяется главным образом при датировке
некоторых доисторических участков возрастом до
6500 лет.
2. Варвовый анализ связан с подсчетом варв —
годичных лент, т. е. слоев осадков, отлагавшихся
в озере за один сезон в тех районах, где озера замер-
зали на зиму. Применяется главным образом при
датировке позднего плейстоцена.
3. Анализ соотношения изотопов кислорода в глу-
боководных осадках позволяет определить измене-
ние количества изотопа кислорода-18, которое кос-
венным образом связано с Глобальными колебания-
ми температуры. Применяется при датировке перио-
дов потепления и похолодания в эпоху плейстоцена
и раньше.
4. ![]()
![]()
![]()
![]() Датировка с помощью радиоуглеродного мето-
Датировка с помощью радиоуглеродного мето-
да основана на измерении количества радиоактивно-
го изотопа углерода (углерода-14) в органическом
веществе. В животных и растениях количество угле-
рода-14 после их смерти уменьшается с постоянной
скоростью. Применяется при датировке органиче-
ских веществ (например, костей) возрастом до 50
тысяч лет.
Определение возраста
радиоуглеродным методом
(справа).
Тело мертвого организма теряет
аккумулированный им
углерод-14 со скоростью,
которую можно измерить, что
позволяет определить возраст
данного организма. Пределы
погрешности возрастают при
увеличении временного
интервала.
1.
Через 5730 лет после смерти
организма в его теле
сохраняется половина
углерода-14.
2.
Через 16 704 года после
смерти сохраняется одна восьмая
углерода-14.
3.
Через 70 тысяч лет после
смерти исчезает почти весь
изотоп. Но применяя более
изощренные методы датировки,
можно определить возраст
![]() и более древних предметов
и более древних предметов
органического происхождения.
5. Термолюминесцентный метод датировки связан
с измерением световой энергии, излучаемой при на-
гревании определенных минералов, которые когда-
то в доисторические времена уже подвергались нагре-
ванию. Энергия испускаемого света говорит о том,
когда произошло нагревание в доисторические вре-
мена. Применяется по отношению к обожженным
глиняным черепкам, глиняным статуэткам, камням
из доисторических костров и т. д. возрастом до 8 ты-
сяч лет.
6. Аминокислотная рацемизация происходит в
организмах после их смерти. Оптически активные
белковые аминокислоты (изменяющие направление
поляризованного света) подвергаются медленной
рацемизации, т. е. превращаются в оптически ак-
тивные. Метод применяется по отношению к кост-
ным остаткам возрастом до 100 тысяч лет и более.
7. Датировка по изотопам уранового ряда основа-
на на скорости радиоактивного распада урана-234
и превращения его в протактиний-231, а затем в то-
рий-230; этот метод позволяет устанавливать воз-
раст событий, имевших место от нескольких тысяче-
летий до 350 тысяч лет назад.
8. При датировке по следам распада измеряется
распад урана в древних каменных предметах; изме-
рение выполняется по числу следов, вызванных рас-
щеплением урана-238, которое происходит в природе
с известной постоянной скоростью. Применяется по
отношению к стоянкам древнего человека.
9. Калий-аргоновый метод датировки основан на
постоянном превращении калия-40 в аргон-40. При-
меняется по отношению к вулканическим породам,
возраст которых превышает 1 миллион лет.
Для палеонтологов особенно пригодны методы 4, 7,
8 и 9. Но загрязнение изучаемых образцов другими
веществами может привести к искажению результа-
тов, и поэтому специалисты перепроверяют получен-
ные данные, используя при возможности несколько
различных методов.
Калий-аргоновый метод
датировки.
Калий-40 превращается в
аргон-40 и кальций-40 со
скоростью, показанной на этой
диаграмме.
1. Содержание калия-40
в только что сформировавшейся
вулканической породе.
2.
Доля калия-40, оставшаяся
после одного периода
полураспада (1310 миллионов
лет).
3.
Доля калия-40. оставшаяся
после следующих 1310 миллионов
лет (после двух периодов
полураспада).
4.
Доля калия-40, оставшаяся
после еще одного периода
1310 миллионов лет (после трех
периодов полураспада).
|
|
Трудности, связанные
с изучением ископаемых
остатков
Несмотря на методы, которые мы только что пере-
числили, поиски предков вида Homo sapiens похожи
на разгадку головоломок, ключи к решению которых
утеряны.
Многие находки остатков ископаемых приматов
представляют собой всего лишь фрагменты нижней
челюсти или зубы, т. е. наиболее долговечные части
скелета. Большинство других костей и, конечно, все
мягкие части тела уже давно исчезли: они были рас-
терзаны хищниками, а потом растащены в разные
стороны другими животными или унесены водой.
Сохранилось очень мало полных скелетов древних
приматов.
Из-за такой скудости ископаемых находок палео-
антропологи вынуждены определять древних гоминид
по неполным остаткам, иногда принадлежащим осо-
бям разного иола и возраста, которые могут сильно
различаться в результате костных заболеваний.
Поэтому не удивительно, что специалисты расхо-
дятся во мнениях относительно того, кому принад-
лежат те или иные ископаемые остатки.
![]()
![]()
![]()
![]() Еще труднее ответить на вопросы об образе жизни
Еще труднее ответить на вопросы об образе жизни
древних людей, так как их действия в большинстве
случаев не оставляли ископаемых следов и сохрани-
лось мало древних орудий, изготовленных из такого
недолговечного материала, как дерево. Мы можем
только догадываться, что древний человек, подобно
некоторым современным людям, ведущим примитив-
ный образ жизни, занимался собирательством и
пользовался при этом лотками, изготовленными из
коры, чтобы уносить «домой» собранные им корни
или плоды.
Кроме того, полное отсутствие находок, относя-
щихся к тем или иным периодам либо районам, остав-
ляет мучительные пробелы в истории ископаемых
приматов и приводит к неправильному воссозданию
недостающих звеньев. Даже датировка ископаемых
остатков часто бывает затруднена из-за наличия
в образцах посторонних примесей или из-за смеще-
ния земных слоев, нарушившего стратиграфическую
последовательность пород.
И тем не менее наши знания о древних людях бы-
стро растут в результате исследования ископаемых
остатков, развития биохимии и благодаря сравни-
тельному изучению современных приматов, вклю-
чая и людей, ведущих примитивный образ жизни.
К 1968 г. во всей Кении было найдено всего лишь три
экземпляра остатков древних гоминид; к 1984 г. толь-
ко на стоянке Кооби Фора было сделано 150 находок.
К 2000 г. достижения техники и изучение еще плохо
исследованных отложений обещают дать значитель-
но больше материалов, что позволит уточнить, когда
разошлись линии эволюции человекообразных
обезьян и человека, как проходила эволюция челове-
ка в раннем плейстоцене, а также установить место
и время появления нашего подвида Homo sapiens.
Как исчезают древние остатки.
На этих трех рисунках видно,
почему ископаемые остатки
древних людей редко
встречаются в нетронутом виде.
1.
Гиены и грифы собираются
растерзать труп древнего
гоминида, лежащий на берегу
озера или реки. Позже его
разломанные кости потрескаются
на солнце и звери могут
растащить их в разные стороны.
2.
Во время паводков
изолированные обломки костей
покрываются слоями
тонкозернистых осадков, а зубы
и другие мелкие косточки
к тому времени, возможно, уже
были унесены рекой вниз
по течению, а потом выброшены
на илистую банку-отмель.
3.
Обнажившийся в пойме реки
в результате изменений ее русла
древний череп подвергся
сильному выветриванию прежде,
чем его нашли специалисты.
|
|

В поисках древнего человека (1)
Роберт Брум со слепком черепа
«плезиантропа»
(Australopithecus africanus).
На следующих четырех страницах пойдет речь о выдающихся
охотниках за ископаемыми — археологах и других ученых, кото-
рые занимались исследованием происхождения нашего вида и его
культурой на ранних этапах развития. (Фамилии приводятся
в порядке латинского алфавита. — Прим. пер.).
Такой ограниченный список, конечно, не может быть полным;
речь пойдет лишь о некоторых ученых, причем основное внимание
уделяется представителям западной науки.
Басе Джордж Флетчер (Bass, George Fletcher). Американский
археолог; его экспедиция 1960 г. по изучению корабля бронзового
века, потерпевшего крушение у берегов Турции, положила начало
научной подводной археологии.
Бинфорд Льюис P. (Binford, Lewis R). Американский археолог;
в 60-х годах нашего века стал основоположником так называемой
«новой археологии» после пересмотра старых концепций в свете
сравнительных археологических и этнографических исследований
в мировом масштабе.
Бинтам Хайрам (Bingham, Hiram) (1875—1956). Американский
археолог; в 1911 г. вторично открыл затерянный в джунглях
Перуанских Анд город инков Вилкабамба (Мачу-Пикчу), яв-
ляющийся одним из чудес доколумбовой цивилизации.
Блэк Девидсон (Black, Davidson) (1884—1934). Уроженец Ка-
нады, анатом; в 1927 г. дал описание найденного в Китае зуба
Sinanthropus pekinensis (Homo erectus). Позднее руководил
значительными раскопками пещерных стоянок Homo erectus на
территории Чжоукоудянь близ Пекина.
Болк Луис (Bolk, Louis). Голландский анатом; в 1926 г. вы-
двинул гипотезу, что человек — это неотенический тип челове-
кообразной обезьяны.
Буше (де Кревкёр) де Перт Жак [Boucher (de Crevecoeur) de
Perthes, Jacques] (1788—1868). Французский археолог; открыл
ручные топоры и ископаемые остатки животных в гравии реки
Сомма; развил мысль о том, что предыстория человека уходит
в далекое геологическое прошлое.
Буль Марселлин (Boule, Marcellin) (1861 —1942). Французский
палеонтолог; выполненное им неправильное описание неандер-
тальца способствовало распространению мнения о том, что это
было неуклюжее и звероподобное существо.
Брейдвуд Роберт Дж. (Braidwood, Robert J.). Американский
археолог; его раскопки города Джармо в Ираке в начале 50-х
годов нашего века показали, что сельское хозяйство на Ближнем
Востоке началось в горном поясе Таурус-Загрос, а не в богатых
долинах «Плодородного полумесяца», расположенного южнее.
Брестед Джеймс Генри (Breasted, James Henry) (1865—1935).
Американский археолог и египтолог; ввел термин «Плодородный
полумесяц» для обозначения простирающегося от Египта до
Ирака пояса плодородных земель, где было положено начало
западной цивилизации.
БрейльАнри (Breuil, Henri) (1877—1961). Французский священ-
ник и археолог; стал крупнейшим специалистом по пещерному
искусству эпохи палеолита.
Брум Роберт (Broom, Robert) (1866—1951). Шотландский палеон-
толог; открыл остатки австралопитеков в Южной Африке и в
1938 г. впервые описал вид Australopithecus robustus, который
он назвал Paranthropus robustus.
![]()
![]() Дарт Реммонд Артур (Dart, Raymond Arthur). Уроженец Австра-
Дарт Реммонд Артур (Dart, Raymond Arthur). Уроженец Австра-
лии, южноафриканский антрополог; в 1925 г. описал первого
найденного австралопитека Australopithecus africanus.
Реймонд Дарт со знаменитым
черепом молодого
австралопитека (слева).
Эжен Дюбуа, нашедший
человека прямостоящего
Homo erectus (справа )
Дарвин Чарльз Роберт (Darwin, Charles Robert) (18(19—1882).
Английский естествоиспытатель; своей теорией эволюции путем
естественного отбора (выдвинутой в 1859 г.) заложил научную
основу изучения происхождения человека, который, по его мне-
нию, впервые появился в Африке.
Доусон Чарльз (Dawson, Charles) (1864-1916). Английский
геолог-любитель; «открыл» так называемого «пилтдаунского
человека» "Eoanthropus dawsonii", которого палеонтолог Артур
Смит (Arthur Srrirth) назвал в 1912 г. недостающим эволюцион-
ным звеном; как выяснилось в 1953 г., после расследований, нача-
тых анатомом Джозефом С. Вейнером (Joseph S. Weiner),' череп
«пилтдаунского человека» оказался плодом мистификации'
Дугласе Эндрю Элликотт (Douglass, Andrew Ellicott) (1867—
1962). Американский астроном и археолог; разработал метод
дендрохронологии (датировки по кольцам деревьев).
Дюбуа (Мари) Эжен (Франсуа Тома) |Dubois, (Marie) Eugene
(Francois Thomas)] (1858- 1940). Голландский анатом; в 1891 г.
на острове Ява сделал первую находку остатков Homo erectus,
которого он назвал Pithecanthropus erectus.
Фиггинс Джесси Д. (Figgins, Jesse D.). Американский палеон-
толог; во время раскопок около Фолсома (штат Нью-Мексико)
в 1927 г. нашел каменный наконечник, застрявший между реб-
рами доисторического мамонта, что явилось первым убедитель-
ным доказательством древности пребывания человека на Аме-
риканском континенте.
Фрер Джон (1-гге. John) (1740--1807), Английский антиквар,
основатель доисторической археологии; в 1790 г. в Хоксне на
территории Восточной Англии (графства Суффолк и Норфолк.
Прим. пер.) нашел кремневые орудия и остатки вымерших живот-
ных, которые залегали в одних и тех же слоях.
Геер Герард Якоб, барон де (Geer, Gerard Jakob, Baron de)
(1858-1943). Шведский геолог; разработал метод хрономет-
рического датирования, основанный на подсчете годичных лент —
варв (годичных слоев осадков, отложенных в озере за один
сезон).
Гудолл Джейн ван Лоик (Goodall, Jane van Lawick). Уроженка
Англии, этнолог; ее детальные исследования поведения шимпанзе
в заповеднике Гомбе Стрим на территории Танзании в 60—70-х
годах нашего века пролили свет на формирование поведения
человека.
Геккель Эрнст Генрих (Haeckel, Ernst Heinrich) (1834 — 1919).
Блестящий немецкий биолог-эволюционист; ввел термин "Pithe-
canthropus" («обезьяночеловек») для обозначения предполагае-
мого недостающего промежуточного звена между человекооб-
разными обезьянами и человеком.
Джефферсон Томас (Jefferson, Thomas) (1743-1826). Третий
президент США; предвосхитил археологические исследования
Северной Америки, проводя раскопки древних могильных хол-
мов в штате Виргиния и выполняя стратиграфические наблюдения.
Йохансон Дональд (Johanson, Donald). Американский антро-
полог; в 1974 г. в Эфиопии нашел древнейший из известных нам
скелетов австралопитека («Люси»), а в 1978 г. назвал этот вид
Australopithecus afarensis.
Кинг Уильям (King, William). Ирландский анатом; в 1864 г.
ввел в обиход научный термин Homo neanderthalensis.
![]() 16 *
16 *
![]()
![]() Дональд Йохансон. нашедший
Дональд Йохансон. нашедший
«Люси» - скелет самки
австралопитеке.
243
Льюис Лики измеряет череп
австралопитека Бойса.
В поисках древнего человека (2)
Кёнигсвальд Густав Генрих Ральф фон (Koenigswald, Gustav
Heinrich Ralph von). Немецкий антрополог; в 1935 г. ввел в науку
термин Gigantopithecus, а также нашел ископаемые остатки
Homo erectus и часть массивной нижней челюсти Meganthropus
palaeojavanicus на острове Ява (1937—1941 гг.).
Ларте Эдуард Арман Изидор Ипполит (Lartet, Edouard Armand
Isidore Hippolyte) (1801 —1871). Французский геолог, археолог
и палеонтолог; ввел термины Dryopithecus и Pliopithecus. прово-
дил раскопки многих пещерных стоянок эпохи палеолита в Дор-
дони.
Ларте Луи (Lartet, Louis). Французский археолог и геолог, сын
Эдуарда Ларте (си.); в 1868 г. при раскопках в гроте Кро-Маньон
(департамент Дордонь) открыл так называемого кроманьонского
человека.
Лики Джонатан (Leakey, Jonathan). Старший сын Льюиса Лики
(см.)\ в 1960 г. в Олдувайском ущелье (Танзания) открыл пер-
вые ископаемые остатки, отнесенные к виду Homo habilis.
Лики Льюис Сеймур Бэзетт (Leakey, Louis Seymeur Bazett)
(1903—1972). Английский антрополог; в .ходе своих экспедиций
в Восточной Африке доказал, что эволюция человека началась
в Африке.
Лики Мэри (Leakey, Mary). Антрополог, родилась в Англии
(1913 г.), жена Льюиса Лики; открыла остатки Australopithecus
boisei (1959 г.), позднее руководила раскопками в Олдувайском
ущелье в Танзании и работами экспедиции, которая в 1977 г. на-
шла древнейшие из известных нам следов ног человека в Лаэтоли
(Танзания).
Лики Ричард Е. (Leakey. Richard E.). Директор Национального
музея Кении, сын Льюиса Лики (см. выше); в 1967 г. руководимая
им экспедиция в районе Омо (Эфиопия) обнаружила, возможно,
древнейший из известных нам черепов вполне современного чело-
века; начиная с 1968 г. при экспедиционных работах в окрестностях
озера Туркана (Рудольф) им было найдено много новых остатков
ранних гоминин.
Льюис Дж. Эдвард (Lewis, G. Edward). Ученый; в 1932 г. первым
определил род Ramapithecus («обезьяна Рамы»), ввел в науку
этот термин и выдвинул ныне оспариваемую гипотезу о том, что это
был предок человека.
Либби Уиллард Франк (Libby, Willard Frank) (1908—1980).
Американский химик; в 1947 г. разработал радиоуглеродный
метод датирования.
Линней Карл (Linnaeus, Carolus) (1707—1778). Шведский бота-
ник; разработал современную систему классификации живых орга-
низмов и в 1758 г. дал научное название нашему виду — Homo
sapiens.
Леббок, сэр Джон (Lubbock, Sir John) (1834—1913). Английский
археолог-любитель; ввел в науку термины «палеолит» (древний
каменный век) и «неолит» (новый каменный век).
Лайелль, сэр Чарльз (Lyell, Sir Charles) (1797—1875). Англий-
ский геолог; помог заложить основы современной геологической
науки.
Маршалл, сэр Джон Хьюберт (Marshall, Sir John Hubert) (1876—
1958). Английский археолог; его раскопки в Хараппе (1921 г.) и
Мохенджодаро (1922 г.), находящихся сейчас на территории
Пакистана, привели к открытию доисторической цивилизации
долины реки Инд, процветавшей в период 2500—1750 лет до н. э.
Марстон А. Т. (Marston, А. Т.). Английский археолог-любитель;
в 1935 г. нашел остатки человека из Сванскомба близ Лондона.
Меллаарт Джеймс (Mellaart, James). Английский археолог; его
раскопки в Чатал-Хююк (Турция) в 1961 —1965 гг. привели к от-
крытию крупнейшего из известных нам неолитических городов на
Ближнем Востоке.
![]()
![]()
![]()
![]() Мендель Грегор Иоганн (Mendel, Gregor Johann) (1822—1884).
Мендель Грегор Иоганн (Mendel, Gregor Johann) (1822—1884).
Австрийский монах и ботаник; открыл законы наследственности
и заложил тем самым основы генетики, которые объясняют био-
логические механизмы эволюции.
Морган Льюис Генри (Morgan, Lewis Henry) (1818 1881). Аме-
риканец, основатель научной антропологии; его работа «Древнее
общество» (1877 г.) явилась первой значительной научной попыт-
кой проследить развитие культуры «от Дикости через Варварство
к Цивилизации».
Питри, сэр Уильям Мэттью Флиндерс (Petne, Sir William Matthew
Flinders) (1853 1942) Английский археолог-египтолог; повысил
научный уровень проведения раскопок, особо подчеркнув важ-
ность внимания к деталям и ценность тщательней публикации ре-
зультатов раскопок.
Пнтт-Риверс Огастас Генрн ( Pitt-Rivers, Augustus Henry) (1827—
1900). Английский археолог; впервые ввел в практику ряд совре-
менных научных методов проведения археологических раскопок.
Шаафгаузен Германн (Schaaffhausen, Hermann). Немецкий an
трополог; в 1856 г. открыл остатки неандертальского человека
и опубликовал отчет об этом.
Шётензак Отто (Schoelensack, Otto) Немецкий профессор; в
1908 г. опубликовал детальное сообщение о нижней челюсти из
Мауэра — челюсти гейдельбергского человека.
Швальбе Густав (Schwalbe, Gustave) (1844 1917). Немецкий
анатом; приводил доводы в защиту того, что Pithecanthropus (Ho-
mo erectus) является предком современного человека.
Семенов С. A. (Semenov, S. А). Русский археолог; в его работе
«Доисторическая техника» (1957 г.) выдвинут принцип экспери-
ментального исследования микронзноса как ключа к пониманию
орудий эпохи каменного века.
Саймоне Элвин Л. (Simons, Elwyn L.). Американский палеонтолог;
в 60-х годах нашего века совершил важные находки ископаемых
остатков древних приматов в Фаюмской депрессии (Египет).
Соленки Ральф (Solecki, Ralph). Американский ученый; его рас-
копки, проведенные в 1953 I960 гг. в пещере Шанидар (Ирак),
пролили новый свет на образ жизни неандертальцев.
Стивене Джон Ллойд (Stephens, John Lloyd) (1805-1852).
Американский археолог; ere исследования руин построек .майя
в 1839-1842 гг. положили начало археологии Центральной Аме-
рики.
Сузуки Хисаши (Susuki, Hisashi). Японский археолог; в 60 х го-
дах нашего века провел раскопки пещеры Амуд в Израиле; най-
денные там костные остатки и орудия явились переходным звеном
между неандертальцем и современным человеком, между мустьер-
ской и позднепалеолитической культурами.
Томсен Кристиан Юргенсон (Themsen, Christian Jurgensen)
(1788-1865). Датский археолог; впервые разделил доисториче-
ский период на каменный, бронзовый и железный века.
Вейденрейх Франц (Weidenreich, Franz) (1873-1948). Уроже
нец Германии, анатом и археолог; известен своей реконструкцией
скелета синантропа («пекинского человека»), которая пережила
гибель остатков оригинала в 1941 г.; ввел в науку термин Homo
erectus (1940 г.).
![]()
![]()
![]() Уилсон Алан К. и Сэрич Винсент М. (Wilson, Alan С. and Sarich,
Уилсон Алан К. и Сэрич Винсент М. (Wilson, Alan С. and Sarich,
Vincent M.). Американские специалисты по молекулярной биоло-
гии; их «молекулярные часы», разработанные в 1969 г., показали,
что линии человекообразных обезьян и человека разошлись позд-
нее, чем это считалось на основе находок ископаемых остатков.
Вулли, сэр Чарльз Леонард (Woolley, Sir Charles Leonard)
(1880-1960). Английский археолог; его раскопки шумерского
города Ур (1922-1934 гг.) помогли понять историю древней циви-
лизации Месопотамии.
![]() Костяной скребок для очистки
Костяной скребок для очистки
кожи от мяса (Национальный
музей Человека в Оттаве).
|
Череп человека из Люцзяня |
В тысячах музеев экспонируются или хранятся орудия и костные
остатки доисторического человека, которые были найдены во время
раскопок бесчисленных погребений и прочих доисторических па-
мятников во всех уголках мира. В некоторых западных странах
главный город каждой области или штата имеет экспозицию, по-
священную жизни доисторических людей. На следующих шести
страницах дан краткий обзор основных музейных коллекций мира. J
(Страны и города приводятся в порядке латинского алфавита.—
Прим. пер.).
АВСТРИЯ (AUSTRIA)
Вена: Музей естественной истории (Vienna: Natural History Mu-
seum). В отделе доисторической жизни сосредоточен материал из
музеев бывшей Австро-Венгерской империи: предметы материаль-
ной культуры каменного века, орудия и оружие бронзового века,
оборудование для плавки металлов и орудия железного века.
БЕЛЬГИЯ (BELGIUM)
Брюссель: Королевский музей искусств и истории (Brussels: Royal
Museum of Art and History). Собранные со всего мира коллекции
состоят из археологических находок доисторического периода.
БОЛГАРИЯ (BULGARIA)
София: Музей истории Софии (Sofia: Museum of Sofia's History).
Местные находки начиная с эпохи неолита.
КАНАДА (CANADA)
Оттава: Национальный музей Человека (Ottawa: National Mu-
seum of Man). В числе древностей имеются предметы из Северной
Америки.
КИТАИ (CHINA)
Пекин: Институт археологии Китайской академии наук [Beijing
(Peking): Institute of Archeology, Academia SinicaJ. В коллекции
особое внимание уделяется доисторическому периоду китайской
истории.
Пекин: Институт палеонтологии позвоночных и палеоантропо-
логии (Beijing (Peking): Institute of Vertebrate Paleontology and
Paleoanthropology]. К известным китайским экспонатам относится
череп человека из Люцзяна.
КОЛУМБИЯ (COLOMBIA)
Ьогота: Национальный музей (Bogota: National Museum). Име-.
ются доисторические ископаемые остатки.
КИПР (CYPRUS)
![]()
Никозия: Археологический музей (Nicosia: Archeological Mu-
seum). Экспонаты иллюстрируют доисторический период истории
острова.
Каир: Египетский национальный музей (Cairo: Egyptian Natio-
nal Museum). Охватывает период от доисторических времен до
VI века н. э.
![]()
ЭФИОПИЯ (ETHIOPIA)
Аддис-Абеба: Национальный музей Эфиопии (Addis Ababa: Na-
tional Museum of Ethiopia). Имеются представляющие научную
ценность ископаемые остатки гоминид, в том числе вида Austra-
lopithecus afarensis.
ФРАНЦИЯ (FRANCE)
Бордо: Музей Аквитании (Bordeaux: Museum of Aquitaine). Кол-
лекции из районов Борделэ и Перигора.
Карнак: Музей доисторического периода (Carnac, Morbihan:
Museum of Prehistory). Коллекция доисторических экспонатов
(керамические изделия, бусы и т. д.), главным образом из по-
гребальных курганов (но наиболее значительными местными до-
историческими памятниками являются длинные ряды огромных,
стоящих вертикально камней).
Лез-Эйзи, Дордонь: Национальный музей доисторического перио-
да (Les Eyzies, Dordogne: National Museum of Prehistory). Уде-
лено особое внимание находкам, сделанным во время раскопок
местных пещер эпохи позднего палеолита.
Монтиньяк-Ляско, Дордонь: Пещера Ляско (Montignac-Lascaux,
Dordogne: Lascaux Cave). Великолепные доисторические наскаль-
ные рисунки; в настоящее время пещера для осмотра закрыта.
Нанси: Музей наук о Земле (Nancy: Museum of Earth Sciences).
Экспозиция прослеживает эволюцию жизни от древнейших орга-
низмов до человека.
Ницца: Музей Терра Амата (Nice: Museum of Terra Amata). Ис-
копаемые остатки, найденные на местной стоянке, возраст которых,
возможно, 400 тысяч лет.
Париж: Лувр (Paris: Louvre Museum). В его коллекциях име-
ются важные экспонаты древнейших европейских цивилизаций.
Париж: Музей Человека (Paris: Museum of Man). Имеются иско-
паемые остатки и предметы материальной культуры неандерталь-
цев и кроманьонцев.
Перигё, Дордонь: Перигорский музей (Perigueux; Dordogne: Рё
rigord Museum). Среди его экспонатов имеются скелет неандер-
тальца и фриз с изображением бизонов эпохи позднего палеолита.
Сен-Жермен-ан-Лэ, Ивелин: Музей национальных древностей
(Saint-Germaine-en-Laye, Yvelines: Museum of National Antiqui-
ties). Некоторые замечательные каменные и костяные орудия и
предметы искусства эпохи позднего палеолита из Франции.
Талане, Жиронда: Институт четвертичного периода, Университет
города Бордо (faience, Gironde: Institute of Quaternary, Borde-
aux University). Костные и каменные орудия эпохи позднего палео-
лита.
|
|

![]() «Венера» из Лосселя (музей
«Венера» из Лосселя (музей
в Бордо).
Череп неандертальца (Музей
Человека в Париже).
ГЕРМАНИЯ (GERMANY)
Берлин: Берлинский Государственный музей Ближнего Восто-
ка (Berlin: Berlin State Museum of Near Asia). Среди ближне-
восточных древностей имеются шумерские печати.
Берлин: Египетский музей (Berlin: Egyptian Museum). Особое
внимание уделено древнеегипетским скульптурам, печатям, ору-
диям, маскам и т. д.
Берлин: Музей доисторического периода и древней истории (Ber-
lin: Museum of Prehistory and Early History). Экспозиция посвя-
щена главным образом доисторическому периоду Европы и Азии
Бонн: Музей земли Рейнланд-Пфальц (Bonn: Rhineland Mu-
seum). Местные экспонаты доисторического периода и череп не-
андертальца.
Франкфурт: Музей доисторического периода и древней истории
![]()
![]() (Frankfurt: Museum of Prehistory and Early History). В основ-
(Frankfurt: Museum of Prehistory and Early History). В основ-
ном посвящен району Франкфурта.
Гамбург: Музей этнографии и доисторического периода (Ham-
burg: Museum of Ethnology and Prehistory). Коллекции со всего
мира.
Штутгарт: Музей земли Баден-Вюртемберг (Stuttgart: Wiirttem-
berg Museum). Посвящен доисторическому периоду, древней ис-
тории и т. д.
ГРЕЦИЯ (GREECE)
Афины: Музей антропологии и этнографии (Athens: Anthropo-
logical and Ethnological Museum). Коллекции со всего мира содер
жат предметы, относящиеся к доисторическому периоду.
Салоники: Музей палеонтологии, Университет города Салоники
(Thessaloniki: Paleontological Museum, University of Thessalo-
niki). В нем находится знаменитый череп из Петралоны (Homo
erectus или архаичная разновидность Homo sapiens).
ГВАТЕМАЛА (GUATEMALA)
Город Гватемала: Археологический музей (Gautemala City: Arche-
ological Museum). Предметы культуры майя и другие экспонаты.
ГОНДУРАС (HONDURAS)
Тегусигальпа: Музей истории и археологии (Tegucigalpa: Museum
of History and Archeology). Экспонированы археологические
находки из местных раскопок.
|
|
![]()
![]() Череп из Шанидара-1
Череп из Шанидара-1
(Иракский музей, Багдад).
Глиняная статуэтка культуры
дзёмон (Национальный музей
Токио).
ВЕНГРИЯ (HUNGARY)
Будапешт: Венгерский национальный музей (Budapest: Hunga-
rian National Museum). Представлены палеолит, неолит, бронзо-
вый и железный века.
ИНДИЯ (INDIA)
Калькутта: Индийский музей (Calcutta: Indian Museum). Древно-
сти доисторического и более поздних периодов.
ИНДОНЕЗИЯ (INDONESIA)
Чианджар: Музей ГедонгАртья (Gianjar: Museum Gedong Artja).
Орудия палеолита, неолита и других доисторических периодов.
ИРАН (IRAN)
Тегеран: Археологический музей (Tehran: Archeological Museum).
Древности эпохи палеолита и более поздних периодов.
ИРАК (IRAQ)
Багдад: Иракский музей (Baghdad: Iraq Museum). Имеются
остатки неандертальцев из Шанидара, неолитическая керамика
и предметы, относящиеся к ранним цивилизациям Месопота-
мии.
ИРЛА НДИ Я (IRELAND)
Дублин: Национальный музей (Dublin: National Museum). Ору-
дия эпохи мезолита и неолита, а также многочисленные золотые
украшения доисторического периода.
ИЗРАИЛЬ (ISRAEL)
Хайфа: Музей естественной истории и доисторического периода
(Haifa: Museum of Natural History and Prehistory).
Иерусалим: Музей доисторического периода (Jerusalem: Museum
of Prehistory). Предметы с доисторических стоянок в Израиле.
ИТАЛИЯ (ITALY)
Милан: Археологический музей (Milan: Archeological Museum).
Неолитические и другие доисторические экспонаты.
Рим: Коллекция Итальянского института палеоантропологии
(Rome: Collections of the Italian Institute of Human Paleonto-
logy).
Рим: Музей антропологии, Римский университет (Rome: Museum
of Anthropology,-the University). Имеются скелеты и черепа, в
том числе неандертальцев.
ЯПОНИЯ (JAPAN)
Токио: Национальный музей Токио (Tokyo: Tokyo National Mu-
seum). Имеются японские находки доисторического периода.
ИОРДАНИЯ (JORDAN)
Амман: Иорданский археологический музей (Amman: Jordan
Archeological Museum). Экспозиции охватывают все периоды от
позднего палеолита до средневековья.
КЕНИЯ (KENYA)
Найроби: Национальный музей Кении (Nairobi: National Museum
of Kenya). Некоторые значительные находки ископаемых при
матов, включая обезьяночеловека Proconsul, австралопитеков,
Homo habilis и Homo erectus.
МЕКСИКА (MEXICO)
Мерида: Юкатанский археологический музей (Merida: Yucatan
Museum of Anthropology). Представлены доиспанские культуры
Мексики.
Мехико: Национальный археологический музей (Mexico City:
National Anthropological Museum). Экспонаты отражают исто-
рию Мексики начиная с доисторических времен.
Череп Homo erectus
(Национальный музей Кении,
Найроби).
НИДЕРЛАНДЫ (NETHERLANDS)
Амстердам: Археологический музей Амстердамского университета
(Amsterdam: Archeological Museum of the University of Am-
sterdam). Экспонируются вазы, изделия из бронзы и другие на-
ходки с Ближнего Востока и из Египта.
Лейден: Национальный музей древностей (Leiden: National
Museum of Antiquities). Предметы с Ближнего Востока и из Егип-
та.
Лейден: Национальный этнологический музей (Leiden: National
Museum of Ethnology). Доисторические экспонаты из Индонезии и
Америки доколумбова периода.
ПАКИСТАН (PAKISTAN)
Карачи: Национальный музей (Karachi: National Museum). Экс-
понируются орудия эпохи палеолита из долины реки Соан и памят-
ники цивилизации долины реки Инд.
ПАНАМА (PANAMA)
Город Панама: Музей панамского человека (Panama City: Mu-
seum of Panama Man). Посвящен археологии и этнографии.
ПЕРУ (PERU)
Лима: Музей искусств (Lima: Museum ofArt). Отражает культуру
доколумбова периода.
ПОЛЬША (POLAND)
Варшава: Национальный археологический музей (Warsaw:
National Museum of Archeology). Посвящен археологии Польши
доисторического и более поздних периодов.
РУМЫНИЯ (ROMANIA)
Бухарест: Национальный музей древностей (Bucharest: National
Museum of Antiquities). Памятники отражают каменный и брон-
зовый века доисторической Румынии.
ЮЖНАЯ АФРИКА (ЮАР) (SOUTH AFRICA)
Претория: Музей Трансвааля (Pretoria: Transvaal Museum).
Среди ископаемых имеются остатки видов Australopithecus afri-
canus и robustus.
ИСПАНИЯ (SPAIN)
Мадрид: Национальный археологический музей (Madrid: Natio-
nal Archeological Museum). Доисторические находки из Амброны.
Сантильяна-дель-Мар, Сантандер: Пещера Альтамира (Santillana
del Mar, Santander: Altamira Cave). Выдающееся пещерное
искусство позднего палеолита.
ШВЕЦИЯ (SWEDEN)
Стокгольм: Исторический музей (Stockholm, Historical Museum).
Доисторические экспонаты из Швеции.
Стокгольм: Музей древностей Средиземноморья и Ближнего Вос-
тока (Stockholm: Museum of Mediterranean and Near Eastern
Antiquities). Одна из экспозиций посвящена доисторической Азии.
ШВЕЙЦАРИЯ (SWITZERLAND)
Берн: Бернский исторический музей (Bern: Bern Historical Mu-
seum). Главным образом доисторический материал начиная с
эпохи палеолита.
Цюрих: Швейцарский национальный музей (Zurich: Swiss Natio-
nal Museum). Посвящен доисторической Швейцарии.
СИРИЯ (SYRIA)
Дамаск: Дамасский национальный музей (Damascus: Damascus
National Museum). Археологические экспонаты представляют
древние цивилизации страны.
ТАНЗАНИЯ (TANZANIA)
Дар-эс-Салам: Национальный музей Танзании (Dar es Salaam:
National Museum of Tanzania.). Среди экспонатов — первый из
найденных черепов Australopithecus boisei.
ТУРЦИЯ (TURKEY)
Анкара: Музей цивилизации Анатолии (Ankara: Museum of Anato-
lian Civilizations). Экспонаты эпох палеолита, неолита, хальколи-
та (медного века), бронзового века и более поздних периодов.
Стамбул: Археологический музей Стамбула (Istanbul: Archeolo-
gical Museum of Istanbul). Имеются шумерский, аккадский, хет-
тский, ассирийский, египетский и другие материалы ранних ци-
вилизаций.
|
|
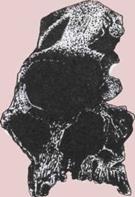
![]() Череп австралопитека
Череп австралопитека
африканского (Australopithecus
ajricanus) (Претория).
|
|

Ручное рубило (Мадрид).
Череп австралопитека Бойса
(Australopithecus boisei)
(Дар-эс-Салам).
ВЕЛИКОБРИТАНИЯ (UNITED KINGDOM)
Эйвбери, Уилтшир: Музей города Эйвбери (Avebury, Wiltshire:
Avebury Museum). Местную поздненеолитические находки (Эй-
вбери окружен доисторическими земляными сооружениями).
Бирмингем: Городской музей (Birmingham: City Museum). Име-
ются доисторические и египетские древности.
Бристоль: Городской музей (Bristol: City Museum). Среди кол-
лекций, собранных со всего мира, имеются экспонаты, относя-
щиеся к древней истории.
Кембридж: Музей археологии и антропологии (Cambridge: Mu-
seum of Archaeology and Anthropology). Экспонируются архео-
логические находки со всего мира начиная с каменного века.
Кардифф: Национальный музей Уэльса (Cardiff: National Mu-
seum of Wales.) Имеется археологическая коллекция.
Чеддар, Сомерсет: Чеддарский пещерный музей (Cheddar, So-
merset: Cheddar Caves Museum). Содержит находки эпохи палео-
лита и более поздних периодов.
Донкастер, Южный Йоркшир: Музей (Doncaster, South York-
shire: Museum). Среди экспонатов предметы доримского периода.
Дорчестер, Дорсет: Музей графства Дорсет (Dorchester, Dorset:
Dorset County Museum). Превосходная коллекция доисториче-
ских орудий.
![]() Деревянный наконечник копья
Деревянный наконечник копья
эпохи палеолита (Британский
музей, Отделение
естествфзнания).
|
|

Один из первых известных нам
письменных документов
табличка из города Киш
в Ираке; ее возраст 5500 лет
(Ашмольский музей, Оксфорд).
Череп «египтопитека»
(Propiiopithecus) (Музей
естествознания имени Пибоди,
Ныо-Хейвен).
Эдинбург: Национальный музей древностей Шотландии (Edin-
burgh: National Museum of Antiquities of Scotland). Обширные
коллекции доисторических экспонатов.
Глазго: Художественная галерея и музей Глазго (Glasgow:
Glasgow Art. Gallery and Museum). Имеются археологические
экспонаты.
Глазго: Хантеровский музей (Glasgow: Hunterian Museum). Ар-
хеологические экспонаты находятся в главном здании универ-
ситета.
Ипсвич, Суффолк: Музей (Ipswich, Suffolk: The Museum). До-
исторические находки из Восточной Англии.
Лондон: Британский музей (London: British Museum). Круп-
нейший музей с обширными археологическими коллекциями всех
периодов из многих стран.
Лондон: Британский музей (Отделение естествознания) (Lon-
don: British Museum (Natural History)]. Крупнейший музей;
содержит экспонаты, прослеживающие место человека в про-
цессе эволюции; имеются некоторые важнейшие ископаемые остат-
ки человека.
Лондон: Музей человечества (London: Museum of Mankind). Глав-
ным образом этнографический музей с археологическими наход-
ками из Северной и Южной Америки.
Манчестер: Манчестерский музей (Manchester: Manchester Mu-
seum). Среди археологических экспонатов много предметов из
Древнего Египта.
Ньюкасл-апон-Тайн: Музей древностей (Newcastle upon Tyne:
Museum of Antiquities). В коллекции имеются модели и рекон-
струкции.
Оксфорд: Ашмольский музей изящных искусств и археологии
(Oxford: Ashmolean Museum of Art and Archeology). Старей-
ший музей Великобритании с археологическими экспонатами из
Европы и Ближнего Востока.
Плимут: Городской музей (Plymouth: City Museum). Имеются
археологические экспонаты.
Солсбери, Уилтшир: Музей Солсбери и Южного Уилтшира (Salis-
bury, Wiltshire: Salisbury and South Wiltshire Museum). Име-
ются важные доисторические находки из Стоунхенджа.
Торки, Девоншир: Музей общества естествознания (Torquay,
Devon: Natural History Society Museum). Доисторические на-
ходки из Кентской пещеры.
США (UNITED STATES)
Остин, Техас: Техасский мемориальный музей (Austin, Texas:
Texas Memorial Museum). Экспозиции отражают эволюцию че-
ловека и культуру американских индейцев.
Беркли, Калифорния: Антропологический музей им. Роберта
Г. Лоуи (Berkeley, California: Robert H. Lewie Museum of Anth-
ropology). Антропологические и археологические экспонаты с раз-
личных континентов.
Чикаго, Иллинойс: Полевой музей естествознания (Chicago,
Illinois: Field Museum of Natural History). Археологические и
антропологические экспонаты.
Цинциннати, Огайо: Музей естествознания города Цинциннати
(Cincinnati, Ohio: Cincinnati Museum of Natural History). Среди
археологического материала имеются памятники культуры аме-
риканских индейцев.
Эль-Пасо, Техас: Музей парка Уилдернесс (El Paso, Texas:
Wilderness Park Museum). Отражены археология и культура
юго-запада США и севера Мексики.
Канзас-Сити: Историко-научный музей Канзас-Сити (Kansas
City: Kansas City Museum of History and Science). Посвящен
истории и археологии индейцев.
Лос-Анджелес: Музей изящных искусств округа Лос-Анджелес
(Los Angeles: Los Angeles County Museum of Art). Имеется ар-
хеологический материал.
Мадисон, Висконсин: Музей исторического общества штата Вис-
консин им. Вашингтона (Madison, Wisconsin: Museum of the
Stale Historical Society of Washington). Охватывает историю шта-
та Висконсин с доисторических времен.
Национальный парк Меса-Верде, Колорадо (Mesa Verde National
Park, Colorado). Имеются сотни руин индейских поселений древ-
ностью до 1300 лет.
Майами, Флорида: Исторический музей Южной Флориды (Miami,
Florida: Historical Museum of Southern Florida). Посвящен
местной антропологии, археологии и т. д.
Нашвилл, Теннесси: Музей штата Теннесси (Nashville, Tennessee:
Tennessee State Museum). Представлены предметы индейской
культуры и местной истории.
Нью-Хейвен, Коннектикут: Музей естествознания им. Пибоди,
Йельский университет (New Haven, Connecticut: Peabody Mu-
seum of Natural History, Yale University). В числе ископаемых
остатков приматов имеется "Aegyptopithecus".
Ныо-Орлеан, Луизиана: Музей штата Луизиана (New Orleans,
Louisiana: Louisiana State Museum). Охватывает местную исто-
рию и_археологию, представленную индейскими орудиями.
Нью-Йорк, штат Нью-Йорк: (Фонд Гейе) Музей американских
индейцев [New York City, New York: (Heye Foundation) Museum
of the American Indian]. Содержит богатую коллекцию тради-
ционных индейских орудий.
Норфолк, Виргиния: Крайслеровский музей (Norfolk. Virginia:
Chrysler Museum). История и археология Норфолка.
Оклахома-Сити, Оклахома: Музей Историческою общества (Ok-
lahoma City, Oklahoma: Historical Society Museum). Посвящен
истории штата с доисторических времен.
Питтсбург, Пенсильвания: Музей естествознания им. Карнеги
(Pittsburgh, Pennsylvania: Carnegie Museum of Natural History).
Имеются антропологические экспонаты.
Портленд, Орегон: Музей естествознания (Portland, Oregon:
Museum of Natural History). Материал по археологии и антропо-
логии индейцев.
Ричмонд, Виргиния: Музей Археологического общества штата
Виргиния (Richmond, Virginia: Museum of the Archeological
Society of Virginia). Особое внимание уделяется археологии этого
штата.
Рочестер, Нью-Йорк: Музей Археологического общества штата
Нью-Йорк (Rochester, New York: Museum of the New York State
Archeological Association). Посвящен археологии и антрополо-
гии северо-востока Северной Америки.
Сакраменто, Калифорния. Индейский музей штата Калифорния
(Sacramento, California: State Indian Museum). Отражены жизнь
и предметы материальной культуры индейцев Калифорнии.
Сент-Луис, Миссури: Музей науки и естествознания (St. Louis,
Missouri: Museum of Science and Natural History). Материалы
по антропологии и археологии индейцев штата Миссури.
Сан-Диего, Калифорния: Музей Человека города Сан-Диего (San
Diego, California: San Diego Museum of Man). Имеются черепа
древних индейцев.
|
|

Золотая корона из Чавина
в Перу (Музей американских
индейцев, Нью-Йорк).
|
Череп человека из Дель-Мар |
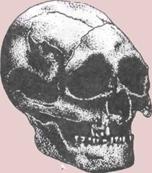
|
|
| Человеческие фигурки, |
Сан-Франциско, Калифорния: Музей восточного искусства (San
Francisco: Asian Art Museum). Предметы искусства и культуры
Азии начиная с неолита.
Сиэтл, Вашингтон: Музей изящных искусств города Сиэтл (Seatt-
le, Washington: Seattle Art Museum). Предметы искусства Древ-
него Египта и Америки доколумбова периода.
Спрингфилд, Иллинойс: Музей штата Иллинойс (Springfield,
Illinois: Illinois State Museum). Имеется материал по индейской
этнографии.
Тусон, Аризона: Музей штата Аризона (Tucson, Arizona: Arizona
State Museum). Представлен индейский материал, особенно куль-
тура гогокам.
Вашингтон, округ Колумбия: Национальный музей естествозна-
ния, Смитсоновский институт (Washington, D. С: National Mu-
seum of Natural History, Smithsonian Institution). Крупный му-
зей; имеются доисторические экспонаты; особое внимание уде-
лено американским индейцам и их образу жизни.
СССР (USSR)
Ленинград: Ленинградское отделение Института археологии
Академии наук СССР (Leningrad: Institute of Archeology). В числе
экспонатов имеется костяной наконечник, застрявший в лопатке
зубра.
Москва: Музей антропологии им. Д. Н. Анучина (Moscow:
D. N. Anuchin Anthropology Museum). Посвящен антропологии
и археологии доисторического периода.
Общие вопросы.
Aiello L. Discovering the Origins of Mankind. Longman, 1982.
Brace С. E. and Montagu A. Human Evolution. Macmillan, 1977.
Clapham Frances M. (editor). The Rise of Man. Sampson Low, 1976.
Coles I. M. and Higgs E. S. The Archaeology of Early Man. Faber and Faber, 1969.
Junnain R., Nelson H., Kurashina H. and Turnbaugh W. A. Understanding Physical Anthropology
Archaeology. West Publishing Co., 1981.
Kennedy G. E. Paleoanthropology. McGraw Hill, 1980.
Leakey R. E. The Making of Mankind. Michael Joseph, 1981.
Lewin R. Human Evolution. Blackwell Scientific Publications, 1984.
Oakley K. P. Man the Tool-Maker. British Museum (Natural History), 1972.
Poirier F. E. Fossil Evidence. С V. Mosby Co., 1981.
Tomkins S. The Origins of Mankind. Cambridge University Press, 1984.
Wood B. The Evolution of Early Man. Peter Lowe, 1976.
Примеч a н и е. Сведения о новейших крупных открытиях публикуются в периодической печати и в
научных журналах, например в "Nature" и "Scientific American".
Глава 1 Что такое человек?
Gribbin J. and Che fas J. The Monkey Puzzle. Bodley Head, 1982.
Romer A. S. Man and the Vertebrates (2 vols). Penguin Books, 1954.
Wood P., Vaczek L, Hambtin D. J. and Leonard J. N. Life Before Man. Time Inc, 1972.
Главы 2—3. Примитивные приматы. Эволюция антропоидов.
Colbert E. H. Evolution of the Vertebrates. Wiley, 1980.
Napier P. Monkeys and Apes. Hamlyn, 1970.
Szalay F. S. and Delson E. Evolutionary History of the Primates. Academic Press, 1979.
Глава 4. Человекообразные обезьяны и человек.
Goodall J. In the Shadow of Man. Houghton Mifflin, 1971.
Gribbin J. and Cherjas J. См. выше.
Смотри также литературу к главам 2 и 3.
Глава 5. «Человекообезьяны» и ранние люди.
Edey M. A. The Missing Link. Time Inc., 1972.
Leakey M. Olduvai Gorge. Collins, 1981.
Leakey R. E. См. выше.
Reader ]. Missing Links. Collins, 1981.
Szalay F. S. and. Delson E. См. выше.
Глава 6. Человек прямоходящий.
Leakey R. and Walker A. "Homo Erectus Unearthed". National Geographic, November, 1985.
White E. and Brown D. M. The First Men. Time Inc, 1972.
Глава 7 Неандертальский человек.
Constable G. The Neanderthals. Time Inc, 1972.
Shackley M. Neanderthal Man. Duckworth, 1980.
Главы 8 —10. Современный человек в Европе. Современный человек осваивает Землю. После леднико-
вого периода.
Ctaiborne Robert. The First Americans. Time Inc, 1973.
Cole S. The Neolithic Revolution. British .Museum (Natural History), 1970.
Hamblin D. J. The First Cities Time Inc, 1973.
Leonard J. N. The First Farmers. Time Inc, 1973.
McEvedy C. and Jones R. Atlas of World Population History. Allen Lane, 1978.
Phillips P. The Prehistory of Europe. Penguin Books, 1981.
Prideaux T. Cro-Magnon Man. Time Inc, 1973.
Глава 11. Познание истории человечества.
Binjord Lewis R. In Pursuit of the Past. Thames and Hudson, 1983.
Brothwell D. R. Digging up Bones. British Museum (Natural History) / Oxford University Press, 1981.
СПРАВОЧНОЕ ИЗДАНИЕ
ЛАМБЕРТ ДАВИД
Кембриджский путеводитель
Редактор издательства Л. Г. Ермолаева
Художественный редактор Н. А. Кравцова
Технический редактор Н. П. Старостина
Корректоры Н. Г. Клубкова, Е. А. Стерлина
ИБ 8422
Сдано в набор 19.07.90. Подписано в печать 25.06.91. Формат
70Х 1001
/16
. Бумага офсетная № 1. Гарнитура литературная. Пе-
чать офсетная. Усл. печ. л. 20,64. Усл. кр.-отт. 82,56. Уч.-изд. л.
25.10. Тираж 50 000 экз. Заказ № 5116/889. Цена 7 руб.
Ордена «Знак Почета» издательство «Недра», Ленинградское
отделение.
193171, Ленинград, С-171, ул. Фарфоровская, 18.
Ленинградская фабрика офсетной печати № 1 дважды ордена
Трудового Красного Знамени Ленинградского производственного
объединения «Типография имени Ивана Федорова» Государствен-
ного Комитета СССР по печати.
191126, Ленинград, ул. Звенигородская, д. 11.
%
7 руб.
Эта книга — краткое, но достаточно полное изложение
современных представлений о биологической эволюции
и ранних стадиях культурного развития человека. По свое-
му содержанию она дополняет кембриджский путево-
дитель «Доисторическая жизнь». Написанный в общедо-
ступной форме лаконичный текст сопровождается много-
численными рисунками, на которых представлены изо-
бражения древних приматов, реконструкции их черепов
и скелетов, эволюционные древа, карты и различные
диаграммы.
В книге рассматривается комплекс уникальных признаков,
присущих человеку, и подробно освещаются вопросы:
как, когда и где появился человек и какими были его обезь-
яноподобные предки.
Рассказано о палеонтологических и археологических ис-
следованиях, об образовании окаменелостей, о поисках
вымерших предшественников рода людского и о том, как
ученые обнаруживают ископаемые остатки древних орга-
низмов, определяют их возраст и воссоздают картины
жизни, шаг за шагом восстанавливая историю развития
человечества.
Для каждого, кто захочет расширить свои знания о проис-
хождении человека, эта книга станет источником ценной
информации и послужит введением в увлекательную об-
ласть палеонтологии и антропологии.
Пиша, территория и люди.
Этими тремя квадратами
изображена сравнительная
площадь территории, которая
нужна для пропитания трех
человек, добывающих себе пищу
разными способами.
1. Охотник-собиратель; 3,86
квадратной мили (10 км2
).
2. Земледелец (не
использующий полив); 0,19
квадратной мили (0,5 км2
).
3. Земледелец (пользующийся
ирригацией); 0,04 квадратной
мили (0,1 км2
).