| Скачать .docx | Скачать .pdf |
Курсовая работа: Экологическая обусловленность чувств животных
Экологическая обусловленность чувств животных
Введение
Рассматривая животный мир как целое, можно проследить основные направления эволюции сенсорных механизмов. Но эволюция происходит в результате естественного отбора, а естественный отбор - форма приспособления к данным условиям среды. Поэтому не следует удивляться тому, что многие различия между животными объясняются не столько их эволюционной историей, сколько экологическими условиями.
Сенсорные модальности наилучшим образом приспособлены к определенным местообитаниям, а отдельные модальности могут видоизменяться в соответствии с образом жизни животного. Это относится не только к связи между животным и окружающей его физической средой, но также и к его общению с другими животными. Эффективность такого общения зависит от соотношения между используемой модальностью и средой, через которую передается информация.
Зрительная модальность, определяя направленность света, позволяет точнее локализовать стимул и опознать конфигурацию пространства, чем любая другая модальность. Не случайно у хищников, например у осьминога, ястреба и кошки, зрительный аппарат развит сильнее, чем у других моллюсков, птиц и млекопитающих соответственно. Зрение позволяет также пользоваться сравнительно долговременными средствами коммуникации в том смысле, что животные могут особым образом изменять поверхность объектов или конструировать артефакты, надолго сохраняющие информацию. Главный недостаток зрения состоит в том, что оно работает хорошо только в определенных условиях. Зрительные сигналы в отличие от звуковых и обонятельных не могут обходить препятствия. В темноте или в сумерках зрение ограничено, хотя многие животные обладают специальными адаптациями, которые позволяют им до некоторой степени преодолевать этот недостаток.
У звука два особых свойства, делающих его важным орудием коммуникации. Во-первых, в звуковых сигналах можно производить быстрые временные изменения. Эта изменчивость позволяет быстро обмениваться информацией, что может быть важно для высокоподвижных видов животных. Кроме того, звуковые сигналы можно произвольно включать и выключать. Так, например, домовый сверчок зрительно хорошо замаскирован. Самец объявляет о своем присутствии продолжительным стрекотанием. Когда он чувствует опасность, он просто перестает стрекотать и замирает, из-за чего хищнику трудно его обнаружить. Второе важное свойство звука состоит в том, что его интенсивность может быть повышена над фоновым уровнем окружающей среды. А со зрительными сигналами это могут сделать лишь несколько видов, которые сами излучают свет. Громкая вокализация птиц и обезьян в тропическом лесу служит примером той пользы, которую приносит звук в преодолении препятствий и шумового фона.
Обоняние обладает некоторыми преимуществами и зрения, и слуха. Запахи могут служить долговременным сигналом и широко используются в этой функции наземными насекомыми и другими животными. Подобно звуковым сигналам, химические сообщения обходят препятствия, и интенсивность их можно поднять выше фонового уровня. Чувствительность обонятельных рецепторов, которая уже обсуждалась в случае бомбикола - феромона тутового шелкопряда, делает химические сигналы особенно пригодными для общения на очень больших расстояниях. Относительная стойкость таких сигналов иногда может быть недостатком, и беспечное оставление видами-жертвами химических следов используют многие хищники. Все три модальности имеют свои достоинства и свои недостатки, и многие животные используют каждую из них, смотря по обстоятельствам. Существует также много других, более специализированных видов чувствительности.
1. Зрительные приспособления к неблагоприятным условиям среды
Животное, глаза которого приспособлены к яркому свету, будет обладать хорошей остротой зрения, восприятием цвета и движения. Такие типично дневные глаза относительно нечувствительны к низким уровням освещенности. Животные, адаптированные к сумеречному освещению, обладают более высокой чувствительностью, но в ущерб цветовому и детальному предметному зрению.
Видеть при слабом освещении нужно животным, ведущим ночной образ жизни, обитающим глубоко в воде и в пещерах. Эти условия не совсем сравнимы, потому что спектр приходящего света смещен в воде. Нет данных о том, что ночные животные обладают повышенной чувствительностью к длинным волнам. Однако некоторые из них владеют приспособлениями, повышающими чувствительность к свету. Глаза с широким зрачком и большим хрусталиком улавливают больше света, чем маленькие глаза. Глазами первого типа обладают, например, опоссум, домовая мышь и рысь. У других ночных животных, например сов и галаго, череп сужен с боков, что привело к цилиндрическому удлинению глаза. Цилиндрические глаза встречаются также у некоторых глубоководных рыб. Глаз неясыти несколько чувствительнее человеческого, но этого недостаточно для успешной ночной охоты. Как мы увидим, совы полагаются также на другие органы чувств. Многие глубоководные животные специально адаптированы к господствующим условиям освещения. Максимумы поглощения у зрительных пигментов глубоководных рыб совпадают с длиной волны, максимально пропускаемой водой. У глубоко ныряющего кита северного плавуна зрительные пигменты сильнее всего поглощают более короткие волны, чем у неглубоко ныряющею серого кита. Точно так же глаза глубоководных морских ракообразных содержат зрительные пигменты с максимумами поглощения в более коротковолновой области, чем у мелководных ракообразных.
В дополнение к свойствам зрительных пигментов сумеречное зрение усиливают также другие специальные приспособления. Как правило, у ночных животных палочек больше, чем колбочек, а у некоторых из них, например у кошачьей акулы игалаго, мало или совсем нет колбочек. С одной биполярной клеткой может быть соединено много палочек, что повышает чувствительность к свету за счет остроты зрения. У многих ночных позвоночных близ фоторецепторов расположены отражающие свет образования, благодаря которым глаза "светятся", когда на них падает луч света. Свет, который проходит через фоторецептор не поглотившись, отражается обратно, что повышает вероятность поглощения. У некоторых рыб тапетум может быть покрыт мигрирующими пигментными зернами.
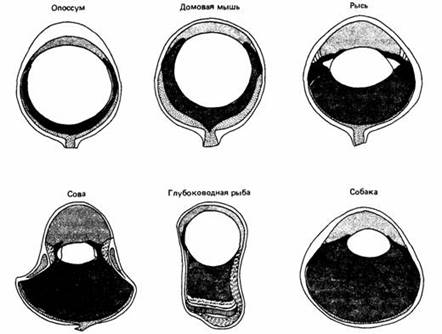
Рис.1. Сравнение глаз ночных животных и собаки, обладающей дневным и ночным зрением.
Например, у обыкновенной колючей акулы адаптированный к темноте тапетум отражает 88% падающего света, а после световой адаптации, когда отражение от тапетума заэкранировано, отражается только 2,5% падающего света. Как мы видим, животные, обитающие при очень слабом свете, приспособлены к нему по-разному. Некоторые из них живут в мутной среде, где зрительный контраст ослаблен из-за рассеяния света взвешенными частицами. В таких условиях возможности улучшения зрительного восприятия весьма ограничены.
Существует грань, за которой условия для зрения так тяжелы, что от глаз как от главного органа чувств приходится отказаться. У большинства животных, обитающих в пещерах, в глубине моря или же в мутной воде, глаза рудиментарные вследствие их регрессии. Например, пещерные земноводные обладают глазами на личиночной стадии, но лишены их во взрослом состоянии. Если этих животных воспитывать на свету, то у взрослых особей развиваются нормальные глаза. У молоди слепой пещерной рыбы имеются глаза, но у взрослых животных они дегенерируют. Садоглу производил генетические скрещивания этих рыб с живущим близ поверхности видом Astyanax с нормальным зрением. В этих исследованиях было обнаружено, что дегенеративное состояние глаз у пещерного вида определяется генами. Ошибневые рыбы обычно живут на больших глубинах, и глаза у них регрессировали. Некоторые виды вторично эволюционировали для жизни в пещерах.
Среди млекопитающих кроты и летучие мыши являются самыми известными примерами дегенерации зрения. Очень маленькие глазки кротов у некоторых видов покрыты кожей. У плотоядных рукокрылых зрение развито хорошо, но у ночных летучих мышей, особенно у тех видов, которые ловят насекомых на лету, зрение очень слабое. Очевидно, при добывании пищи они должны полагаться на другие органы чувств.
2. Органы чувств, заменяющие зрение
Животные с очень слабым зрением, живущие при сумеречном свете, должны рассчитывать на другие органы чувств. Так, у придонных кошачьих сомов и ошибневых имеются разные сенсорные усики, которыми они ощупывают субстрат. Эти усики снабжены многочисленными осязательными рецепторами и хеморецепторами. Однако такие органы чувств не могут заменить зрение как источник информации о величине и положении предметов в окружающей среде. Рыбы с нейромастами и органами боковой линии, чувствительными к вибрации, могут обнаруживать движущиеся предметы и получать некоторую информацию о неподвижных предметах по отраженным от них движениям воды. Но наилучшей заменой зрения животным служат электромагнитная чувствительность и особые слуховые приспособления.
Многие низшие животные способны ориентироваться в искусственных электрических полях, но мало что известно о сенсорной основе такого поведения. Некоторые виды рыб используют электрическую чувствительность при обычной ориентации и коммуникации, и ученым известно многое об их электросенсорных системах. Чувствительность к магнитным полям тоже обнаружена у ряда животных. Так, некоторые бактерии ориентируются к северному магнитному полюсу и реагируют на магнит в лабораторных условиях. С помощью электронного микроскопа у таких бактерий были обнаружены цепочечные структуры, содержащие кристаллы магнетита, которые также найдены в брюшке медоносных пчел и в сетчатке голубей. В Северном полушарии бактерии следуют наклонению магнитного поля Земли, и оно направляет их вниз, в анаэробный ил, их естественную среду обитания. В Южном полушарии у бактерий обратная полярность. Магнитоориентированное поведение изучалось также у пчел и голубей, а некоторые исследователи считают, что к магнитным полям чувствителен и человек.
Рыбы используют электричество тремя разными способами.1) Так называемые "сильноэлектрические" рыбы, такие, как электрический скат и электрический угорь, производят электрические разряды, способные оглушить жертву, но лишены электрической чувствительности.2) Электрочувствительные рыбы, например Scyliorhinus и некоторые другие акулы, не производят электричества. Однако кошачьи акулы способны обнаружить даже зарывшуюся в песок добычу по локальному искажению электрического поля Земли. Для этого служат особые органы чувств - так называемые ампулы Лоренцини, широко распространенные по всей поверхности тела, особенно вблизи головы.3) Так называемые "слабоэлектрические" рыбы генерируют свои электрические поля и чувствительны к электрическим изменениям среды. Обычно это ночные рыбы, обитающие в мутной воде, где зрение неприменимо. У них два типа электрочувствительных рецепторов: ампульные, которые отвечают на медленно изменяющиеся электрические поля, и клубневые, реагирующие только на быстрые их изменения. У некоторых видов один тип рецепторов, у других - оба типа. Эти рыбы генерируют слабые электрические поля посредством электрических органов, которые представляют собой видоизмененные мышцы или аксоны. Электрические разряды, как правило, испускаются с частотой до 300 импульсов в секунду. Некоторые рыбы способны менять частоту импульса, что используется для коммуникации с другими рыбами или как часть заглушающей реакции избегания, которая снижает действие полей, генерируемых другими особями вида. Иными словами, когда одна рыба испытывает электрическое воздействие другой, она может изменить частоту своих импульсов, чтобы снизить это вмешательство. Электрорецепторы служат также для локализации предметов в окружающей воде по искажениям, которые эти предметы вызывают в электрическом поле. Некоторые рыбы, например Gymnarchus, отличают хорошие проводники от плохих, например металлический стержень от пластикового. Более подробное описание механизмов электрорецепции у слабоэлектрических рыб можно найти у Эверта.
Слух заменяет зрение многим видам, и у некоторых из них образовались весьма интересные и специализированные добавления к нормальному слуху. Все эти адаптации способствуют точной локализации источника звука. Сравнение сигналов, приходящих в оба уха, служит главным способом локализации этого источника у позвоночных. Человек с одним ухом может, поворачивая голову, искать направление максимальной силы сигнала, потому что голова создает определенную звуковую тень. Два уха делают возможным одновременное сравнение, что позволяет локализовать источник гораздо быстрее и точнее. Если уши достаточно далеко отстоят друг от друга, то создается разница во времени прихода, в фазе звуковых волн, имеющих определенное направление, и их интенсивности. Таким образом, мелкие животные сравнивают только интенсивность звуков, а люди пользуются и монауральными, и бинауральными способами, и долгое время считалось, что они являются видом с наилучшей способностью к обнаружению источника звука. Но благодаря работе Пэйна, одной из первых по этому вопросу, мы знаем теперь, что по слуховым способностям сова сипуха далеко превосходит человека. Сипуха охотится ночью. Она может локализовать и поймать свободно движущуюся мышь в полной темноте. Она даже способна определить направление движения животного и благодаря этому располагает свои когти вдоль длинной оси его тела. Сипуха особенно чувствительна к разнице во времени прихода звука в оба уха. Это позволяет ей определять азимут его распространения. Различия в интенсивности звука также дают сведения о расстоянии до его источника по этому азимуту. В данном отношении сипуха достигает примерно такой же точности, как человек, но примерно втрое точнее его в определении высоты, на которой находится источник звука.
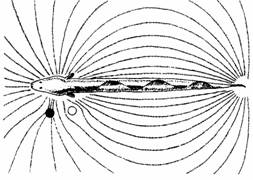
Рис.2. Электрическое поле Gymnarchus. Справа от животного оно не искажено; слева искажено хорошим проводником и плохим проводником. Животное может обнаружить эти два объекта, почувствовав их воздействие на электрическое поле.
Такая точность достигается главным образом благодаря строению лицевой части головы и асимметрии в положении ушей. Правое ухо направлено слегка вверх, а левое - слегка вниз. Правое ухо чувствительнее к высокочастотным звукам выше средней горизонтальной плоскости головы, а левое - к высокочастотным звукам ниже горизонтальной плоскости. При движении звука вниз его высокочастотные компоненты становятся громче в левом ухе и тише в правом. При движении звука вверх происходит обратное. Это дает точную информацию о высоте нахождения источника звука. Таким образом, сипуха пользуется низкочастотными компонентами звуков для локализации их источника в горизонтальной плоскости, а высокочастотными - для определения его положения по вертикали. Она не смешивает эти два типа информации, даже несмотря на то, что оба они основаны на сравнении звуков, приходящих в оба уха.
Самой совершенной заменой зрения является эхолокация, при которой животное испускает высокочастотные сигналы и обнаруживает предметы по возникающему от них эху.

Рис.3. Строение лицевой части головы совы; некоторые перья удалены, чтобы показать асимметричное расположение ушей.
Принцип здесь такой же, как в военных радиолокаторах. Простые формы эхолокации встречаются у землероек, жирных козодоев и гималайских саланганов, которые ночуют и гнездятся в пещерах. Более совершенными ее формами обладают дельфины и другие морские млекопитающие, но своей вершины она достигает у летучих мышей.
После работы Гриффина появилось множество исследований механизма эхолокации у рукокрылых. Теперь мы неплохо знаем физиологию их издающего и принимающего сигналы аппаратов и механизмы мозга, участвующие в эхолокации. Читатель может найти более подробное их описание у Эверта и Гатри.
В процессе эхолокации летучие мыши испускают залпы ультразвуковых, т.е. не слышных для человека, импульсов малой длительности и высокой частоты. Такие короткие импульсы позволяют точно определять время образования эха, а значит, и расстояние до образующего его предмета.
Звуки, производимые другими животными и ветром, обычно бывают низкочастотными, поэтому маловероятно, чтобы на ультразвуковые сигналы летучих мышей накладывались помехи. Лабораторные опыты показали, что искусственные звуки частотой выше 20 кГц дезориентируют полет летучих мышей. Другое преимущество высоких частот состоит в возможности точной фокусировки, что делает возможным распознавание мелких объектов. Рукокрылые производят ультразвуковые сигналы особо устроенной гортанью и испускают их губами, как голоспинные листоносы Pteronotus, или из специальной формы ноздрей, как подковоносы Rhinolophus и представители семейства Phyllostomidae.
У летучих мышей много также специальных приспособлений, позволяющих им определять время и локализовать место возникновения эха от их ультразвуковых сигналов. У большинства рукокрылых, хватающих насекомых на лету, большие наружные уши, форма которых повышает дирекциональную чувствительность.
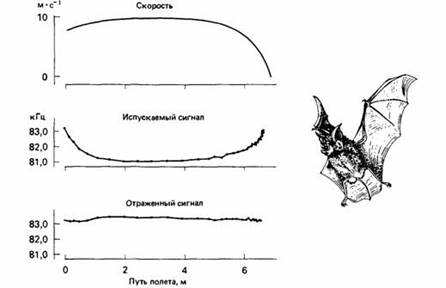
Рис.4. Эхолокация подковоноса, приближающегося к неподвижному предмету.
По мере приближения к нему скорость полета снижается. Летучая мышь приспосабливает частоту своих сигналов так, чтобы частота отражаемого ультразвука оставалась постоянной. После каждого испускаемого животным сильного сигнала эта чувствительность снижается специальными мышцами во внутреннем ухе. При очень коротких испускаемых импульсах, как у представителей сем. Vespertilionidae, конец импульса не перекрывается с началом его эха. Поскольку эхо приходит быстрее от близких объектов, импульсы постепенно укорачиваются по мере приближения объекта, и в результате такое перекрывание устраняется.
У других летучих мышей издаваемые импульсы и эхо от них перекрываются, поэтому, чтобы улучшить обнаружение эха, им нужны другие средства. Например, большой подковонос приспосабливает частоту своих сигналов так, чтобы частота возвращающегося эха поддерживалась в узких пределах. Для мыши, летящей к объекту, воспринимаемая частота эха всегда выше, чем частота испускаемого ультразвука. Это объясняется эффектом Допплера, возникающим вследствие относительного движения животного и объекта: чем быстрее они движутся навстречу друг другу, тем выше воспринимаемая частота излучения. Для компенсации такого эффекта летучие мыши меняют частоту своих сигналов так, чтобы воспринимаемая частота была как можно ближе к постоянной. Таким способом мышь может оценить скорость своего полета, а также направление и относительную скорость полета жертвы.
Как замена зрения эхолокационные способности рукокрылых производят большое впечатление. Лабораторные исследования показывают, что летучая мышь с размахом крыльев в 40 см может пролететь в полной темноте сквозь сетку с ячейками 14 на 14 см из нейлоновых нитей толщиной всего 80 мкм. Летучие мыши научались также ловить мелкие частички пищи, брошенные в воздух в полной темноте, и отличать съедобные предметы от несъедобных по небольшим особенностям их формы. Малая бурая ночница (Myotis lucifugus) может ловить на лету очень мелких насекомых, например плодовых мушек и комаров, с "результативностью" два насекомых в секунду.

Рис.5. Подковонос пролетает через сетку из нейлоновых нитей толщиной 80 мкм с ячейками 14 х 14 см. Слева - длительность и частота эхолокационных сигналов.
3. Зрительное опознание жертвы и хищника
Большинство хищников встречается с множеством разных видов, которые служат им добычей и которых необходимо отличать от других организмов. При этом чаще всего используются три признака - величина, движение и форма.
Когда хищники должны выбирать между особями-жертвами, отличающимися только размерами тела, они обычно хватают самую крупную. Эта стратегия наиболее эффективна в отношении затрат энергии. Однако по мере увеличения размера обычно наступает предел, за которым стимул больше не воспринимается как добыча. Например, когда обыкновенной жабе предъявляют как "добычу" объекты разной величины, она реагирует положительно на экземпляры, размеры которых находятся в определенных пределах, но активно избегает более крупные стимулы.
Как жаба судит о величине объекта? Проще всего делать это по величине изображения на сетчатке, измеряемого градусами угла зрения. Для объекта постоянной величины угол зрения меняется с расстоянием объекта от глаза. Близкие объекты выглядят крупнее, чем удаленные.
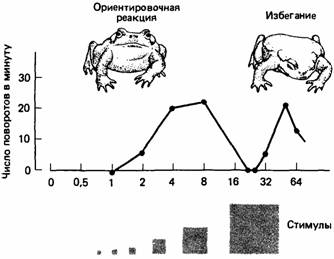
Рис.6. Жабы реагируют на маленькие движущиеся квадраты, ориентируясь в их направлении, но при определенных размерах квадрата эта реакция исчезает и большие квадраты вызывают реакцию избегания.
Чтобы выбрать добычу определенной величины, жабе нужно оценить абсолютную величину видимого объекта, учитывая и его размеры на сетчатке, и расстояние до него. Жабы-повитухи (Alytes obstetricans] в процессе развития постепенно научаются выбирать добычу постоянного размера. Сразу после метаморфоза они предпочитают искусственную приманку некоторого углового размера почти независимо от расстояния. Через шесть месяцев они ориентируются на объекты определенного абсолютного размера независимо от расстояния, т.е. за это время они как-то научаются включать суждения о расстоянии в свои оценки размеров. В некоторых случаях наблюдается склонность к более крупной добыче, но не из-за активного предпочтения, а потому, что она лучше различима. Так, радужные форели чаще ловят крупных ракообразных, чем мелких, потому что крупные видны на большем расстоянии.
Для того чтобы узнать добычу, некоторым видам, например лягушкам и жабам, нужно, чтобы она двигалась. Обыкновенная каракатица в норме нападает только на движущихся креветок. Но если у каракатицы отнять только что пойманную и парализованную ею креветку, то это головоногое немедленно снова нападет на нее даже на неподвижную. Некоторые хищники предпочитают добычу, которая движется беспорядочно. Так, личинки стрекоз предпочитают животных, совершающих зигзагообразные движения, а солнечная рыба скорее нападет на искусственную рыбку, если та извивается, чем на плавно движущийся объект. Иногда имеет значение соотношение формы и движения. Так, когда жабам предъявляют темную движущуюся полосу на белом фоне, они сразу же нападают на нее, когда полоса перемещается вдоль своей оси подобно червяку.
Узнавание добычи по форме - настолько сложное дело, что тут трудно сформулировать какие-нибудь общие правила. В ряде поведенческих работ Робинсон давал в неволе насекомых тамаринам - восновном насекомоядным обезьянам Нового Света. В некоторых опытах им предъявляли богомолов - нормального, без головы, с двумя головами и т.д. Результаты показывают, что для узнавания добычи этим обезьянам важно было опознать голову. В опытах с палочниками в качестве приманки тамарины склонны были пренебрегать насекомыми без ног, но быстро хватали экземпляры с хорошо заметными ногами или маленькие палочки с прикрепленными к ним ножками насекомых. Таким образом, для опознания добычи тамаринам важны головы и конечности насекомых. Чтобы спастись от хищника, насекомые-жертвы выработали разнообразные защитные приспособления, скрывающие конечности и искажающие очертания тела.
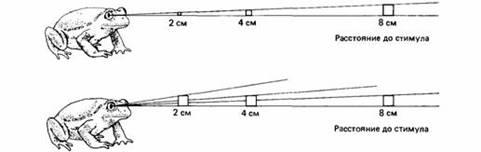
Рис.7. Молодые жабы оценивают размеры по углу зрения, а взрослые жабы могут судить о размерах независимо от угла зрения.
В опытах с абстрактными двумерными фигурами Робинсон обнаружил, что двусторонняя симметрия, вероятно, служит одним из признаков добычи, на который обычно ориентированы хищники.
Те же принципы применимы и к узнаванию жертвами хищников. Так, например, силуэт ястреба, движимый над утятами или гусятами, вызывает реакцию страха, когда движется только в определенном направлении. Это объясняется тем, что короткая шея и длинный хвост характерны для ястреба, а длинная шея и короткий хвост - признаки летящего гуся. Жабы избегают фигур, похожих на змею с поднятой головой. Пиявка, движущаяся толчками, воспринимается как добыча, если ее передняя присоска находится на почве, но если эта присоска поднята в воздух, жаба принимает пиявку за врага.
Как мы видим, поведенческие исследования показывают, что жабы хватают мелкие продолговатые объекты, расположенные горизонтально, но не реагируют на такие же объекты в вертикальном положении. Жаба обычно питается насекомыми, личинками, червяками и т.п.
Ее хищническое поведение состоит из ориентации головы и туловища, зрительной фиксации добычи, захватывания ее в результате вытягивания шеи и языка, глотания и вытирания морды передними лапами. Чтобы вызвать хищническое поведение, нужен мелкий движущийся объект.
На крупные движущиеся объекты жаба отвечает оборонительным поведением. Хотя для изучения сенсорных процессов могут быть применены разные чисто поведенческие методы, физиологическое исследование тоже может давать ценные сведения о деятельности органов чувств и о типе информации, которую они посылают в мозг.

Рис.8. Жабы реагируют положительно на прямоугольники, движущиеся вперед узкой стороной, и отрицательно на движущиеся вперед широкой стороной.

Рис.9. Этот силуэт похож на ястреба, если его двигать в одном направлении, и на гуся, если его двигать в противоположном направлении.
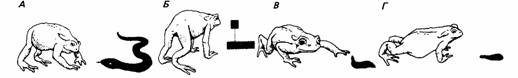
Рис.10 . Реакции жабы на простые модели: А - змея; Б - абстрактный рисунок; В - пиявка с поднятой головой; Г - пиявка с опущенной головой. Первые три вызывают реакцию избегания, последняя исследуется.
Но для того чтобы установить, как мозг использует такую информацию, требуется сочетание поведенческого и физиологического подходов. Такой подход был применен Эвертом и его коллегами в обширных исследованиях по опознанию добычи и врагов жабами.
Физиологические работы показывают, что некоторое опознание добычи происходит на уровне сетчатки. Леттвин и др. отводили электрическую активность от зрительного нерва лягушки, когда объекты двигались в поле зрения. Они обнаружили четыре типа ответов, которые, по-видимому, соответствуют четырем типам ганглиозных клеток в сетчатке. Было установлено, что эти клетки являются детекторами:
1) неподвижной границы,
2) темного выпуклого движущегося объекта,
3) изменения контраста или движения и 4) затемнения. Жаба, очевидно, обладает тремя типами ганглиозных клеток, аксоны которых в составе зрительного нерва идут в зрительную покрышку головного мозга. Информация, поступающая в мозг, включает угловой размер и скорость движения объекта, степень контраста с фоном и общий уровень освещенности. Однако, чтобы распознать добычу, жабе этой информации недостаточно.
Методом дегенерации можно проследить путь волокон зрительного нерва в разные отделы головного мозга, включая зрительную покрышку и претектальную область таламуса. Сетчатка одного глаза проецируется топографически на поверхностные слои противоположной зрительной покрышки. Эта проекция имеет вид карты, на которой каждая область поля зрения соответствует определенной области зрительной покрышки. Электрическая стимуляция покрышки у свободно движущейся жабы приводит к ориентировочной реакции по направлению к соответствующей части поля зрения, т.е. вызывает такое поведение, как будто соответствующая часть зрительной покрышки стимулируется видом добычи. Электрическая стимуляция проекции сетчатки на таламической претектальной области вызывает у жабы реакцию избегания.
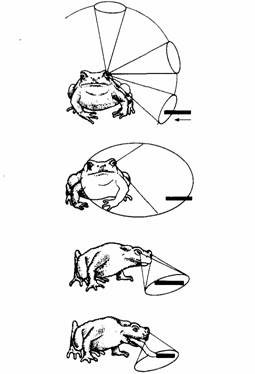
Рис.11. Обнаружение и схватывание добычи жабой. Сверху вниз: добыча попадает в боковое поле зрения животного; жаба поворачивается к добыче, подводя ближе к ней центр поля зрения, а в результате используя бинокулярное поле; затем жаба схватывает добычу языком.
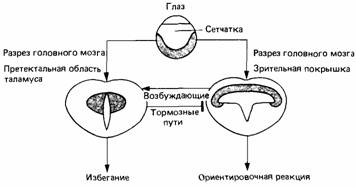
Рис.12. Соотношение между частями головного мозга жабы, участвующими в регуляции избегания и в ориентировочной реакции на зрительно обнаруживаемые объекты.
Хирургическое разрушение части головного мозга приводит к тому, что животные начинают хватать любой движущийся предмет. Разрушение зрительной покрышки уничтожает всякую реакцию на движущиеся стимулы, в том числе и поведение избегания. Эти данные побудили Эверта постулировать, что проекция сетчатки на претектальную область таламуса вызывает поведение избегания, но что для этого также требуется некоторый приток возбуждения от зрительной покрышки. Проекция сетчатки на зрительную покрышку служит основой реакции схватывания всех движущихся объектов, но реакция на крупные или похожие на врага объекты тормозится претектальной областью таламуса и схватываются только мелкие объекты.
Гипотеза Эверта подтверждается физиологическими работами по изучению электрической активности нейронов в зрительной покрышке и претектальной области таламуса в ответ на стимуляцию сетчатки и других связанных с этой функцией частей головного мозга. Это исследование представляет большой интерес не только как демонстрация соответствующего участия сетчатки и мозга в фильтрации стимула, но так же как прекрасный пример того, чего можно достичь разумным сочетанием поведенческих и физиологических методов.