| Скачать .docx | Скачать .pdf |
Курсовая работа: Биологические основы выращивания сазана (Cyprinus carpio L)
Содержание
ВВЕДЕНИЕ
Глава I. Биологическая характеристика объекта в связи со средой обитания и образом жизни
1.1 Общая биология вида
1.2 Эмбриональное развитие
1.3 Постэмбриональное развитие сазана
Глава II. Влияние различных факторов среды на исследуемый объект
2.1 Влияние температуры
2.2 Влияние освещенности, уровня и течения воды на исследуемый объект
2.3 Влияние гидрохимических показателей
Глава III. Управление половыми циклами у рыб различными методами
Глава IV. Биологические основы кормления исследуемого объекта
Глава V. Транспортировка икры, личинок, молоди и взрослых особей выбранного объекта
Глава VI. Биологические основы акклиматизации
Список используемой литературы
Введение
В условиях, когда уловы океанической рыбы и других морепродуктов сокращаются, а рыбные запасы внутренних водоемов находятся в критическом состоянии и поддерживаются в основном за счет искусственного воспроизводства, единственным надежным источником увеличения объемов пищевой рыбопродукции является аквакультура. По данным ФАО в 2000 г. общий мировой вылов всех водных организмов, включая аквакультуру, составил 141,8 млн. т. в том числе промысел - 96 млн. т. (67,7%), аквакультура - 45,8 млн. т. (32,2%). По сравнению с 1990 г. объем продукции аквакультуры увеличился почти на 30 млн. т.
Аквакультура - самая динамично развивающаяся отрасль производства продуктов питания. Аквакультура в России, как и во всем мире, имеет несомненные преимущества перед другими отраслями, производящими белки животного происхождения. Производство продукции аквакультуры является высокоэффективным, что обусловлено прежде всего тем, что гидробионтам не требуется большое количество корма для роста и развития. Важным преимуществом рыбоводства перед другими отраслями сельского хозяйства является также огромная плодовитость рыб. Она позволяет содержать небольшое число производителей для обеспечения крупномасштабного производства товарной продукции.
Высокие репродуктивные возможности рыб, быстрый рост при низких кормовых затратах, наличие маточного поголовья, производство рыбы в местах ее потребления - все это позволяет уже в течение ближайших лет резко увеличить объемы ее выращивания, притом в широком ассортименте - от обычных столовых (карпы, толстолобики, караси, сомы) до деликатесных видов (форель, сиги, стерлядь, осетр, бестер и др.).
В последние десятилетия в большинстве стран мира аквакультура стала приоритетным направлением рыбного хозяйства. По данным ФАО с 1990 по 1999 г. объем продукции мировой аквакультуры увеличится с 18,3 млн. т. до 42,8 млн. т., т.е. в 2,6 раза, и достиг 31,3% мировой морепродукции. Существующая в мире тенденция увеличения доли выращиваемой рыбопродукции по отношению к продукции, полученной за счет океанического промысла, несомненно, станет характерной и для нашей страны, располагающей для этого необходимым потенциалом.
Основным направлением развития аквакультуры нашей страны в настоящее время является прудовое рыбоводство, имеющее многовековую историю развития. Однако особое внимание ему стали уделять с начала 60-х годов прошлого века, когда в технологический процесс выращивания рыбы наряду с карпом был введен комплекс дальневосточных растительноядных рыб и началось интенсивное кормление карпа комбикормами различных рецептур. С этого времени темпы роста и эффективность прудового рыбоводства во многом определялись количеством и качеством используемых кормов. В этом отношении прудовое рыбоводство вступило в конкуренцию с другими отраслями сельского хозяйства.
Однако, как показывают расчеты, корма при выращивании рыбы используются эффективнее, чем при выращивании сельскохозяйственных животных, поскольку соотношение пластического и энергетического обмена у рыб более благоприятно с точки зрения использования пищи на рост, чем у теплокровных животных. Так, при выращивании рыбы в прудах она оплачивает корм приростом массы в 4,3 и 2,5 раза лучше, чем крупный рогатый скот и свиньи соответственно, и на 20% лучше, чем бройлеры. Себестоимость прироста массы товарной рыбы в 3 и 2 раза ниже себестоимости привеса крупного рогатого скота и свиней. Затраты труда на получение 1 т. мяса крупного рогатого скота в 4 раза; свинины - в 1,9; мяса кур - в 2,5 раза выше, чем на производство 1 т. рыбы. На выращивание 1 т. рыбы в прудовом рыбоводстве требуется существенно меньше капитальных вложений, чем на получение 1 т. мяса.
Кроме лучших экономических показателей, получаемых при выращивании прудовой рыбы, она является высококачественным белковым пищевым продуктом, легче усваивается, чем мясо теплокровных животных, и нисколько не уступает ему по составу незаменимых аминокислот и содержанию витаминов. В 100 г. съедобной части рыбы содержится 17,3 г. белка, в то время как в мясе - 16,5 г.
При выращивании рыбы достигается не только экономический, но и экологический эффект, поскольку рыба, в частности растительноядные, является мощным средством оздоровления водной среды.
Положительные стороны аквакультуры и значительный подъем экономического потенциала нашей страны в 70 - 80-е годы определили ускоренное ее развитие в этот период.
Большой объем капитальных вложений позволил в 1975-1990 гг. увеличить площадь нагульных прудов в 1,6 раза. Причем объемы производства рыбы росли не только за счет увеличения ввода новых площадей, но и за счет повышения интенсификации.
Достаточно высокая экономическая эффективность рыбоводства обеспечивалась за счет использования достижений научно-технического прогресса, освоения прогрессивных технологий выращивания рыбы, повышения степени механизации производственных процессов, применения экономических рецептур кормов, успешной селекционно-племенной работы. Научно-технический прогресс осуществляется при достаточно крупном государственном финансировании научных исследований. Например, во ВНИИПРХе объем финансирования завершенных научно-исследовательских работ в сопоставимых ценах за первые два года периода 1989-1999 гг. в 15 раз превышал их объем за последние два года этого периода. В условиях административно-плановой экономики государство поддерживало рыбоводное хозяйство путем установления системы двойных прейскурантов на живую рыбу с возмещением разницы из бюджета.
К сожалению, с переходом экономики страны к рыночным отношениям объемы производства аквакультуры резко упали. Существенное повышение цен на комбикорма, минеральные удобрения и другие материалы привело к увеличению себестоимости рыбы и снижению спроса на нее. Эти и другие негативные моменты, связанные с общей экономической дестабилизацией в стране, привели к сокращению объема выращивания рыбы в начале 90-х годов дол 50-54 тыс. т. И только в последние четыре года наметилась тенденция к увеличению производства рыбы, объем которого в 2000 г. достиг 73,5 тыс. т.
После распада СССР Российская Федерация оказалась самым крупным производителем продукции аквакультуры на постсоветском пространстве. И хотя российские предприятия не миновал спад производства, в стране, несомненно прослеживается тенденция к быстрому развитию аквакультуры.
В жестких условиях становления рыночной экономики чрезвычайно актуальным стало Постановление Правительства РФ от 31.10.1999 г. № 1201 «О развитии товарного рыбоводства и рыболовства, осуществляемого во внутренних водоемах Российской Федерации», в котором были определены задачи по увеличению объемов выращивания и вылова рыбы во внутренних водоемах РФ в 2005 г. на 250 тыс. тонн по сравнению с 1998 г. и с доведением этого объема, начиная с 2006 г. до 600 тыс. т. в год (Рыбоводство и рыболовство, 2001)
Последним важным шагом в установлении прочной законодательной базы в рыбном хозяйстве нашей страны явилось Постановление Правительства РФ от 05.09.2003 г. № 557 «Об утверждении Положения о Государственном комитете Российской Федерации по рыболовству», где, в частности, говорится: «6. Основными задачами Государственного комитета Российской Федерации по рыболовству являются:
1) реализация в пределах своей компетенции государственной политики и осуществление государственного управления в сфере использования, изучения, сохранения и воспроизводства водных биологических ресурсов и среды их обитания;
2) обеспечение в пределах своей компетенции соблюдения требований законодательства Российской Федерации в области охраны, контроля и регулирования использования водных биологических ресурсов и среды их обитания;
3)осуществление в пределах своей компетенции охраны водных биологических ресурсов и среды их обитания;
Основные функции.
Государственный комитет Российской Федерации по рыболовству в соответствии с возложенными на него задачами выполняет следующие функции:
1)разрабатывает и представляет в установленном порядке в Правительство Российской Федерации предложения: об общих допустимых уловах водных биологических ресурсов; о квотах на вылов (добычу) водных биологических ресурсов в пределах общих допустимых уловов;
2)разрабатывает объемы товарного выращивания рыб и других водных животных в прудах, озерах и садковых хозяйствах; правила рыболовства; районы и сроки промысла водных биологических ресурсов; устав службы на судах; рыбопромыслового флота; устав службы органов рыбоохраны; научные программы и ежегодные планы морских ресурсных исследований; нормативы и правила в области использования и воспроизводства водных биологических ресурсов и среды их обитания (в пределах своей компетенции); перечень промысловых видов водных биологических ресурсов.
3) осуществляет в установленном порядке: организацию и ведение государственного учета, государственного кадастра и государственного мониторинга состояния водных биологических ресурсов; организацию выполнения научных программ и ежегодных планов морских ресурсных исследований, изучения состояния водных биологических ресурсов.
4) участвует в установленном порядке: в разработке проектов федерального бюджета на соответствующий год, прогнозов и программ социально-экономического развития Российской Федерации.
Глава I. Биологическая характеристика объекта в связи со средой обитания и образом жизни
1.1 Общая биология вида
ВидСазан (Cyprinus carpio L.)
Тип Хордовые (Chordata)
Подтип Позвоночные (Vertebrata)
Надкласс Челюстноротые (Gnathostomata)
Ряд Рыбы (Pisces)
Класс Костные рыбы (Osteichthyes)
Подкласс Лучеперые (Actinopterygii)
Надотряд Костистые рыбы (Teleostei)
Отряд Карпообразные (Cypriniformes)
Подотряд Карповидные (Cyprinoidei)
Семейство Карповые (Cyprinidae)
Род Сазаны (Cyprinus)
Сазан (CyprinuscarpioL.) – обитает в пресных и солоноватых водах бассейнов Черного, Азовского, Средиземного, Каспийского и Аральского морей, озеро Иссык-Куль, в бассейнах рек Тихого Океана от Амура на севере, до Индокитая на юге (рис.1.).

Рис.1.Сазан.
Образует 2 под вида: типичный, населяющий воды Европы и Средней Азии; и дальневосточный – обитающий в водах Китая и бассейне Амура. В результате акклиматизации сазан и его культурная форма карп теперь расселены по всему Земному шару.
Длина до 1 м., масса до 20 кг (и более). Пресноводная или полупроходная рыба. Полупроходной сазан нагуливается в предустьевых участках моря, а на нерест поднимается в реки. Сазан – быстрорастущая, неприхотливая рыба. Половая зрелость наступает при длине 25-20 см в 3-5 летнем возрасте. Плодовитость высокая – от 96 тыс. до 1,8 млн. икринок. Рабочая плодовитость 300-350 тыс. шт. В Амуре – 450 тыс. шт.
Икрометание происходит весной в прибрежной зоне при температуре не ниже 13 0 С, а разгар нереста наблюдается при 18-20 0 С. Нерест групповой: на одну самку приходится 3-4 самца. Икра откладывается порциями на мягкой растительности обычно в утренние часы. Длительность развития икры от 3 до 6 дней. Вышедшие из икры личинки приклеиваются специальными железами – «цементным органом» к растениям и в течение 5-6 суток питаются содержимым желточного мешка, затем переходят на активное питание. Питание меняется в зависимости от возраста рыбы. Молодь питается планктонными ракообразными, взрослые – ракообразными, моллюсками, червями, водными растениями. Зимуют в глубоких ямах в устьях рек или в предустьевых пространствах. Зимой совершенно или почти полностью прекращает питание и впадает в состояние близкое к спячке. Живет более 30 лет. Имеет большое промысловое значение. Культурная форма сазана – карп – обладает относительно быстрым темпом роста, при небольшой требовательности к условиям внешней среды.
Сазана относят к группе весеннее - летне нерестующих рыб. По отношению к температуре они являются эвритермными, т. е. живут в водоемах, в которых температура воды изменяется в течение года в широких пределах.
1.2 Эмбриональное развитие
Этап I. Перед началом дробления.
Стадия 1.
Икринка не обводняется, в первые минуты после оплодотворения, яйцо прозрачно, пигментированная оболочка плотно прилегает к поверхности яйца.
Стадия 2.
Начало обводнения икринки и образование плазматического бугорка. Оболочка отделилась. Возраст 10 минут.
Стадия 3.
Окончательно обводненная икринка, т.е. перевитиллиновая полость достигла максимальных размеров. Диаметр 1,2-1,3 мм.
Этап II. Стадия бластулы и морулы.
Стадия 4.
Начало дробления, образование двух бластомеров. Возраст 2 часа.
Стадия 5.
Образование четырех бластомеров.
Стадия 6.
Образование восьми бластомеров.
Стадия 7.
Образование шестнадцати бластомеров.
Стадия 8.
Крупноклеточная, или ранняя морула.
Стадия 9.
Мелкоклеточная, или поздняя морула.
Стадия 10.
Бластула. Образуется щелевидная полость – бластоцель.
Этап III. Гаструляция.
Стадия 11.
Начало гаструляции. Бластодерма перемещается по поверхности желточного мешка.
В краевой зоне бластодермы образуются зачатки тела зародыша.
Стадия 12.
Образование желточной пробки.
Стадия 13.
Окончание гаструляции, замыкание желточной пробки.
Этап IV. Формирование эмбриона.
Стадия 14.
Образование глазных пузырей и начало сегментации мезодермы, заметны контуры хорды.
Стадия 15.
Образование глазных бокалов – появление щелевидного углубления в зачатках глаз. Возраст 18 часов, число сегментов 10.
Этап V. Вращающийся эмбрион.
Стадия 16.
Начало обособления хвостового отдела зародыша от желтка. В глазных бокалах появляются хрусталики образование зачатков слуховых пузырей. Тело изредка изгибается.
Стадия 17.
Дальнейшие обособления хвостового отдела.
Стадия 18.
Выпрямление хвостового отдела. Начало активного вращения зародыша внутри оболочки образуется обонятельная капсула.
Этап VI. Вылупление эмбриона из оболочки (рис. 2)
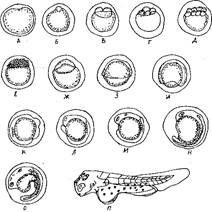
Рис. 2. Развитие икры сазана:
Примечание: А – неоплодотворенная икринка; Б – набухшая икринка с образовавшимся зародышевым диском; В – стадия двух бластомеров; Г – стадия четырех бластомеров; Д – стадия 8 бластомеров; Е – стадия крупноклеточной морулы; Ж – стадия бластулы; З – стадия бластодерма охватывает половину желтка; И – стадия появления зародышевого валика; К – стадия образования первых сомитов в туловище; Л – образование глазных пузырей; М – формирование слуховых плакод; Н – формирование хрусталиков; О – начало пигментации; П – только что выклюнувшаяся личинка.
1.3 Постэмбриональное развитие сазана
Этап развития А.
Через 5 дней после нереста в пруду появляется молодь сазана на этапе А. На этом этапе сазан имеет большой желточный мешок грушевидной формы. Голова слегка пригнута к желточному мешку. Рот нижний, открытый, неподвижный. Глаза пигментированы. Перепончатая жаберная крышка покрывает первые жаберные дужки. Впереди глаза расположена обонятельная ямка, в слуховой капсуле – 2 отолита. Основание грудного плавника расположено вертикально. Плавниковая складка несколько расширена в хвостовой части. Желточный мешок сильно пигментирован. Голова пигментирована слабо, на ней имеются только отдельные пигментные клетки.
Длина головы сазана на этом этапе равняется 13,0%, высота- 15,5% длины тела, длина рыла – 3,6% длины тела, диаметр глаза – 7,7%. Заглазничное расстояние равняется 6,0% длины тела. Высота тела над грудными плавниками – 15,3% длины тела. Высота тела перед спинным плавником – 18,1%, наименьшая высота тела – 5,8%. Длина хвоста – 30,1%, длина хвостового плавника – 5,4%. Личинки висят, приклеившись к растениям или стенки аппарата, время от времени отрываются, затем опять приклеиваются.
На этом этапе развития личинки реагируют на свет положительно. Свободные эмбрионы благодаря наличию на голове органов приклеивания к подвижной растительности приклеиваются ближе к поверхности воды. Этап развития А длится двое суток (рис.3).
Этап развития В.
После двух суток после вылупления личинки сазана переходят на следующий этап развития В. Переход с этапа А на этап В происходит при размере личинки 5,5 мм. Молодь сазана переходит на активное питание остатка желтка пятые сутки при длине 5,7 мм. При переходе личинки на этап В плавательный пузырь наполняется воздухом. Желточный мешок значительно уменьшается. Рот становится конечным, подвижным, но еще не закрывается полностью. Плавниковая складка становится шире. Жаберная перепонка почти полностью закрывает жаберные дужки. Кишка представляет собой длинную прямую трубочку, на голове много пигментных клеток.
Кроме того, пигментные клетки расположены вдоль спины и на нижней поверхности брюха. На этапе развития В длина головы увеличивается до 21,8% длины тела, высота головы доходит до 16,2%, длина рыла та же, что на предыдущем этапе, заглазное расстояние увеличилось до 11,4% длины тела.
Увеличивается высота тела над грудными плавниками до 16,0%. Высота тела перед спинным плавником уменьшается до 15,4% длины тела вследствие уменьшение количества желтка. Наименьшая высота тела увеличивается до 7,0%. На этом этапе личинки плавают поодиночно в толще воды, недалеко от берега. В пруду личинки питаются в основном ветвистоусыми рачками Bosmina. Этап развития В длится четверо суток (рис.3).
Этап развития С1 .
При длине 5,7 мм личинка сазана переходит на этап развития С1 . На этом этапе полностью отсутствует желток. Рот закрывается. Увеличивается плавательный пузырь. Хвостовая лопасть становится больше. Появляется скопление мезенхимы в хвостовой лопасти под задним концом хорды. Немного увеличивается голова, длина ее на этом этапе – 22,9% длины тела, высота головы почти та же, что на предыдущем этапе (16,2%), величина рыла и диаметр глаза почти такая же, что и на предыдущем этапе. Увеличивается заглазничное расстояние до 12,5%. Высота тела над грудными плавниками увеличивается до 17,%. Высота тела перед спинным плавником увеличивается до 16,1%.
На этом этапе развития личинки сазана держатся недалеко от берега на глубине 30-40 см, плавают по одиночке. Питаются они главным образом ветвистоусыми рачками. Этап развития С1 длится четверо суток (рис.3).
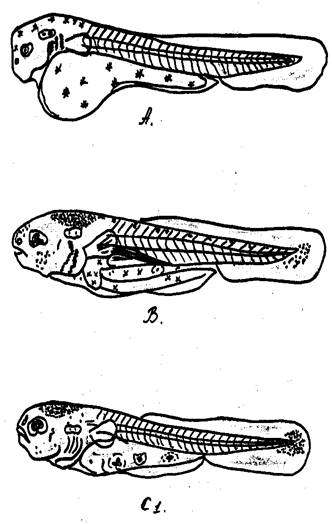
Рис.3 Стадии (А, В, С1 ) постэмбрионального развития сазана
Этап развития С2 .
Личинки сазана переходят на этап С2 при длине 6,5 мм. Этап продолжается до достижения личинкой длины 8,7 мм. Жаберная крышка у них полностью закрывает жабры. В листах будущих спинного и анального плавников – сгущения мезенхимы. В хвостовой лопасти начинают окостеневать лучи. Конец хорды загнут вверх. Хвостовая лопасть несколько вытянута в направлении назад и вверх.
Увеличивается длина головы до 37,9% и высота головы – 17,8%. Длина рыла увеличивается до 4,2%. Заглазничное расстояние и диаметр глаза почти такие же, как и на предыдущем этапе. Высота тела над грудными плавниками увеличивается до 19,4%, высота тела перед спинным плавником – до 19%, наименьшая высота тела не изменяется по сравнению с высотой на предыдущем этапе, длина хвоста уменьшается до 26.5%. Держится личинка недалеко от берега на мелководье по одиночке. На этом этапе развития личинки по прежнему продолжают питаться главным образом ветвистоусыми рачками, но впервые на этом этапе развития в их пище начинают встречаться личинки хирономид. Этап С2 длится трое суток (рис.3а).
Этап развития D1 .
При длине 8,7 мм молодь сазана переходит на этап развития D1 . На этом этапе плавательный пузырь становится двойным, передняя камера его начинает наполняться воздухом. Рот конечный, слабо выдвижной. В спинном и анальном плавниках появляются мезенхимные лучи. Появляются зачатки брюшных плавников. Лучи в хвостовом доходят до его заднего края. Задний конец хорды сильно загнут вверх. Хвостовой плавник трехлопастной. Длина головы у сазана остается почти такой же, как на предыдущем этапе. Диаметр глаза увеличивается до 8%. Увеличивается заглазничное расстояние до 14%, высота тела над грудными плавниками до 20,7%, высота перед спинным плавником до 22% длины тела. Немного увеличивается наименьшая высота тела (до 6,8% вместо 6,0% на предыдущем этапе). Длина хвоста уменьшается до 23,6%. Длина хвостового плавника увеличивается до 9,9%. Личинки сазана, до сих пор держатся у поверхности воды, к этому времени опускаются на глубину до 0,5 м. Держатся они все еще поодиночке. Пищей в прудах им служит главным образом организмы тех же размеров, что и на предыдущем этапе. Этап развития D1 длится трое суток (рис. 3а).
Этап развития D2 .
Личинки сазана при длине тела 9,4 мм переходят на этап развития D2 . НА этом этапе развития хвостовой плавник двухлопастной. Выемка в нем увеличивается и начинается расчленение его лучей. В спинном и анальном плавниках развиваются костные лучи, доходящие в каждом плавнике до его края. Увеличиваются брюшные плавники. В грудных плавниках появляются мезенхимные лучи. Плавниковая складка от спинного плавника тянется до основания хвостового и под его основанием. Рот выдвигается сильнее, чем на предыдущем этапе.
Относительный размер головы составляет 30,4% длины тела. Высота головы увеличивается до 18,5%. Увеличивается длина рыла – до 5,9%, диаметр глаза до 8,8%, заглазничное расстояние до 15,8%, высота тела над грудными плавниками до 23,1%, высота тела перед спинным плавников до 25%, наименьшая высота тела до 9,1%. Длина хвоста увеличивается до 22,1%.
Сильно увеличивается длинахвостового плавника до 18,6%. Личинки сазана держатся недалеко от берега на глубине 0,5 м поодиночке. Утром они приближаются к берегу, к вечеру отходят от него и спускаются в более глубокие слои воды. В кишечнике молоди сазана на этом этапе встречаются в основном Bosmina и некоторое количество личинок хирономид. Количество личинок хирономид в пище по сравнению с тем, что было на предыдущих этапах (С2 и D1 ) увеличилось. Этап развития D2 длится пять суток (рис. 3а).
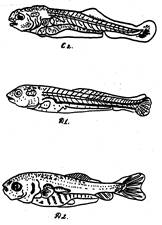
Рис.3 а Стадии (С2 , D1, D2 ) постэмбрионального развития сазана
Этап развития Е.
Длина тела личинки – 1,8 мм. К концу этого этапа во всех парных и непарных плавниках уже развиты костные лучи. Спинной плавник легка выемчатый. В хвостовом плавнике выемка увеличилась. Преанальная складка небольшая. Над и под хвостом плавниковая складка полностью редуцировалась. Обонятельная ямка начинает принимать форму восьмерки, т.к. образует небольшие выросты от верхнего и нижнего краев. Увеличилась голова, длина ее – 33%, высота – 21,4%, длина рыла увеличилась до 7%, заглазничное расстояние до 16.2%, высота тела над грудными плавниками до 25,5%, высота тела перед спинным плавником до 29,5%, высота тела наименьшая увеличилась незначительно до 9,8%. Увеличилась длина хвостового плавника до 21,9%. На этом этапе развития Е личинки сазана держатся поодиночке на глубине около 0,5 м и струе (у водослива). Питается сазан на этом этапе главным образом личинками хирономид. Этап Е длится трое суток (рис.3 б).
Этап развития F.
Молодь сазана на этап развития переходит при длине 14,8 мм. При переходе н этот эта у сазана вдоль продольной горизонтальной миосепты начинает развиваться чешуя, к концу этапа она покрывает почти все тело. В углах рта появляется у рта первая усиков. Отверстие обонятельной ямки принимает форму восьмерки благодаря увеличению в ней двух встречных выростов ее дорзального и вертикального краев. Длина головы увеличилась до 35,3%, длина рыла – до 8,5%, уменьшился диаметр глаза до 7,9%, заглазничное расстояние стало уменьшаться (15% вместо 16,2% на предыдущем этапе). Высота тела над грудными плавниками почти та же, что на предыдущем этапе, увеличилась высота тела перед спинным плавником до 32,6%, увеличилась наименьшая высота тела до 12,1%, уменьшилась длина хвоста до 21,9%, длина хвостового плавника увеличилась до 22,8%. Мальки сазана на этом этапе держатся преимущественно у дна, часто около водослива. При тихой погоде мальки плавают стайками. Отличаются большой пугливостью. На этом этапе молодь сазана, вероятно переходит на донное питание. В ее пище встречается много личинок хирономид. Длится этап шесть суток (рис.3б).
Этап развития Q.
Основная масса молоди сазана переходит на этап развития Q при длине тела 21,7 мм. На этом этапе сазан полностью покрывается чешуей. Обонятельное отверстие становятся двойным, появляется зачаток канала боковой линии.
Увеличивается высота тела над грудными плавниками до 29,4%, перед спинным плавником до 34,1%. Остальные пропорции тела остаются почти такие же, как на предыдущем этапе. Мальки обычно плавают стайками, реже поодиночке, недалеко от берега (рис.3б).

Рис. 3 б. Постэмбриональное развитие сазана стадии (Е,F,Q).
Глава II. Влияние различных факторов среды на исследуемый объект
2.1 Влияние температуры
Основным абиотическим фактором, определяющим границы возможного распространения растительноядных рыб в пресных водоемах, является температурный режим. Температурный оптимум у карпа несколько ниже, чем у других растительноядных рыб. Температура действует на рыбу как непосредственно – изменяя интенсивность ферментативных процессов, происходящих в организме, активность потребления пищи, характер обмена веществ, ход развития половых желез и прочее, так и косвенно, оказывая свое влияние на улучшение или ухудшение развития естественной кормовой базы. У сазана при температуре 26-28 снижается потребление кислорода, а при температуре свыше 35 0 C он может погибнуть. Сазан относится к теплолюбивым видам рыб, нерест у него весеннее – летний, при температуре воды 8 -20 0 C, иногда при температуре 17 -25 0 C. Икра его развивается обычно при тех же температурах, при которых происходит нерест. При постепенном повышении или понижении температур (по отношению к оптимальным температурным условиям) нормальные течения жизненных процессов в организме рыбы нарушаются, она не размножается и неохотно потребляет корм, дыхание и кровообращение замедляются. Резкое изменение температуры воды, если даже она и не выходит за границы оптимальных температурных условий, вызывает у рыб нервный шок, который приводит обычно к гибели. В зависимости от температуры воды изменяется количество растворенного в ней кислорода, который необходим для дыхания рыб. Так, при понижении температуры содержание кислорода в воде повышается, а при понижении снижается. Обмен веществ и потребление кислорода увеличивается у рыб с повышением температуры воды и, наоборот, уменьшается с ее понижением. Температура воды оказывает также большое влияние на развитие в водоеме кормовых организмов, являющихся пищей для рыб. Отклонение температуры воды от оптимальной для организмов зоопланктона вызывает снижение их численности и биомассы. Рыбовод должен постоянно контролировать температуру воды на рыбоводных предприятиях, а при необходимости, обусловленной биологической целесообразностью, изменять ее до желаемой величины.
2.2 Влияние освещенности, уровня и течения воды на исследуемый объект
Известно, в результате таяния снегов и дождей повышается уровень воды в реке, увеличивается скорость течения, мутность и, следовательно, снижается степень прозрачности воды, которая существенно влияет на ее освещенность. Прозрачность воды зависит от содержания в толще воды неорганических и органических взвешенных частиц, а также мельчайших растительных и животных организмов. Большое количество взвешенных частиц глины и песка в воде вызывает отмирание фито– и зоопланктона, затрудняет дыхание рыб и ухудшает их питание. Мутность воды, образованная взвесью частиц из отмерших растительных и животных организмов, ухудшает гидрохимический режим водоема. Поступление в реку воды и резкий подъем уровня, увеличивают скорость течения – немаловажные абиотические факторы в жизни рыб.
Освещенность имеет большое значение, от нее зависит строение органов зрения, которые играют значительную роль для ориентировки рыб во время движения. Освещенность влияет на развитие рыб. Влияние освещенности на рост и выживаемость личинок сазана в последние время подвергается тщательному изучению. У личинок кроме жаберного полноценное значение имеет дыхание через покровы тела, в том числе с участием пигментов. Возможно, поэтому в этот период жизни личинок сазана находятся в поверхностных слоях воды, а свет, вероятно, является обязательным фактором их существования. При уменьшении освещения, особенно в условиях дефицита кислорода усиливается пигментация личинок. Личинки выглядели черными и через некоторое время погибали. Возрастные особенности отношения личинок сазана к освещенности связанны не только с физиологическими процессами. Наблюдается уменьшение роли освещенности в питании личинок по мере их роста и развития. В начале развития личинок в отсутствии корма предпочитают меньшую интенсивность освещения. В зонах с меньшим освещением личинки были малоподвижны, тогда как при большом освещении они совершали постоянные активные перемещения.
2.3 Влияние гидрохимических показателей
Значение солевого состава в жизни рыб огромно. От солевого состава и количества растворенных в воде минеральных солей зависит развитие одноклеточных водорослей – пищи для беспозвоночных животных, а также рыб. Фосфор и кальций имеющие важное значение при формировании костной ткани и синтезе белков, рыбы могут получать не только из пищи, но и непосредственно из воды. Магний, калий, натрий, серу, железо, медь йод, фтор, молибден и другие химические элементы, необходимые для нормального роста и развития, они могут также получать из воды. Однако рыбовод должен помнить, что повышенное содержание в воде той или иной соли может оказать на рыбу вредное воздействие, а в некоторых случаях вызывать ее гибель. Растворенные в воде минеральные соли поддерживают у рыб постоянное осмотическое давление, обеспечивающее работу всех внутренних органов: всасывание в кровь через стенки кишечника питательных веществ, а также выделение продуктов обмена. Кислород необходим рыбам для дыхания. Они ассимилируют его из воды. По отношению к содержанию кислорода в воде сазана можно отнести к группе рыб, которые могут жить при небольшом количестве кислорода в воде (4-5мг/л). Содержание кислорода в воде изменяется в зависимости от температуры, атмосферного давления (чем выше давление, тем больше растворимость кислорода), интенсивности перемещения водных слоев, от солености (чем выше соленость, тем меньше растворяется кислород). Недостаток кислорода вызывает массовую гибель рыб (замор), это происходит при содержании кислорода менее 2мг/л. Наиболее благоприятными для жизни рыб, являющихся объектами массового искусственного разведения, нейтральная или слабощелочная реакция среды (pH 7,0 – 7,5). Рыбы являющиеся объектами прудового рыбоводства, а в частности и сазан, переносят значительные колебания pH. Влияние активной реакции среды на жизнь рыб изменяется в зависимости от солевого состава воды. Понижение значения pH повышает требовательность рыб и их икры к концентрации кислорода в воде. При кислой реакции кислорода в воде должно быть значительно больше, чем при нейтральной или слабощелочной реакции. Сазан способен расти в озерах, где в воде присутствуют гидрокарбонаты и сульфаты до 2 – 3,5г/л, хлориды до 8 – 12г/л. При pH менее 6,0 и более 9,0 резко возрастает отход личинок карповых рыб при посадке в выростные озера. Для личинок карповых допустимым считается pH более 6,0. Личинки карповых рыб особенно чувствительны к дефициту кислорода из-за высокой интенсивности обмена. Для нормального роста и развития личинок достаточно 5-6мг кислорода /л.
Глава III. Управление половыми циклами у рыб различными методами
Длительное содержание производителей в прудах при нерестовой температуре приводит к их перезреванию. Поэтому нерестовую кампанию следует проводить в возможно сжатые сроки 25-30 дней. Производителей содержат в зимовальных прудах до начала нерестовой кампании. Перед началом работы по искусственному разведению производят разгрузку зимовальных прудов. Отлов производителей из зимовалов проводят по воде хамсаросовым неводом. Из невода рыбу отбирают с помощью матерчатых рукавов длиной 1-1,3 м, посаженных с одной стороны на металлический обруч, диаметр которого 30-35 см. Отловленных производителей переносят в носилки с водой, снабженные брезентовыми крышками. Длина носилок 1,2-1,5 м, ширина – 40-45 см. При загрузке зимовалов производителей сортируют по видам, полу и степени готовности к нересту. Готовность к нересту определяется по внешнему виду рыб. Самок обычно делят на три группы: первая группа – лучшие, наиболее зрелые самки. Брюшко самки мягкое на ощупь, отвислое. Иногда заметна припухлость в области генитального отверстия. Эту группу самок используют для работы в первую очередь. Вторая группа – самки с аналогичными внешними признаками, но менее ярко выраженными. Такие самки могут быть использованы позднее, после окончания работы с самками первой группы. Третья группа – самки по внешнему виду почти не отличимые от самцов. Таких самок для получения икры не используют, а сразу же высаживают на летний нагул.
Самцов по внешним признакам делят на две группы: первая группа – самцы легко отдающие молоки. Внутренняя поверхность грудных плавников шероховатая. При движении пальца от конца плавника к телу у п. т. они менее острые, в виде бугорков. Вторая группа – самцы выделяют очень мало молок или вовсе не текут. Таких самцов используют в конце сезона или не берут для работы вообще.
После сортировки производителей отсаживают в пруды для преднерестового содержания, где они находятся до использования для получения потомства. Для преднерестового содержания производителей наиболее пригодны не большие, легко облавливаемые водоемы (площадь 0,05-0,2 га), глубина 1,5-2 м. В мелких, хорошо прогреваемых прудах производителей содержать нельзя, т. к. они быстро перезревают. Пруды для преднерестового содержания должны быть хорошо спланированы, быстро осушаться и заполняться водой (не более 2-3 ч). Желательно обеспечить в этих прудах постоянный водообмен, чтобы предотвратить чрезмерный прогрев вод. Посадка производителей в пруды для преднерестового содержания допускается до 1000 шт./га (но не более 150 ц/га). В прудовых хозяйствах единственным способом, обеспечивающим в настоящее время получение половых продуктов сазана является метод гипофизарных инъекций, т. е. физиологический метод. После введения вещества гипофиза рыбы переходят в нерестовое состояние независимо от наличия нерестовой обстановки. Для созревания необходимы хорошие кислородные условия и температура воды не ниже 19-20 0 С. В рыбоводстве пользуются ацетонированными гипофизами. Для получения половых продуктов от карповых рыб используют гипофизы половозрелого сазана, заготовленные в преднерестовый период. Гипофизарная инъекция способна стимулировать созревание самок, имеющих яичники в завершенной IV стадии зрелости. Делать гипофизарную инъекцию рыбам, находящимся на более ранних этапах созревания, бесполезно. Установлено, что преднерестовые изменения в овоцитах, превращающих их в зрелые икринки. Вторым – овуляция, т. е. освобождение икринок из фолликулярной оболочки, удерживающей ее в яичнике. I этап созревания осуществляется под влиянием очень малых доз гонадотропного гормона. II этап – под воздействием значительно большего количества гормонов. Первый раз вводится очень малая доза гормона, составляющая 1/8-1/10 часть общей намеченной дозы гормона. Через сутки после такой предварительной инъекции проводится вторая - разрешающая инъекция. Количество гипофиза колеблется в пределах 3-6 мг на 1 кг массы самки. Оно может изменяться в зависимости от степени зрелости гонад. Практически дозу определяют, перемножая массу самки на выбранное в интервале от 3 до 6 мг количество гипофиза и корректируют то количество в последующих партиях в зависимости от того, на сколько полно самки отдают икру. Для работы обычно используют текучих самцов. Однако для получения достаточного количества молок самцам также делают гипофизарную инъекцию. Инъекцию самцам делают за час до проведения разрешающей инъекции самкам. Самцам массой 5-7 кг вполне достаточно ввести 4-6 мг гипофиза на рыбу. При необходимости получения большего количества молок и большим крупным самцам, имеющим часто массу до 10 и более кг следует вводить по 12-15 мг вещества гипофиза на рыбу. При вылове из преднерестовых прудов самок взвешивают и измеряют, после чего близких по массе и величине гонад самок группируют по партиям, что упрощает проведение инъекций. Инъекцию проводят в носилках с водой.
Рекомендуется следующий порядок проведения работ. В первый день в 18-19 часов производится отлов самок в преднерестовых прудах для предварительных инъекций. Количество отобранных для инъекций самок определяется их рабочей плодовитостью и мощностью инкубационного цеха. На следующий день в те же часы производится разрешающая инъекция самкам за 1 час до инъецирования. На 2-3 самки берут одного самца. После инъекции производителей помещают в нерестовые садки.
Температура воды играет важную роль при получении половых продуктов методом гипофизарных инъекций. При оптимальных температурах и правильном определении дозы гипофизарных инъекции от одной пары производителей можно получить икру и молоки значительно больше и лучшего качества, чем при крайних температурах и несоответствии дозы гипофизарных инъекций. У производителей сазана гормональная стимуляция проводится при наступлении нерестовых температур, раннем получении икры в условиях регулируемого температурного режима, при температуре воды ниже нерестового порога в условиях нерегулируемого температурного режима. В зависимости от степени зрелости половых продуктов и температуры воды используют разные схемы гипофизарных инъекций. Известно, что оптимальной температурой для получения половых продуктов от сазана является 17-22 0 C. Инъецируют производителей в рыбоводной люльке с мягким покрытием или непосредственно в бассейне, приспуская воду настолько, чтобы верхняя часть спины рыбы находилась над водой. Время начала инъецирования рассчитывают таким образом, чтобы получение половых продуктов приходилось на дневное время. Для инъецирования применяют шприцы вместимостью 10-20мл. При инъекции игла вводится в спинную мышцу, позади головы, несколько выше боковой линии. Место введения раствора после извлечения иглы нужно прижать пальцем и одновременно слегка промассажировать. Для предупреждения травмирования производителей при проведении инъекций и контроля за созреванием и сцеживанием половых продуктов применяются различные анестезирующие средства - ихтиокальм, хинальдин и другие. Рыб содержат в анестезирующем растворе в период проведения той или иной операции, а затем помещают в чистую воду.
Скорость созревания самок сазана в зависимости от температуры воды
Таблица №4
| Температура воды, 0 С | Время созревания самок, ч. |
| 20-22 | 10-12 |
| 23-25 | 9-11 |
| 26-28 | 7-10 |
Существует три схемы гипофизарных инъекций.
По первой схеме при нерестовых температурах воды получение зрелых производителей обеспечивается однократной инъекцией. Доза гипофиза составляет 2-2,5мг на 1кг массы для самки, для самцов - в 2 раза меньше. Время содержания самок при нерестовых температурах до инъекции 4-5 суток.
По второй схеме при раннем получении икры в условиях регулируемого температурного режима во всех случаях необходимо дробную схему гипофизарных инъекций. В зависимости от степени зрелости яичников эта схема применяется в нескольких вариантах:
а) в диапазоне нерестовых температур стабильные результаты созревания самок можно получать при двукратном введении им гонадотропного материала; величина доз гипофизарных инъекций в зависимости от температуры воды различна: при температуре 17-18 0 C первая доза равна 0,5мг, вторая-2,5мг на 1кг массы самки; при температуре 19-20 0 C первая доза составляет 0,3мг, вторая 2мг на 1кг массы самки; промежуток между первой и второй инъекциями должен быть 12 часов; при этом можно получить одновременное созревание самок;
б) хороших результатов созревания (90-100%) можно добиться при постепенном введении увеличивающихся доз гонадотропного материала; для стимуляции развития ооцитов, ядра которых находятся еще в центре, наиболее эффективно применять трехкратные гипофизарные инъекции.
По третьей схеме инъекции будут зависеть от степени зрелости половых продуктов у самок. Так, от производителей, яичники которых находятся в состоянии, близком к зрелости можно получить икру при двукратной схеме введения гонадотропного материала. Если же яичники самок находятся в состоянии, далеком от зрелости, созревание самок можно достичь при трехкратном введении гонадотропного материала постепенно повышающимися дозами.
Глава IV. Биологические основы кормления исследуемого объекта
Естественная пища сазана состоит из животных обитающих в грунтах (зообентос) и в толще воды (зоопланктона), высшей растительности (зарослевая фауна или перифитон), остатков высшей растительности и их семян. Зообентос, как правило, представлен в основном личинками насекомых. К ним относятся личинки комаров Chironomus называемые в быту «мотылем», стрекоз, жуков Dutiscus и. т. д. Молодь Сазана после вылупления из икры питается остатками желтка и мелкими формами планктонных организмов, таких как инфузории и коловратки. К питанию личинками и мелкими формами зоопланктона, молодь переходит при достижении длины 7мм и массы 4-5мг. При массе 5-10мг и длине 8-11мм в их пище наряду с ветвистоусыми и веслоногими рачками появляются планктонные формы личинок хирономид. По мере дальнейшего роста масса 15-20мг, длина11-13мм в пищевом комке встречаются крупные формы зоопланктона и возрастает количество хирономид. При масее100-300мг и длине 18-25мм на ряду с зоопланктоном большое место в питании начинают занимать бентосные организмы, появляются водоросли. Во взрослом состоянии Сазан имеет широкий спектр питания. Излюбленной пищей являются донные организмы: личинки хирономид, олигохеты, моллюски. Охотно питается зоопланктоном. Может потреблять растительность и детрит. Способность Сазана, как и других карповых рыб, адаптироваться к разнообразным условиям питания, обеспечила ему в процессе эволюции возможность освоения различных экологических ниш, а также послужила биологическим обоснованием для применения растительных кормов при его выращивании в прудовых хозяйствах.
Переваримость пищи у Сазана
Показатель переваримости характеризует то количество питательных веществ корма, которое поступает в организм рыб после осуществления пищеварительных процессов. Из основных питательных веществ наиболее быстро и полно расщепляются и всасываются белковые соединения. Переваримость белка колеблется в среднем в диапазоне 70-80%, а составляющих его аминокислот 45-90%. Углеводная часть кормов в целом доступна организму рыб значительно хуже, чем белковые вещества (в среднем 35-55%). Их переваримость находится в диапазоне 10-35%.
Также для нормального роста и развития карпам требуется определенный набор питательных веществ, включающий белки, углеводы, липиды, минеральные вещества, витамины. Сбалансированный состав выше перечисленных веществ используется в рецептурах комбикормов для карповых. Например, ВБС-РЖ-85 – продукционный комбикорм (табл. №5).
Рецепт ВБС-РЖ-85
Продукционного комбикорма для выращивания в прудовых хозяйствах сеголетков карпа, массой от 0,5 до 25 г и более
Таблица №5
| Компоненты | % |
| Соевый шрот | 9 |
| Подсолнечный шрот | 20 |
| Пшеница | 40 |
| Горох | 10 |
| Паприн | 16 |
| Отруби | 3 |
| Мел | 1 |
| Кормовая крупка из отходов филлофоры | 1 |
В 100 г гранулированного корма содержится, г:
Влаги не более 13,0
Сырого протеина не менее 26,0
Сырого жира 1,0-1,5
Сырой золы 5,4
Сырой клетчатки 6,0
Потребность в белках и аминокислотах
Белки являются одним из главных элементов клеток и тканей и выполняют широкий диапазон функций. Они входят в состав клеточных мембран и обеспечивают структурную эластичность и жесткость мышц, эластичность скелета и тканей других органов, входят в состав ферментов. Обеспечивают защитную функцию, являясь основой антител, и принимают участие в процессах регуляции обмена веществ в составе гормонов. Белки в отличие от жиров и витаминов не откладываются в запас. При недостатке их в организме или голодании происходит разрушение цитоплазмы клеток и в первую очередь мышц и печени. Степень полноценности белка пищи во многом зависит от его химического состава. В составе белка обычно около 20 аминокислот. Десять из них (лизин, аргинин, гистидин, треонин, лейцин, изолейцин, Валин, метионин, триптофан и фенилаланин) считаются незаменимыми, так как они не синтезируются из других веществ и должны поступать с пищей. Проявление недостаточности отдельных незаменимых аминокислот в питании рыб, не имеют в большинстве случаев яркой клинической картины и чаще всего характеризуются признаками, которые имеют место при неполноценном питании вообще.
Это снижение темпа роста, потеря аппетита, снижение общей резистентности (устойчивости) организма. Количественные потребности в протеине любого организма, в том числе и карпа, неодинаковы на протяжении жизни. Они изменяются в зависимости от стадии развития рыб, экологических условий (температура, кислород) и состояния здоровья. Быстрорастущая молодь, личинки и мальки нуждаются в большем количестве белка, чем взрослые рыбы. Согласно научным данным содержание белка в корме личинок и мальков карпа должно составлять 45-40%. С возрастом потребности карпа в белке расширяются. При выращивании в прудах молодь массой 10-15г хорошо растет на кормах с 28-30% содержанием белка, массой100-200 г- с 26-23%, более 200 г- с 23-20%.
Потребность в жирах
Жиры как питательные вещества являются источниками энергии и содержат в своем составе многие жизненно важные вещества, такие, как жирорастворимые витамины, незаменимые жирные кислоты и. т. д. Значение жиров в пище рыб не ограничивается ролью поставщиков энергии. По своим функциям в организме они служат структурными и рецепторными компонентами клеточных оболочек, а также являются передатчиками биологических сигналов. Потребность карпа в жирах точно не установлена по данным ученых, карп без видимых вредных последствий может переносить до 40% доброкачественного жира в корме. Наиболее типичными признаками дефицита жира в организме являются: замедление роста, снижение аппетита, заболевания кожи и плавников, выражающиеся в нарушении их пигментации и последующем некрозе. При содержании жиров в комбикормах менее 2,5% приводит к нарушению обменных процессов, что приводит в организме рыб к снижению эффективности использования белков и комбикорма в целом. Карпы выращиваемые в прудах, получают жиры не только с комбикормами, но и с естественной пищей. Карп должен получать еще и арахидоновую кислоту, что обусловлено его теплолюбивостью. Поэтому для молоди карпов, обладающей очень высокой скоростью роста, обеспеченность естественной пищей как источником незаменимой арахидоновой и других жирных кислот имеет важное значение.
Снижение обеспеченности рыб естественной пищей в два раза приводит к дефициту незаменимых жирных кислот, снижению активности питания рыб в 1,5-2 раза, угнетению роста, уменьшению в их теле общих запасов белка, жира и увеличению воды. Старшие возрастные группы (осенние двухлетки, трехлетки и далее) имеют определенные запасы незаменимых жирных кислот, отложенные в жировых депо, поэтому симптомы недостаточности проявляются у них менее остро, чем у молоди. Обогащение комбикормов растительными жирами (7-8%) стало применяться в качестве профилактики жаберных заболеваний. Добавление в комбикорма для карпа жиров, повышает их продуктивное действие, усиливает общую устойчивость организма в стрессовых ситуациях и активизирует темп роста.
Потребность в углеводах
Известно, что часть энергетических потребностей животного организма даже при наличии других источников обязательно должна покрываться за счет расщепления углеводов. Питательная ценность углеводов тесно связанна с их химическим строением и соотношением отдельных групп углеводов в кормах. Немаловажное значение имеет их удобоваримость, т. е. возможность пищеварительной системы рыб осуществлять расщепление и всасывание этих соединений. Конечными продуктами расщепления углеводов являются моносахариды. Их делят на две подгруппы: пентоз и гексоз. Известно, что у карпа наиболее быстро и полно всасываются гексозы, в частности галактоза и глюкоза. Оптимальное количество углеводов для карпа составляет 40-50%.
Потребность в минеральных элементах
Рациональное кормление рыб должно удовлетворять их потребности как в органических, так и в минеральных веществах, так как только в этих условиях может быть обеспечен нормальный рост и развитие организма. Минеральные вещества выполняют структурную функцию, входя в состав опорных элементов костной ткани и оболочек всех тканей. В составе различных соединений они участвуют в процессах переваривания и всасывания, синтеза и распада, а также обезвреживания ядовитых веществ и выделения. Минеральные вещества обеспечивают осмотическое давление и постоянство других физико-химических свойств внутренней среды организма. По количественному содержанию в животных и растительных тканях минеральные элементы делятся на макро- и микроэлементы. К макроэлементам относят кальций, фосфор, магний, калий, натрий, хлор, серу. Их содержание в организме рыб колеблется в широких пределах, составляя более 100мг/кг. К микроэлементам относят железо, медь, марганец, цинк, йод и др. (содержание в пределах 90-0,01мг/кг). В среднем содержание общей суммы минеральных элементов в теле рыб составляет 2,5-6%. Особенностью минерального питания рыб является наличие двух путей поступления элементов в организм рыб: с пищей через рот и посредством осмотического проникновения из воды через жабры и покровные ткани. В зависимости от того, в какой воде ведется выращивание (пресной или солоноватой) рыбы вынуждены постоянно либо восполнять недостаток минеральных веществ, либо экскретировать их избыток. Ограниченное или избыточное поступление минеральных элементов в организм рыб, может привести к снижению аппетита, возникновению патологических изменений, торможению роста. Например, при совместной или раздельной недостаточности в рационе фосфора, кобальта, магния, цинка развивается остеодистрофия которая проявляется в искривлениях позвоночника, деформации лобных и челюстных костей, ротового аппарата, редукции жаберных крышек. Потребность карпа в кальции зависит от возраста и массы рыб. Известно, что молодь нуждается в большем его количестве, чем старшие возрастные группы. Максимальная потребность отмечена в период окостенения скелета и развития чешуи. Это относится к молоди массой около 100мг. При повышении температуры и активизации обменных процессов в организме рыб потребность в кальции возрастает.
Глава V. Транспортировка икры, личинок, молоди и взрослых особей выбранного объекта
Растительноядных рыб перевозят в живорыбных вагонах и автомашинах и чаще всего в полиэтиленовых пакетах авто – и авиатранспортом (рис.4.).
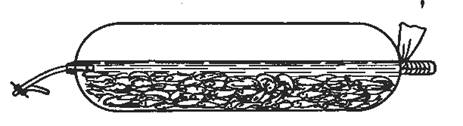
Рис. 4. Полиэтиленовый пакет для перевозки живой рыбы
Посадочный материал укладывают в фанерные, жестяные ящики с поперечными планками размером 60x75x10 или 55x55x10см. Ящики устанавливают один на другой по 7-8шт. в стопке. Дно выстилают мхом, марлевыми тюфячками, матами из рогоза и камыша. Во время полета рыбу орошают охлажденной водой. Погрузка рыбы длится быстрее не более 15-20 минут, а разгрузка – не более 10. При перевозке в живорыбных вагонах в течении до 12 суток в один вагон загружают до 800кг трех – и четырехлетков сазана, они хуже переносят перевозку чем белые амуры. Для молоди принята норма 30 тысяч штук на каждую живорыбную машину. В этих условиях при длительности перевозки 10 часов отход не превышает 5%. Сеголетков достигнув веса 15 гр, перевозят в этих машинах при температуре воды 5 -10 0 C по норме 5 тысяч штук на каждую. Во время пути необходима постоянная аэрация при помощи компрессора. Рыбы большего веса – 500-700г при длительности транспортировки 25 часов перевозятся при температуре 5-8 0 C и норме посадки в одну автомашину в количестве 300-400 штук. Широко используют полиэтиленовые пакеты шириной 42-44см. (при толщине пленки 0,1-0,14мм). Для перевозки производителей и вообще крупной рыбы применяют пакеты объемом 50-80л. Для рыб старшего возраста длина пакета увеличивается до 1,35-1,55м. В соответствии с возрастом и условиями перевозки транспортируемой рыбы пакеты изготавливают из разного количества слоев полиэтилена: для личинок – двухслойные, для рыб старших возрастов трехслойные. Перед транспортировкой, производителей выдерживают несколько суток без пищи, отмывают в носилках от грязи и слизи и затем загружают в пакеты. Пакеты перевозят в наклонном состоянии под углом в 45 градусов. Нормы загрузки рекомендуются следующие: в автомашине ГАЗ-51-50-60 штук; в автомашине ЗИЛ-150-70-80 штук; в вертолете МИ-4-25-30 штук; в самолете АН-2-до30 штук; в самолете ИЛ-14-до 100 штук. Производителей лучше перевозить осенью при температуре 5-7 0 C, весьма нежелательна их транспортировка весной. На выживаемость перевозимой рыбы влияют несколько факторов, основным из которых является содержание кислорода в воде, накопление продуктов жизнедеятельности и свободное пространство. Большое значение также придается качеству и физиологическому состоянию перевозимых объектов. Вода для перевозки рыбы должна быть чистой, прозрачной, без механических и органических примесей. Очень важно, чтобы перевозимая рыба не испытывала резких колебаний температуры. Разница температуры воды, в которой рыба находилась до погрузки и воды, в которой она будет перевозится, не должна превышать 1-2 0 C, также как и при выгрузки рыбы. Личинок можно транспортировать через 2-4 суток после выклева, вскоре после заполнения плавательного пузыря воздухом. При загрузке пакетов не допускаются травмированные, слабые и не жизнестойкие личинки, их отбраковывают. Личинки плохо переносят тряску, что имеет место при перевозке их от аэропортов до рыбхозов по плохой дороге, на неприспособленном транспорте. Недопустимы во время перевозке резкие колебания температуры: повышение за пределы 300 и падение ниже 150 . Пакеты укладывают в картонные коробки размером 33x60x33см. Или 40x65x40см, предварительно выстланные хлорвиниловой пленкой, завязывают все это веревкой диаметром не менее 1см. Вес коробки 20кг. При длительной перевозке и высокой температуре воду меняют и добавляют новую порцию кислорода. Кроме пакетов, для перевозки личинок сазана используют канистры, загрузка в них в 2 раза быстрее.
При транспортировке оплодотворенной икры прежде всего стремятся создать условия, которые обеспечивают нормальный ход процессов ее обмена. Техника перевозки предусматривает также условия, предохраняющие икру от резких сотрясений и, следовательно, механических повреждений. Оплодотворенную икру сазана перевозят на дальние расстояния в специальной изотермической таре во влажной атмосфере, причем в теплый период года прибегают к различным способом охлаждения атмосферы внутри ящиков, чтобы замедлить развитие икры и уменьшить интенсивность ее газообмена. При этом следует избегать переохлаждения икры. Оплодотворенную икру лучше всего перевозить после появления у подвижных эмбрионов пигментированных глаз. Транспортировка оплодотворенной икры не представляет особых трудностей и при соблюдении необходимых биотехнических требований обычно дает хорошие результаты. Для перевозки неоплодотворенной икры ее закладывают в сухую банку, которую плотно закрывают пробкой и помещают в термос. Банка должна полностью заполнятся икрой без свободного воздушного пространства. Перевозку икры при температуре воздуха выше 7 0 C осуществляют в ящиках со льдом. Для этого рамки с икрой размещают на 10см выше дна ящика, а наверх кладут пустую рамку, которую загружают льдом. Максимальная гибель икры при транспортировке не должна превышать 5%. Доставленную на место икру помещают в инкубационные аппараты, где продолжается развитие эмбрионов.
Глава VI. Биологические основы акклиматизации
Акклиматизация – процесс приспособления переселенных в другой водоем особей вида к новым условиям среды, в результате чего из их потомства образуется популяция. Этот процесс протекает медленно и связан с глубокой перестройкой, происходящей в организме.
Акклиматизация рыб и кормовых беспозвоночных является составной частью комплексных мероприятий по воспроизводству рыбных запасов и кормовых ресурсов в водоемах.
Задачей акклиматизационных работ являются повышение продуктивности и хозяйственной ценности водоемов, улучшение видового состава фауны, а также сохранение и увеличение численности отдельных ценных видов водных организмов за счет расширения ареала.
Различают пять основных понятий акклиматизации: интродукция, вселение, зарыбление, акклиматизация, натурализация. Кроме этих основных понятий акклиматизации, часто употребляются следующие: поэтапная, реакклиматизация, аутоакклиматизация.
Сазана я решил акклиматизировать в оз. Балхаш для этого целесообразно использовать вселение – переселение особей вида в водоем, условия среды в котором мало или совершенно не отличаются от условий жизни данного вида в материнском водоеме. Вселенные особи вида успешно размножаются на новом месте обитания без какой-либо предварительной внутренней перестройки организма. Биологические особенности потомства переселенных особей вида не изменяются. При этом типом акклиматизации будет являться внедрение, которое проводится при наличии относительно свободной ниши, в которой переселенец занимает свое место, используя имеющиеся в водоеме резервы корма, и не вступает или почти не вступает в конкурентные отношения с аборигенами.
Балхаш, как и многие другие озера пустынной зоны, отчетливо приурочен к подножью горных цепей. Площадь его бассейна составляет 501000 км2 . Балхаш по своим гидрохимическим, температурным показателям является наиболее подходящим водоемом для вселения сазана.
Температурный режим
Нагрев воды начинается в марте, а наибольшим оказывается в апреле. Отдача тепла водою наблюдается в июле. Максимальное охлаждение водных масс Балхаша приходится на октябрь, а минимальное – на декабрь в период образования льда.
Осенний переломный момент в средних месячных температурах воды и воздуха наблюдается в сентябре, в то время как отстаивание воды начинается уже в июле. Ноябрь является месяцем наибольшей отдачи тепла водою, имеет наименьшую разность между средними температурами воды и воздуха. Декабрь имеет не минимальный температурный градиент между водою и воздухом. В термическом режиме Балхаша проявляется весьма сложное влияние ландшафта. Нагрев воды в марте объясняется влиянием более теплых вод южных притоков, а потеря тепла водными массами в августе – влиянием испарения. Наибольшая отдача тепла в ноябре объясняется осенней циркуляцией, охлаждением поверхностных слоев до +4 0 С и опусканию их на дно. Слабое изменение запасов тепла в декабре находится в прямой связи с выделением водою скрытой теплоты льдообразования.
Западный Балхаш имеет больший запас тепла в течение всего лета и меньший в течение всей зимы, быстрее охлаждается осенью и скорее нагревается весной, чем Восточный Балхаш. Мобильность теплового баланса Западного Балхаша нельзя объяснить влиянием температуры воздуха. Восточный Балхаш имеет более низкие месячные температуры воздуха, между тем остывание зимою его идет значительно медленнее. Несомненно, что большая мобильность теплового баланса Западного Балхаша определяется особенностями его котловины.
Среднемесячные температуры воды и воздуха в Западном и Восточном Балхаше
Таблица №6
| Месяц | Западный Балхаш | Восточный Балхаш | ||
| воздуха | воды | воздуха | воды | |
| Январь | -18,0 | 0,2 | -25,6 | 0,2 |
| Февраль | -16,7 | 0,04 | -23,6 | 0,1 |
| Март | -4,1 | 0,2 | -5,9 | 0,6 |
| Апрель | 10,7 | 8,6 | 5,2 | 4,5 |
| Май | 16,2 | 16,7 | 14,9 | 13,5 |
| Июнь | 23,9 | 21,1 | 19,8 | 19,4 |
| Июль | 26,6 | 23,8 | 21,5 | 20,1 |
| Август | 24,6 | 21,4 | 20,9 | 19,8 |
| Сентябрь | 15,4 | 15,4 | 14,2 | 14,2 |
| Октябрь | 7,4 | 8,4 | 7,3 | 7,7 |
| Ноябрь | -0,8 | 2,4 | -1,7 | 1,7 |
| Декабрь | -11,5 | 0,2 | -15,4 | 0,4 |
Итак, водные массы Западного Балхаша, имеющего среднюю глубину 4,79 м, находятся в более благоприятном положении для воздействия со стороны окружающей среды – атмосферы, дна и притоков, чем воды Восточного Балхаша, имеющего среднюю глубину 8,79 м.
Минимальные и максимальные значения температуры балхашской воды за длительный промежуток времени свидетельствует о том, что прогрев воды начинается в апреле сразу же после вскрытия озера, которое обычно наступает в 1-й декаде апреля. Но сильные ветры и вызываемое ими перемешивание водных масс в апреле могут очень сильно охлаждать воду, о чем свидетельствуют низкие минимальные температуры воды этого месяца. В течение мая и июня воды в Балхаше нагреваются еще выше, чем в апреле, а термический режим носит более устойчивый характер и не имеет резких депрессий. В дальнейшем с июня интенсивность нагрева воды начинает ослабевать. Кульминационный момент обычно наблюдается во 2-й декаде июля, когда температура в поверхностных горизонтах воды достигает до 28,2 0 С. Начиная с августа охлаждение поверхностных горизонтов воды происходит также интенсивно, как и нагрев. Со второй декады августа обычно наблюдается постепенное охлаждение озера.
В связи с большой мелководностью осеннее охлаждение в Западном Балхаше происходит быстрее, чем в Восточном. Отсюда существенная разница в датах замерзания и вскрытия между Западным и Восточным Балхашем.
Наибольшие и наименьшие средние температуры поверхности воды Балхаша (в 0 С)
Таблица №7
| Месяц | Температура | |
| максимальная | минимальная | |
| Январь | 0,56 | 0,1 |
| Февраль | 0,54 | 0,0 |
| Март | 1,27 | 0,0 |
| Апрель | 14,4 | 0,8 |
| Май | 18,5 | 11,1 |
| Июнь | 24,3 | 16,0 |
| Июль | 28,2 | 18,6 |
| Август | 23,9 | 17,7 |
| Сентябрь | 18,5 | 8,3 |
| Октябрь | 17,1 | 3,2 |
| Ноябрь | 2,7 | 0,0 |
| Декабрь | 1,0 | 0,0 |
| Среднегодовая | 12,5 | 6,3 |
Особенности термического режима заставляют отнести Балхаш к однородным тепловодным озерам умеренной зоны.
Соленость
Главную роль в химизме озерных вод аридной зоны играют Са(НСО3 )2 , Mg(НСО3 )2 , Na2 SO4 , MgSO4 и NaCl. Поскольку основной особенностью пустынной зоны является преобладание испарения над осадками, каждое озеро пустынной области имеет отрицательный водный баланс, если его рассматривать за достаточно длительный геологический промежуток времени. Современный Балхаш имеет отрицательный с геологической точки зрения баланс влаги.
Разные части озера имеют различную соленость. Так, Западный Балхаш, является пресным или почти пресным, а Восточный Балхаш обладает высокой соленостью.
Балхашская вода имеет постоянное содержание СаСО3 . В балхашской воде не содержится СаСl2 , ни МgCl2 , но имеется Na2 SO4 . Это озеро, по классификации Н. С. Курнакова, относят к озерам сульфатного типа.
Углекислый газ присутствует только в западной половине Балхаша, а в восточной половине озера его нет. Отсутствие СО2 приводит к тому, что выпавшие в осадок карбонаты не могут переходить в бикарбонаты и остаются на дне.
Важным опресняющим фактором для Балхаша является испарение воды в его заливах. Озеро может повышать свою соленость ежегодно на 0,04‰. Балхаш получает до 80% воды в западном конце, от чего вода его по мере движения на восток приобретает все большую и большую соленость. И помимо этого изменениям концентрации солей дело не ограничивается. Вода в восточной части Балхаша по своему составу не тождественна воде его западной части. Под влиянием испарения и концентрирования происходит выпадение на дно одних солей и в результате этого относительное обогащение воды другими. Отмирающие моллюски и другие организмы не обогащают воду Балхаша кальцием. Грунтовые воды Балхаша иногда приносят Nа2 СО3 .
Питание сазана в озере Балхаш
Можно составить представление о сезонной динамике зоопланктона и его групп. Летняя плотность зоопланктона является максимальной, осенне-зимняя – минимальной, а весенняя занимает промежуточное положение. Очень неравномерна плотность распределения зоопланктона наблюдается в более опресненном западном Балхаше. Сырой объем планктона в Балхаше составляет 16,6 г/м3 . Средняя биомасса его в Балхаше 3,34 г/м3 или 21,07 кг/га. Чем больше глубина, тем выше биомасса зоопланктона.
По отношению к солености воды водоросли распределяются на солоноватоводные – 53 (около 17%), солоноватоводно-опресненные 140 (45%), пресноводные – 115 (38%).
В Балхаше главная масса донных и планктонных водорослей состоит из пресноводной флоры. В целом в видовом отношении флора водорослей Балхаша бедна.
Кормовая база озера Балхаш не будет препятствовать существованию в нем сазана. В связи с бедностью донной области основной пищей, по-видимому, будет являться детрит с примесью личинок хирономид и олигохет, зоопланктон – (в основном веслоногие ракообразные), растительная пища, которая будет преобладать над животной.
Газовый режим
Благодаря мелководности частому ветровому перемешиванию всей толщи воды и невысокой окисляемости кислородный режим озера в летний период устойчивый. Изменений между западной и восточной частями озера нет. Но все же есть некоторые. В летний период более мелководный западный Балхаш имеет несколько более насыщению воды кислородом. Летний кислородный минимум в придонных слоях в августе не опускается ниже 3,11 мг/л О2 .
Другим важным элементом газового режима вод Балхаша является содержание в них двуокиси углерода. Двуокись углерода присутствует лишь там, где рН воды не превышает 8,4. При рН от 8,2 до 8,4 лишь следы СО2 .
План акклиматизации.
1. Выбор района вселения в озеро Балхаш.
2. Вселение объекта конец весны начало лета на стадии малька.
3. Освоение территории в течение одного года.
4. Первое поколение через 3-5 лет.
5. Фаза взрыва через 7 лет.
6. Натурализация через 10 лет.
Список используемой литературы
1. Абросов В.Н. Озеро Балхаш, 1983.
2. Архангельский В. В., Васильева Л. М. Перспективы пастбищной аквакультуры ценных видов рыб на водоемах ильменного типа дельты Волги. Журнал «Рыбное хозяйство», 2003, №6, стр.44-45.
3. Аскеров Т.А. Некоторые данные о нерест. инстинкте сазана при гипофиз. инъекциях. В сб: Современное сост-е метода гипоф. инъекций. Астрахань, 1969. С.22-24.
4. Берлянц Т.Б., Лысикова К.В., Тамарин А.Р. Перевозка икры карповых рыб на дальние расстояния. (Борьба с наруш. в развит. эмбрионов карповых рыб при инкубации и перевозке икры). М. Мин-во рыбной пром. 1956. стр.28.
5. Виноградов В.К. Биологические основы разведения и выращивания растит. рыб и новых объектов рыбов. и акклимат. Дисс. докт. биол. наук в форме доклада – М:1985.-60с. (ВНИИПРХ).
6. Гамыгин Е.А., Турецкий В.И. Сборник научных трудов. Вопросы разработки и качества комбикормов. М.: ВНИИПРХ. – 1989 – Вып. 57. – 156 стр.
7. Гриншпун О.Я., Гудыша Б.И., Васюта В.В. Реакция организма карпа на введение гип. инъекции – Рыб. хоз-во, 1983. №8. С.36-37
8. Зайцев Р.А. Способ перевозки производителей и личинок растит. рыб.-Рыбное хоз-во, 1969, №11. С.16-17
9. Иванов А.П. Рыбоводство в естественных водоемах. – М.: Агропромиздат, 1988. – 367 стр.
10. Моисеев П.А., Азизова Н. А., Куранова И. И. Ихтиология. – М.: Легкая и пищевая промышленность, 1981. – 384стр.
11. Мотенков Ю. Перевозка растит. рыб в полиэтил. пакетах Белый толстолобик в бассейне. Рыбовод. и рыболовство, 1965. №2. Стр.8
12. Никольский Г.В. Экология рыб. – М.: Высшая школа, 1974. – 366 стр.
13. Пономарев С.В., Гамыгин Е.А., Никоноров С.И., Пономарева Е.Н., Грозеску Ю.Н., Бахарева А.А. Технологии выращивания и кормления объектов аквакультуры юга России. (Справочное учебное пособие). – Астрахань: «Нова плюс», 2002. – 264 стр.
14. Федяев В.Е. Прудовое рыбоводство страны: Прошлое, настоящее, будущее. Журнал «Рыбное хозяйство», 2003, №1, стр. 38-39.