| Похожие рефераты | Скачать .docx | Скачать .pdf |
Реферат: Основы биохимии
СОДЕРЖАНИЕ
Вопрос 10. Приведите примеры биологически активных пептидов. Какие функции они выполняют?
Вопрос 20. Как связана простетическая группа фосфопротеинов с белковым компонентом. Показать ее на примере аминокислот серина и треонина
Вопрос 30. Энзинопатологии (фенилкетонурия, альбинизм и алкаптонурия)
Вопрос 40. Назвать коферментные формы витаминов биотина и В12, указать в каких обменных процессах они участвуют
Вопрос 46. Напишите формулу сиаловой кислоты. Охарактеризуйте расположением гликопротеинов в биомембране. Какую функцию они выполняют?
Вопрос 50. Написать реакцию НАД-зависимого дгидрирования малата в цикле КРЕБСА
Вопрос 52. Что такое окислительное фосфорелирование? Указать участки сопряжения окисления и фосфорелирования в цепях переноса электронов
Вопрос 64. Напишите фрагменты молекул клетчатки, крахмала. Укажите какой углевод не переваривается в ЖКТ человека и почему?
Вопрос 75. Написать в виде схемы реакции синтеза гликогена
Вопрос 84. Какие вещества необходимы для синтеза жирных кислот на синтаве жирных кислот. Как связан процесс синтеза жирных кислот с обменом углеводов?
Вопрос 88. Какими липопротеинами крови доставляются жиры из печени и кишечника к жировым депо? Каков состав этих липопротеинов?
Вопрос 96. В состав пищи входили пептиды следующего состава: ала-сер-гли-дис -сен-лиз-три-вал-лей. Назовите ферменты и соответствующие связи, которые они расщепляют: а) в желудке, б) в просвете тонкой кишки,
в) в слизистой оболочке тонкой кишки
Вопрос 106. Декарбоксилирование аминокислот. Строение кофермента
декарбоксилаз. Биологическая роль образовавшихся соединений
Вопрос 108. Напишите орнитиновый цикл мочевинообразования. Укажите локализацию процесса
Вопрос 121. Через какие активные соединения клетки гормоны реализуют свое действие?.
Вопрос 126. Охарактеризуйте механизм действия адреналина и глюкагона на липидный обмен
Вопрос 130. Клинико-диагностическое значение определения активности амилазы сыворотки крови и мочи
Вопрос 133. Нарушение прямого и соотношения непрямого билирубина при желтухах. Охарактеризуйте гемолитическую, паренсиматозную и обтурационную желтухи
Вопрос 134. С какими свойствами белков плазмы крови связана их
способность регулировать объем крови и ее онкотическое давление? Какая белковая фракция играет наиболее существенную роль в обеспечении этой функции?
Вопрос 143. Приведите схему строения антител. Обозначьте на схеме участки взаимодействия с антигеном
Список используемой литературы
Вопрос 10. Приведите примеры биологически активных пептидов. Какие функции они выполняют?
Ответ. В последние годы значительно повысился интерес к структуре и функциям встречающихся в свободном состоянии в организме низкомолекулярных пептидов, выполняющих ряд специфических биологических функций. Короткие пептиды, содержащие до 10 аминокислот, принято называть олигопептидами; в то же время полипептиды и белки считаются взаимозаменяемыми, хотя термином «полипептиды» чаще обозначают продукты с мол. м. менее 10000. В некоторых биоактивных пептидах имеются необычные аминокислоты, не встречающиеся в природных белках, или производные обычных аминокислот (гормоны, антибиотики). Мнение о том, что пептиды могут играть роль промежуточных продуктов на пути синтеза белка, не подтвердилось, поскольку этот процесс во всех клетках у всех живых организмов осуществляется de novo матричным путем.
Природные пептиды, наделенные биологической активностью, в зависимости от характера действия и происхождения принято делить на 4 группы: 1) пептиды, обладающие гормональной активностью (вазопрессин, окси-тоцин, кортикотропин, глюкагон, кальцитонин, меланоцитстимулирующий гормон, рилизинг-факторы гипоталамуса и др.; 2) пептиды, принимающие участие в процессе пищеварения (в частности, гастрин и секретин); 3) пептиды, источник которых – α2-глобулиновая фракция сыворотки крови (такие, как ангиотензин, брадикинин и каллидин); 4) нейропептиды.
В последнее время выяснены некоторые закономерности синтеза физиологически активных пептидов из биологически инертных предшественников – белков в результате процесса, называемого посттрансляционной модификацией (постсинтетические превращения белковой молекулы). Известно, например, что ангиотензины (представленные октапептидами), оказывающие выраженное сосудосуживающее действие, образуются из присутствующего в сыворотке крови неактивного белка ангиотензиногена в результате последовательного действия ряда протеолитических ферментов (ренина и особого фермента, участвующего в превращении неактивного ангиотензина I в активный ангиотензин II).
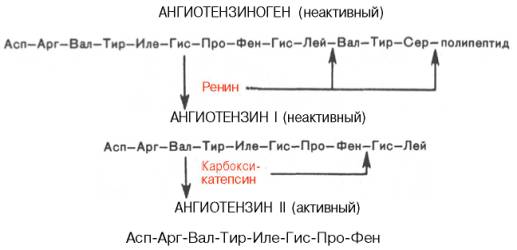
К группе вазоактивных (оказывающих влияние на тонус сосудов) пептидов относятся, кроме того, широко применяемые в медицинской практике брадикинин и каллидин.
Брадикинин представляет собой нонапептид:
H–Apг–Про–Про–Гли–Фен–Сер–Про–Фен–Apг–ОН.
Каллидин представлен декапептидом, образующимся из неактивного плазменного белка кининогена, и отличается от брадикинина присутствием на N-конце еще одного аминокислотного остатка (Лиз):
Н–Лиз–Арг–Про–Про–Гли–Фен–Сер–Про–Фен–Apг–ОН.
Совсем недавно из экстрактов ткани предсердия (но не из желудочков сердца) человека и животных были выделены биологически активные пептиды, регулирующие тонус сосудистой системы и электролитный обмен. Физиологический эффект их оказался противоположным влиянию системы ренин–ангиотензин–альдостерон. Он выражается в сосудорасширяющем действии, усилении клубочковой фильтрации и стимуляции выведения натрия и хлоридов за счет угнетения их реабсорбции в канальцах. Эти пептиды получили название а триопептидов (от лат. atrio – предсердие). Они построены из разного числа аминокислот (от 23 до 100), но обязательным условием для проявления биологического эффекта является наличие в молекуле 17-членной кольцевой структуры, образующейся за счет дисуль-фидной связи между остатками цистеина.
Внутриклеточным посредником действия атриопептидов оказался циклический гуанозинмонофосфат (цГМФ), синтез которого осуществляется в результате активирования мембранного фермента гуанилатциклазы; действие аденилатциклазы, напротив, тормозится под влиянием атриопеп-тидов.
Во всех животных тканях и в некоторых растениях широко распространен низкомолекулярный трипептид глутатион, функции которого пока не выяснены достаточно полно, хотя он открыт сравнительно давно. Глута-тион представляет собой атипичный трипептид (в котором в образовании одной из пептидных связей участвует не α-карбоксильная, а γ-карбок-сильная группа глутамата) следующего строения: γ-глутамил-цистеинил-глицин:
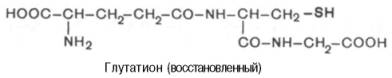
Цистеин является составной частью глутатиона, поэтому последний может находиться в восстановленной (SH) и в окисленной (S-S) формах (сокращенно обозначаются Г-SH и Г-S-S-Г), что, по-видимому, имеет отношение к биологической роли глутатиона в организме.
Интерес к природным пептидам в значительной степени обусловлен необычно высокой их биологической активностью. Они оказывают мощное фармакологическое действие на множество физиологических функций организма. В то же время были замечены низкая стабильность и быстрый распад их в организме при физиологических значениях рН среды. Все это способствовало развитию исследований как в области препаративного выделения природных пептидов из органов и тканей (включая получение биологически активных пептидов из предшественников методами ограниченного протеолиза ряда хорошо известных гормонов), так и в области химического синтеза. Получение ряда биологически активных нейропепти-дов из гормонов гипофиза, в частности эндорфинов и энкефалинов, наделенных мощным обезболивающим действием (путем связывания рецепторов определенных клеток мозга), в сотни и тысячи раз превосходящим аналгезирующий эффект морфина.
Из ткани мозга выделен также δ-пептид сна; ряд других нейропептидов принимает участие в биохимических механизмах памяти, страха, обучения и т.д. Для повышения стабильности пептидов при введении в организм предприняты попытки химического синтеза пептидов, в которых один или несколько аминокислотных остатков L-ряда замещают остатками D-аминокислот. Подобная замена, не вызывая снижения биоактивности, защищает пептид от воздействия протеиназ тканей, способствуя пролонгированию эффекта препарата.
Среди естественно встречающихся небольших пептидов следует указать на антибиотик грамицидин S, выделенный из Bacillus brevis и представляющий собой циклический декапептид:
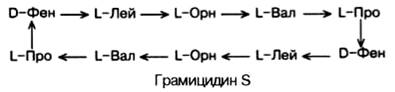
Как видно, в структуре грамицидина S имеются 2 остатка орнитина (Орн), производные аминокислоты аргинина и 2 остатка неприродных D-изомеров фенилаланина. Стрелки указывают направление синтеза от NН2-групп к СООН-группам каждого остатка, и вследствие цикличности грамицидин S не имеет конца.
Широкое применение, особенно в пищевой промышленности, в качестве заменителя сахара получил искусственный (генноинженерный синтез) ди-пептид, состоящий из L-изомеров аспарагиновой кислоты и метилового эфира фенилаланина, названный аспартамом:
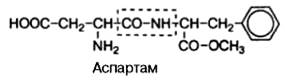
Аспартам в сотни раз слаще сахара и легко распадается в организме на две свободные аминокислоты, абсолютно безвредные для организма; поэтому он рекомендован в качестве заменителя сахара больным диабетом. Это пример пептида, наделенного огромным биологическим эффектом.
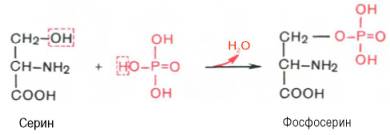
Вопрос 20. Как связана простетическая группа фосфопротеинов с белковым компонентом. Показать ее на примере аминокислот серина и треонина
Ответ. К белкам этого класса относятся казеиноген молока, в котором содержание фосфорной кислоты достигает 1%; вителлин, вителлинин и фосвитин, выделенные из желтка куриного яйца; овальбумин, открытый в белке куриного яйца; ихтулин, содержащийся в икре рыб, и др. Большое количество фосфопротеинов содержится в клетках ЦНС. Фосфопротеины занимают особое положение в биохимии фосфорсодержащих соединений не только в результате своеобразия структурной организации, но и вследствие широкого диапазона функций в метаболизме. Характерной особенностью структуры фосфопротеинов является то, что фосфорная кислота оказывается связанной сложноэфирной связью с белковой молекулой через гидроксильные группы β-оксиаминокислот, главным образом серина и в меньшей степени треонина. На одну молекулу белка обычно приходится 2–4 остатка фосфата.
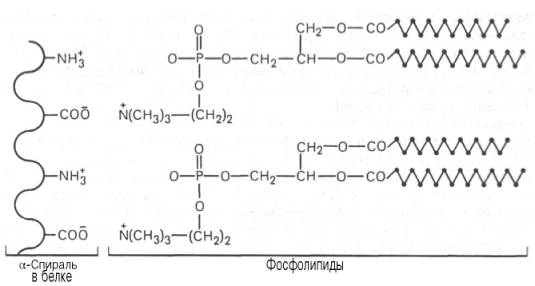
Рис. 1. Ионный тип связи между белками и фосфолипидами.
Новые данные свидетельствуют о том, что в клетках фосфопротеины синтезируются в результате посттрансляционной модификации, подвергаясь фосфорилированию при участии протеинкиназ. Таким образом, уровень фосфопротеинов в клетке зависит в значительной степени от регулирующего действия ферментов, катализирующих фосфорилирование (протеин-киназы) и дефосфорилирование (протеинфосфатазы). Следует отметить, что фосфопротеины содержат органически связанный, лабильный фосфат, абсолютно необходимый для выполнения клеткой ряда биологических функций. Кроме того, они являются ценным источником энергетического и пластического материала в процессе эмбриогенеза и дальнейшего постна-тального роста и развития организма.
Особо следует отметить, что некоторые ключевые ферменты, регулирующие процессы внутриклеточного обмена веществ, также существуют как в фосфорилированной, так и в дефосфорилированной форме. Этим подчеркивается значение фосфорилирования–дефосфорилирования в процессах химической модификации макромолекул, участвующих в интегральных процессах метаболизма.
Вопрос 30. Энзинопатологии (фенилкетонурия, альбинизм и алкаптонурия)
Ответ. Фенилкетонурия , фенилпировиноградная олигофрения, наследственное заболевание из группы ферментопатий, в основе которого лежит аномалия аминокислотного обмена вследствие отсутствия или резкого снижения активности фермента фенилаланингидроксилазы. Описана в 1934 норв. учёным А. Фёллингом (A. Foiling) (болезнь Фёллинга). Частота фенилкетонурии – 1 случай на 10–15 тыс. новорождённых; наследуется по аутосомно-рецессивному типу. При фенилкетонурии фенилаланингидроксилаза сохраняет только около 5% активности, в связи с чем нарушается обмен фенилаланина и вследствие этого – тирозина, триптофана и др., накапливаются промежуточные продукты обмена – фенилэтиламин, фенилпировиноградная кислота и др. и возникает дефицит метаболитов, необходимых для нормального функционирования организма. В частности, вероятная причина умственных расстройств – дефицит медиаторов нервной системы (адреналина, норадреналина, серотонина и др.). Таким образом, при фенилкетонурии возникает комплекс взаимосвязанных метаболических расстройств, состоящий из первичного ферментного нарушения и обусловленных им др. нарушений обмена.
Фенилкетонурия проявляется главным образом выраженной олигофренией (идиотией или имбецильностью). Диагностируется в первые дни жизни ребёнка с помощью экспресс-методов – микробиологических или биохимических. Последние основаны на определении пировиноградной кислоты в моче посредством индикаторов (проба Фёллинга). Лечение сводится главным образом к специальной диете (резкое ограничение продуктов, содержащих фенилаланин).
Альбинизм (albinismus) представляет собой врожденное отсутствие кожного пигмента. Этиология и патогенез изучены недостаточно. Известно, что в результате нарушения синтеза ферментов тирозиназы, дофаоксидазы прекращается образование меланина из тирозина, диоксифенилаланина. О сложности механизмов происхождения альбинизма свидетельствуют нередко сопутствующие ему такие аномалии, как врожденная глухота, дефекты интеллекта, патология глаз и другие.
Выделяют тотальный, неполный и частичный альбинизм.
Тотальный альбинизм наследуется аутосомно-рецессивно со средней частотой 1: 10000-20000. Предполагают, что носители мутантного гена составляют 1,5% среди всех нормально пигментированных людей.
Депигментация кожи и придатков наблюдается с рождения, сопровождается сухостью кожи, нарушением потоотделения, иногда гипо- или гипертрихозом, особенно на открытых участках. У больных легко возникают солнечные ожоги, актинический хейлит. Они предрасположены к развитию кератом, эпителиом, телеангиэктазий. Из-за отсутствия пигмента в тканях глаза зрачки кажутся красными. Характерными являются горизонтальный нистагм и выраженная светобоязнь. Часто наблюдаются сходящееся косоглазие, снижение остроты зрения в результате нарушений рефракции, катаракты, возможна микрофтальмия. Нередко наблюдаются бесплодие, иммунодефицит (отсюда частые инфекции), пороки развития, сокращение продолжительности жизни, олигофрения.
Неполный альбинизм (син.: альбиноидизм), в отличие от предыдущей формы, наследуется аутосомно-доминантно, в некоторых случаях — рецессивно. Имеет место снижение активности тирозиназы, но не блокада ее синтеза. Наблюдается гипопигментация кожи, волос, радужки, иногда фотофобия. Других дефектов и аномалий не регистрируется.
Частичный альбинизм (син.: пиебалдизм) наследуется аутосомно-доминантно. Проявления выявляются при рождении. Характеризуется появлением участков ахромии на коже живота, лица, нижних конечностей, прядей седых волос. Депигментированные пятна неправильной формы с резкими границами, на их поверхности имеются мелкие темно-коричневые пятнышки. Вокруг ахромичных пятен кожа может быть пигментированной. Поражений других органов обычно не бывает. Частичный альбинизм является одним из проявлений синдромов Чедиака-Хигаси, Клейна-Ваарденбурга, Титце, Менде, Хермански-Пудлака, Кросса-МакКьюзика-Брина.
Дифференциальный диагноз альбинизма проводят с витилиго, синдромами Клейна-Ваарденбурга, Алеззандрини, Фогта-Коянаги-Харады.
Алкаптонурия (alcaptomiria; алкаптон (гомогентизиновая кислота) + греч. uron моч ) - наследственная болезнь, обусловленная нарушением обмена тирозина вследствие пониженной активности фермента гомогентизиназы и накоплением в тканях организма гомотентизиновой кислоты; проявляется у взрослых пигментацией различных тканей, развитием артрозов, а у детей - лишь темным окрашиванием мочи и иногда ушной серы; наследуется по аутосомно-рецессивному типу.
Вопрос 40. Назвать коферментные формы витаминов биотина и В12, указать в каких обменных процессах они участвуют
Ответ. Биологическая роль. Биотин подробно изучен благодаря работам Ф. Линена. Известные к настоящему времени биотиновые ферменты (т.е. ферменты, содержащие в качестве кофермента биотин) катализируют два типа реакций:
1) реакции карбоксилирования (с участием СО2 или НСО3–), сопряженные с распадом АТФ
RH + HC03– + ATO<=> R-COOH +AflO+H3P04;
2) реакции транскарбоксилирования (протекающие без участия АТФ), при которых субстраты обмениваются карбоксильной группой
R1-COOH + R2H <=> R1H + R2-COOH.
Получены доказательства двустадийного механизма этих реакций с образованием промежуточного комплекса (карбоксибиотинилфермент).
К реакциям первого типа относятся, например, ацетил-КоА- и пируват-карбоксилазные реакции:
C H 3– C O – S - K o A + CO2+ АТФ <=> H O O C – C H 2– C O – K o A + АДФ + Pi.
Пируваткарбоксилаза является высокоспецифичным ферментом, катализирующим уникальную реакцию усвоения СО2 в организме животных. Сущность реакции сводится к пополнению запасов оксалоацетата (щаве-левоуксусная кислота) в лимоннокислом цикле (так называемые «анаплеро-тические», «пополняющие» реакции), т.е. его синтезу из СО2 и пирувата:
Пируват + CO2+ АТФ + H2O —> Оксалоацетат + АДФ + Pi+ 2H+
Реакция протекает в две стадии: на первой стадии, связанной с затратой энергии, СО2 подвергается активированию, т.е. ковалентному связыванию с биотином в активном центре фермента (Е-биотин):
![]()
На второй стадии СО2 из комплекса переносится на пируват с образованием оксалоацетата и освобождением фермента:
![]()
Примером второго типа реакций является метилмалонил-оксалоаце-тат-транскарбоксилазная реакция, катализирующая обратимое превращение пировиноградной и щавелевоуксусной кислот:
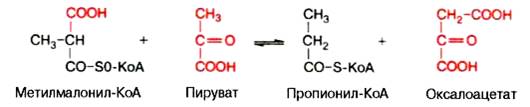
Реакции карбоксилирования и транскарбоксилирования имеют важное значение в организме при синтезе высших жирных кислот, белков, пури-новых нуклеотидов (соответственно нуклеиновых кислот) и др.
Распространение в природе и суточная потребность. Биотин содержится почти во всех продуктах животного и растительного происхождения, главным образом в связанной форме. Богаты этим витамином печень, почки, молоко, желток яйца. В растительных продуктах (картофель, лук, томат, шпинат) биотин находится как в свободном, гак и в связанном состоянии. Для человека и животных важным источником является биотин, синтезируемый микрофлорой кишечника. Суточная потребность взрослого человека в биотине приблизительно 0,25 мг.
Биологическая роль. Выявлены ферментные системы, в составе которых в качестве простетической группы участвуют не свободный витамин В12, а так называемые В12-коферменты, или кобамидные коферменты. Последние отличаются тем, что содержат 2 типа лигандов: метильную группу и 5'-дезоксиаденозин. Соответственно различают метилкобаламин СН3-В12 и дезоксиаденозилкобаламин. Превращение свободного витамина В12 в В12-коферменты, протекающее в несколько этапов, осуществляется в организме при участии специфических ферментов в присутствии в качестве кофакторов ФАД, восстановленного НАД, АТФ и глутатиона. В частности, при образовании 5-дезоксикобаламинового кофермента АТФ подвергается необычному распаду с отщеплением три-фосфатного остатка по аналогии еще с одной единственной реакцией синтеза 5-аденозилметионина из метионина и АТФ. Впервые В12-коферменты были выделены Г. Баркером и сотр. в 1958 г. из микроорганизмов, позже было доказано их существование в тканях животных.
Химические реакции, в которых витамин В12 принимает участие как кофермент, условно делят на 2 группы в соответствии с его химической природой. К первой группе относятся реакции трансметилирования, в которых метилкобаламин выполняет роль промежуточного переносчика метильной группы (реакции синтеза метионина и ацетата).
Синтез метионина требует, помимо гомоцистеина, наличия N5-метил-ТГФК и восстановленного ФАД и протекает согласно уравнению:

Фермент, катализирующий эту реакцию, был открыт в печени человека и ряда животных, а также у микроорганизмов. Получены доказательства, что механизм реакции включает перенос метильной группы N5-СН3-ТГФК на активный центр фермента с образованием метил-В12-фермента и последующий перенос этой группы на гомоцистеин. Блокирование этой реакции, наблюдаемое при авитаминозе В12, приводит к накоплению N5-СН3-ТГФК и соответственно выключению из сферы химических реакций еще одного важного кофермента.
Вторая группа реакций при участии В12-коферментов заключается во внутримолекулярном переносе водорода в реакциях изомеризации. Механизм этих реакций соответствует схеме:

Видно, что протон водорода движется (перемещается) между двумя соседними атомами углерода и не обменивается с протонами воды. Предполагают, что сначала водород от субстрата переносится на 5-дезокси-кобаламин, а затем обратно на субстрат, меняя местоположение. Например, глутаматмутазная реакция (взаимопревращения глутаминовой и β-метиласпарагиновой кислот), метилмалонилмутазная реакция (обратимое превращение метилмалонил-КоА в сукцинил-КоА), глицерол- и диол-дегидратазные реакции, ферментативные реакции восстановления рибо-нуклеотидов до дезоксирибонуклеотидов и др. В организме человека из указанных процессов открыта только реакция изомеризации метил-малонил-КоА в сукцинил-КоА.
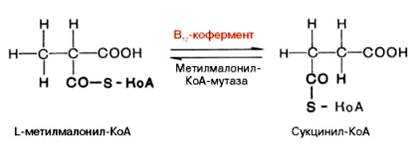
Следует подчеркнуть, что реакция изомеризации метилмалонил-КоА требует наличия 5'-дезоксиаденозилкобаламина в качестве кофермента, в то время как реакция метилирования нуждается в метилкобала-мине. Этими обстоятельствами могут быть объяснены некоторые биохимические симптомы недостаточности витамина В12, в частности метил-малонилацидурия и гомоцистинурия. Кроме того, описаны болезни, обусловленные наследственными дефектами синтеза только дезоксиаденозил-кобаламина или обоих В12-коферментов; в этих случаях даже 1000-кратная доза витамина В12 не оказывала лечебного эффекта. В настоящее время высказывается предположение о более широком участии В12-коферментов в ферментативных реакциях трансметилирования, дезаминирования (например, этаноламиддезаминазная реакция) и др. Предстоит, однако, приложить немало усилий, чтобы выяснить молекулярные механизмы действия витамина В12 на процесс кроветворения. Положительный эффект при лечении пернициозной анемии полусырой печенью обусловлен, как стало известно, наличием витамина В12, хотя следует указать, что большего лечебного эффекта можно добиться при одновременном введении внутреннего фактора слизистой оболочки желудка.
Распространение в природе и суточная потребность. Витамин В12 является единственным витамином, синтез которого осуществляется исключительно микроорганизмами; ни растения, ни ткани животных этой способностью не наделены. Основные источники витамина В12 для человека – мясо, говяжья печень, почки, рыба, молоко, яйца. Главным местом накопления витамина В12 в организме человека является печень, в которой содержится до нескольких миллиграммов витамина. В печень он поступает с животной пищей, в частности с мясом, или синтезируется микрофлорой кишечника при условии доставки с пищей кобальта. Суточная потребность в витамине В12 для взрослого человека составляет около 3 мкг (0,003 мг).
Вопрос 46. Напишите формулу сиаловой кислоты. Охарактеризуйте расположением гликопротеинов в биомембране. Какую функцию они выполняют?
Ответ. Гликопротеины – сложные белки, содержащие, помимо простого белка или пептида, группу гетероолигосахаридов. В настоящее время их принято называть гликоконъюгатами. В состав гликоконъюгата входит углеводный компонент (гликановая фракция), ковалентно связанный с неуглеводной частью (агликановая фракция), представленной белком, пептидом, аминокислотой или липидом.
Повышенный интерес к науке об углеводах – гликобиологии – в настоящее время объясняется открытием существенной роли изменений структуры гликоконъюгатов в развитии таких болезней, как рак, иммунодефицит человека, ревматоидные артриты, астма и др. Оказалось, что нарушение реакции гликозилирования двух главных классов глико-конъюгатов (гликопротеинов и ганглиозидов) приводит или к накоплению предшественников этих веществ, или к синтезу «укороченных» сахарных цепей гликоконъюгатов. Более того, установлено, что во взаимодействии между некоторыми вирусами и клетками-мишенями главную роль играют углеводные компоненты. В частности, гликопротеин gp120 вируса иммунодефицита человека (содержит большой процент углевода) имеет высокое сродство к гликопротеину CD4Т-лимфоцита. В этом взаимодействии, узнавании, являющемся высокоспецифичным, гликозилированные фрагменты, вероятнее всего, играют важную патогенетическую роль. Известно также, что при ревматоидных артритах часто синтезируются аномальные антитела (аномальные иммуноглобулины – все гликопротеины) с необычайно короткими сахарными цепями, что вызывает стимуляцию иммунной системы против самого организма. Из этих примеров видно, что, помимо гликобиологии, наступило время признания и таких наук, как гликопа-тология и гликотерапия.
Помимо гликопротеинов, различают также протеогликаны, состоящие из белка и гликозаминогликанов (прежнее название мукополиса-хариды); последние состоят из цепей сложных углеводов: аминосахаров, уроновых кислот, серной кислоты и отдельных моносахаридов. Типичными гликозаминогликанами являются гиалуроновая кислота, хондроитинсерная кислота и гепарин.
К типичным гликопротеинам относят большинство белковых гормонов, секретируемые в жидкие среды организма вещества, мембранные сложные белки, все антитела (иммуноглобулины), белки плазмы крови, молока, овальбумин, интерфероны, факторы комплемента, группы крови, рецеп-торные белки и др. Из этого далеко не полного перечня гликопротеинов видно, что все они выполняют специфические функции: обеспечивают клеточную адгезию, молекулярное и клеточное узнавание, антигенную активность опухолевых клеток, оказывают защитное и гормональное, а также антивирусное действие.
Химический состав гликопротеинов более или менее установлен, структура определена только у ряда из них. К полипептиду присоединяются гетероолигосахаридные цепи, содержащие от 2 до 10, реже 15 мономерных остатков гексоз (галактоза и манноза, реже глюкоза), пентоз (ксилоза, арабиноза) и конечный углевод, чаще всего представленный N-ацетилга-лактозамином, L-фукозой или сиаловой кислотой; в отличие от про-теогликанов гликопротеины не содержат уроновых кислот и серной кислоты.
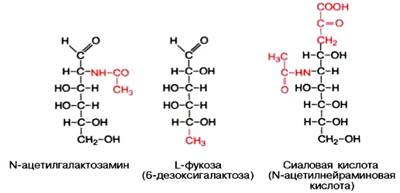
Типы связей между углеводными компонентами и белками определены только у ряда гликопротеинов, аминокислотный состав и структура которых известны (иммуноглобулины, гормоны); они включают О-гликозидные связи (с ОН-группами серина, треонина и оксилизина), N-гликозидные связи (с амидными группами аспарагина, реже глутамина или ω-NH2-группами лизина и аргинина) и эфирные гликозидные связи со свободными СООН-группами глутаминовой и аспарагиновой кислот.
Синтез гликопротеинов осуществляется в рибосомах эндоплазматиче-ского ретикулума (в цистронах), затем присоединяются сахарные цепи (постсинтетическое гликозилирование), и далее белок транспортируется до биомембран клетки и включается в состав мембранных белков или секре-тируется.
Углеводные компоненты соединены ковалентно с азотом аспарагина молекулы белка. Однако предварительно олигосахаридная часть соединяется с липидным переносчиком – долихолфосфатом (липид, содержащий от 15 до 20 изопреновых остатков) и переносится на полипептидную цепь в эндоплазматическом ретикулуме, при этом транспортер освобождается:

Долихолфосфат (n = 15-30)
Синтезированные гликопротеины далее переносятся в аппарат Гольджи, где осуществляются окончательное гликозилирование и сортировка по назначению.
Структура одного из нескольких гетероолигосахаридных остатков в молекуле гликопротеинов, в частности иммуноглобулинов, может быть представлена в виде следующей схемы (использованы сокращения: Глю – глюкоза, NАцГлюА – N-ацетилглюкозамин; Гал – галактоза; Ман – манно-за; NАцНейр – N-ацетилнейраминовая кислота):
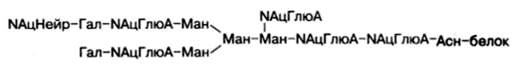
Рассмотрим известные к настоящему времени данные о синтезе, строении (структуре) и свойствах ряда гликопротеинов.
Интерфероны. Интерфероны – это ингибиторы размножения многих типов вирусов. Открыто несколько типов интерферонов (α, β и γ), некоторые из них получены методами генетической инженерии. Это сравнительно небольшие сложные белки с мол. массой у разных видов животных и человека от 25000 до 38000–40000). Они образуются в клетке в ответ на внедрение вирусной нуклеиновой кислоты, ограничивая вирусную агрессию (инфекцию). Известно также, что группа видоспецифических α-интерфе-ронов синтезируется макрофагами, в то время как γ-интерферон продуцируется Т-клетками и стимулируется интерлейкином-2. Показано также, что γ-интерферон в свою очередь повышает цитотоксическую активность макрофагов, Т-клеток и естественных клеток-киллеров. Интерфероны наделены антипролиферативной активностью и считаются основными защитными белками не только против вирусной инфекции, но и при опухолевых поражениях.
Следует отметить, однако, что до сих пор не раскрыты молекулярные механизмы, при помощи которых интерфероны тормозят размножение вирусов. Известно только, что интерфероны ингибируют биосинтез всех белков (и хозяйских, и вирусных), вероятнее всего, на уровне процесса трансляции. Возможно, что интерферон индуцирует синтез особого белка-ингибитора, который затем связывается с рибосомами и блокирует трансляцию, или интерферон переводит один из активных эукариотических белковых факторов инициации в неактивный фактор путем фосфорилиро-вания.
Иммуноглобулины. Иммуноглобулины, или антитела, также относятся к классу гликопротеинов, выполняют защитную функцию, обезвреживая поступающие в организм чужеродные вещества – антигены любой химической природы. Синтезируются иммуноглобулины плазматическими клетками, образовавшимися из лимфоцитов. Учение об иммунитете оформилось в самостоятельную науку – иммунологию, изучающую структуру и функции антител вообще и иммуноглобулинов в частности. Мы представим современные сведения о некоторых физико-химических свойствах и структуре иммуноглобулинов человека (табл. 2.4). Различают 5 классов иммуноглобулинов: IgG, IgM, IgA, IgD и IgE. Детально изучены структура и функция IgG.
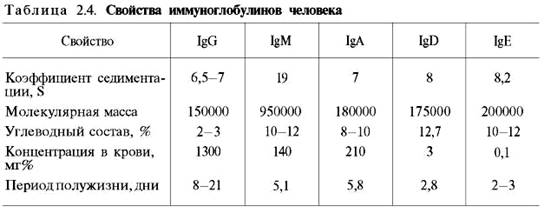
Разные классы иммуноглобулинов сильно различаются не только по молекулярной массе, но и по концентрации в крови; имеются данные, что различаются они и по биологическим свойствам.
Подробно изучена структура IgG. Он имеет Y-образную форму и тет-рамерное строение; состоит из двух идентичных легких L-цепей (от англ. light) и двух идентичных тяжелых Н-цепей (от англ. heavy) с мол. массой 23000–24000 и 50000–70000 соответственно. Известно также, что каждая из этих цепей имеет 2 типа доменов – вариабельные (V) участки, состоящие из 108 аминокислотных остатков, и константные (С) участки, состоящие из 110 и 350 аминокислотных остатков соответственно в L- и Н-цепях (рис. 2).
Из других гликопротеинов, выполняющих ряд важнейших биологических функций, следует отметить все белки плазмы крови (за исключением альбуминов), трансферрин, церулоплазмин, гонадотропный и фолликуло-стимулирующие гормоны, некоторые ферменты, а также гликопротеины в составе слюны (муцин), хрящевой и костной тканей и яичного белка (овомукоид). Углеводные компоненты, помимо информативной функции, значительно повышают стабильность молекул, в состав которых они входят, к различного рода химическим, физическим воздействиям и предохраняют их от действия протеиназ, определяя тем самым биологическую роль гликопротеинов. Являясь составной частью клеточной мембраны, гликопротеины участвуют, кроме того, в иммунологических реакциях, ионном обмене, процессах межклеточной адгезии и т.д.
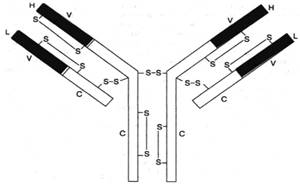
Рис. 2. Структура IgG человека. Показаны легкие (L) и тяжелые (Н) цепи, дисульфидные связи и вариабельные V (красные) и константные С (светлые) участки.
Вопрос 50. Написать реакцию НАД-зависимого дгидрирования малата в цикле КРЕБСА
Ответ. Цикл трикарбоновых кислот впервые был открыт английским биохимиком Г. Кребсом. Он первым постулировал значение данного цикла для полного сгорания пирувата, главным источником которого является гликолитическое превращение углеводов. В дальнейшем было показано, что цикл трикарбо-новых кислот является тем центром, в котором сходятся практически все метаболические пути. Таким образом, цикл Кребса – общий конечный путь окисления ацетильных групп (в виде ацетил-КоА), в которые превращается в процессе катаболизма большая часть органических молекул, играющих роль «клеточного топлива»: углеводов, жирных кислот и аминокислот.
Образовавшийся в результате окислительного декарбоксилирования пирувата в митохондриях ацетил-КоА вступает в цикл Кребса. Данный цикл происходит в матриксе митохондрий и состоит из восьми последовательных реакций (рис. 3). Начинается цикл с присоединения ацетил-КоА к оксалоацетату и образования лимонной кислоты (цитрата). Затем лимонная кислота (шестиуглеродное соединение) путем ряда дегидрирований (отнятие водорода) и двух декарбоксилирований (отщепление СО2) теряет два углеродных атома и снова в цикле Кребса превращается в оксалоацетат (четырехуглеродное соединение), т.е. в результате полного оборота цикла одна молекула ацетил-КоА сгорает до СО2 и Н2О, а молекула окса-лоацетата регенерируется. Рассмотрим все восемь последовательных реакций (этапов) цикла Кребса.
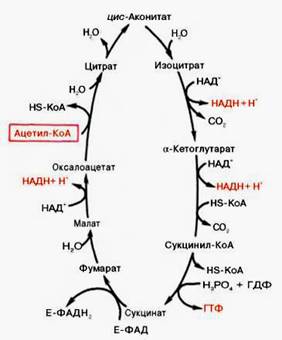
Рис. 3. Цикл трикарбоновых кислот (цикл Кребса).
Первая реакция катализируется ферментом цит-рат-синтазой, при этом ацетильная группа ацетил-КоА конденсируется с оксалоацетатом, в результате чего образуется лимонная кислота:
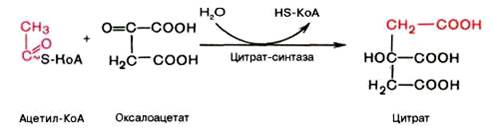
По-видимому, в данной реакции в качестве промежуточного продукта образуется связанный с ферментом цитрил-КоА. Затем последний самопроизвольно и необратимо гидролизуется с образованием цитрата и HS-KoA.
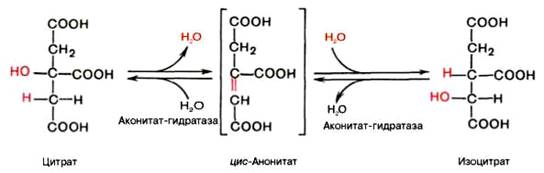
В результате второй реакции образовавшаяся лимонная кислота подвергается дегидратированию с образованием цис-аконитовой кислоты, которая, присоединяя молекулу воды, переходит в изолимонную кислоту (изоцитрат). Катализирует эти обратимые реакции гидратации–дегидратации фермент аконитатгидратаза (аконитаза). В результате происходит взаимоперемещение Н и ОН в молекуле цитрата:
Третья реакция, по-видимому, лимитирует скорость цикла Кребса. Изолимонная кислота дегидрируется в присутствии НАД-зависимой изо-цитратдегидрогеназы.
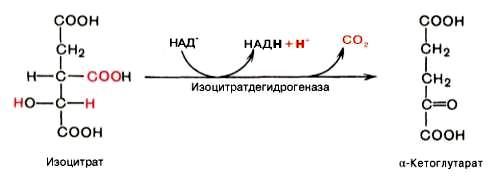
В ходе изоцитратдегидрогеназной реакции изолимонная кислота одновременно декарбоксилируется. НАД-зависимая изоцитратдегидрогеназа является аллостерическим ферментом, которому в качестве специфического активатора необходим АДФ. Кроме того, фермент для проявления своей активности нуждается в ионах Mg2+или Мn2+.
Во время четвертой реакции происходит окислительное декарбокси-лирование α-кетоглутаровой кислоты с образованием высокоэнергетического соединения сукцинил-КоА. Механизм этой реакции сходен с таковым реакции окислительного декарбоксилирования пирувата до ацетил-КоА, α-кетоглутаратдегидрогеназный комплекс напоминает по своей структуре пируватдегидрогеназный комплекс. Как в одном, так и в другом случае в реакции принимают участие 5 коферментов: ТПФ, амид липоевой кислоты, HS-KoA, ФАД и НАД+.
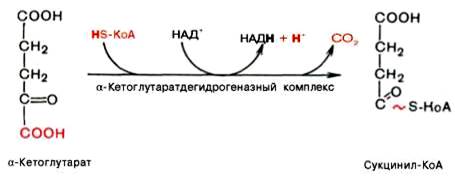
Пятая реакция катализируется ферментом сукцинил-КоА-синтета-зой. В ходе этой реакции сукцинил-КоА при участии ГТФ и неорганического фосфата превращается в янтарную кислоту (сукцинат). Одновременно происходит образование высокоэргической фосфатной связи ГТФ за счет высокоэргической тиоэфирной связи сукцинил-КоА:
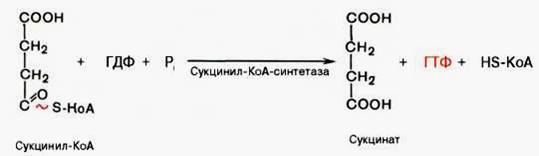
В результате шестой реакции сукцинат дегидрируется в фумаровую кислоту. Окисление сукцината катализируется сукцинатдегидрогеназой, в молекуле которой с белком прочно (ковалентно) связан кофермент ФАД. В свою очередь сукцинатдегидрогеназа прочно связана с внутренней ми-тохондриальной мембраной:
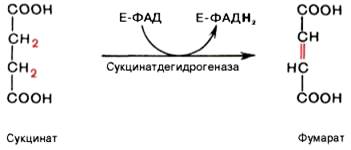
Седьмая реакция осуществляется под влиянием фермента фума-ратгидратазы (фумаразы). Образовавшаяся при этом фумаровая кислота гидратируется, продуктом реакции является яблочная кислота (малат). Следует отметить, что фумаратгидратаза обладает стереоспецифичностью – в ходе реакции образуется L-яблочная кислота:
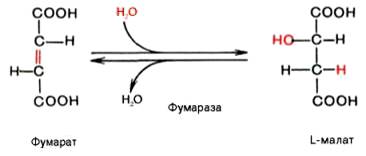
Наконец, в ходе восьмой реакции цикла трикарбоновых кислот под влиянием митохондриальной НАД-зависимой малатдегидрогеназы происходит окисление L-малата в оксалоацетат:

Как видно, за один оборот цикла, состоящего из восьми ферментативных реакций, происходит полное окисление («сгорание») одной молекулы ацетил-КоА. Для непрерывной работы цикла необходимо постоянное поступление в систему ацетил-КоА, а коферменты (НАД+ и ФАД), перешедшие в восстановленное состояние, должны снова и снова окисляться. Это окисление осуществляется в системе переносчиков электронов в дыхательной цепи (в цепи дыхательных ферментов), локализованной в мембране митохондрий. Образовавшийся ФАДН2 прочно связан с СДГ, поэтому он передает атомы водорода через KoQ. Освобождающаяся в результате окисления ацетил-КоА энергия в значительной мере сосредоточивается в макроэргических фосфатных связях АТФ. Из 4 пар атомов водорода 3 пары переносят НАДН на систему транспорта электронов; при этом в расчете на каждую пару в системе биологического окисления образуется 3 молекулы АТФ (в процессе сопряженного окислительного фосфорилирования), а всего, следовательно, 9 молекул АТФ. Одна пара атомов от сукцинатдегидрогеназы-ФАДН2 попадает в систему транспорта электронов через KoQ, в результате образуется только 2 молекулы АТФ. В ходе цикла Кребса синтезируется также одна молекула ГТФ (субстратное фосфорилирование), что равносильно одной молекуле АТФ. Итак, при окислении одной молекулы ацетил-КоА в цикле Кребса и системе окислительного фосфорилирования может образоваться 12 молекул АТФ.
Если подсчитать полный энергетический эффект гликолитического расщепления глюкозы и последующего окисления двух образовавшихся молекул пирувата до СО2 и Н2О, то он окажется значительно большим.
Как отмечалось, одна молекула НАДН (3 молекулы АТФ) образуется при окислительном декарбоксилировании пирувата в ацетил-КоА. При расщеплении одной молекулы глюкозы образуется 2 молекулы пирувата, а при окислении их до 2 молекул ацетил-КоА и последующих 2 оборотов цикла трикарбоновых кислот синтезируется 30 молекул АТФ (следовательно, окисление молекулы пирувата до СО2 и Н2О дает 15 молекул АТФ). К этому количеству надо добавить 2 молекулы АТФ, образующиеся при аэробном гликолизе, и 6 молекул АТФ, синтезирующихся за счет окисления 2 молекул внемитохондриального НАДН, которые образуются при окислении 2 молекул глицеральдегид-3-фосфата в дегидрогеназной реакции гликолиза. Следовательно, при расщеплении в тканях одной молекулы глюкозы по уравнению С6Н12О6 + 6О2 —> 6СО2 + 6Н2О синтезируется 38 молекул АТФ. Несомненно, что в энергетическом отношении полное расщепление глюкозы является более эффективным процессом, чем анаэробный гликолиз.
Необходимо отметить, что образовавшиеся в процессе превращения глицеральдегид-3-фосфата 2 молекулы НАДН в дальнейшем при окислении могут давать не 6 молекул АТФ, а только 4. Дело в том, что сами молекулы внемитохондриального НАДН не способны проникать через мембрану внутрь митохондрий. Однако отдаваемые ими электроны могут включаться в митохондриальную цепь биологического окисления с помощью так называемого глицеролфосфатного челночного механизма (рис. 4).

Рис. 4. Глицеролфосфатный челночный механизм. Объяснение в тексте.
Ци-топлазматический НАДН сначала реагирует с цитоплазматическим ди-гидроксиацетонфосфатом, образуя глицерол-3-фосфат. Реакция катализи-руется НАД-зависимой цитоплазматической глицерол-3-фосфат-дегидроге-назой:
Дигидроксиацетонфосфат + НАДН + Н+ <=> Глицерол-3-фосфат + НАД+.
Образовавшийся глицерол-3-фосфат легко проникает через митохонд-риальную мембрану. Внутри митохондрии другая (митохондриальная) глицерол-3-фосфат-дегидрогеназа (флавиновый фермент) снова окисляет глицерол-3-фосфат до диоксиацетонфосфата:
Глицерол-3-фосфат + ФАД <=> Диоксиацетонфосфат + ФАДН2.
Восстановленный флавопротеин (фермент-ФАДН2) вводит на уровне KoQ приобретенные им электроны в цепь биологического окисления и сопряженного с ним окислительного фосфорилирования, а диоксиаце-тонфосфат выходит из митохондрий в цитоплазму и может вновь взаимодействовать с цитоплазматическим НАДН + Н+. Таким образом, пара электронов (из одной молекулы цитоплазматического НАДН + Н+), вводимая в дыхательную цепь с помощью глицеролфосфатного челночного механизма, дает не 3, а 2 АТФ.
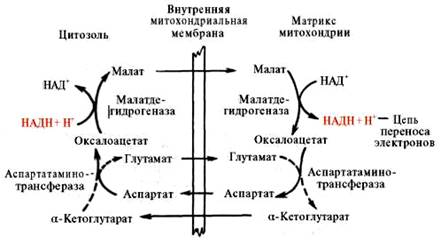
Рис. 5. Малат-аспартатная челночная система для переноса восстанавливающих эквивалентов от цитозольного НАДН в митохондриальный матрикс. Объяснение в тексте.
В дальнейшем было показано, что с помощью данного челночного механизма лишь в скелетных мышцах и мозге осуществляется перенос восстановленных эквивалентов от цитозольного НАДН + Н+ в митохондрии.
В клетках печени, почек и сердца действует более сложная малат-ас-партатная челночная система. Действие такого челночного механизма становится возможным благодаря присутствию малатдегидрогеназы и ас-партатаминотрансферазы как в цитозоле, так и в митохондриях.
Установлено, что от цитозольного НАДН + Н+ восстановленные эквиваленты сначала при участии фермента малатдегидрогеназы (рис. 5) переносятся на цитозольный оксалоацетат. В результате образуется малат, который с помощью системы, транспортирующей дикарбоновые кислоты, проходит через внутреннюю мембрану митохондрии в матрикс. Здесь малат окисляется в оксалоацетат, а матриксный НАД+ восстанавливается в НАДН + Н+, который может теперь передавать свои электроны в цепь дыхательных ферментов, локализованную на внутренней мембране митохондрии. В свою очередь образовавшийся оксалоацетат в присутствии глутамата и фермента АсАТ вступает в реакцию трансаминирования. Образующиеся аспарат и α-кетоглутарат с помощью специальных транспортных систем способны проходить через мембрану митохондрий.
Транспортирование в цитозоле регенерирует оксалоацетат, что вызывает к действию следующий цикл. В целом процесс включает легкообратимые реакции, происходит без потребления энергии, «движущей силой» его является постоянное восстановление НАД+ в цитозоле гли-церальдегид-3-фосфатом, образующимся при катаболизме глюкозы.
Итак, если функционирует малат-аспартатный механизм, то в результате полного окисления одной молекулы глюкозы может образоваться не 36, а 38 молекул АТФ (табл. 10.1).
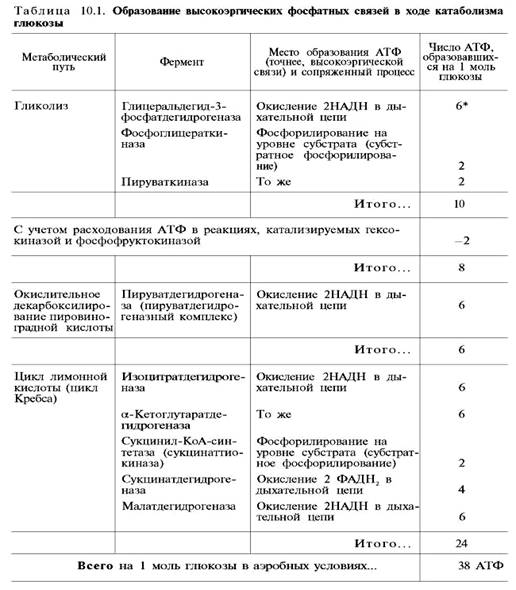
В табл. 10.1 приведены реакции, в которых происходит образование высокоэргических фосфатных связей в ходе катаболизма глюкозы, с указанием эффективности процесса в аэробных и анаэробных условиях.
Вопрос 52. Что такое окислительное фосфорелирование? Указать участки сопряжения окисления и фосфорелирования в цепях переноса электронов
Ответ. Окислительное фосфорелирование - это синтез АТФ из аденозиндифосфата и неорг. фосфата, осуществляющийся в живых клетках, благодаря энергии, выделяющейся при окислении орг. в-в в процессе клеточного дыхания. В общем виде О.ф. и его место в обмене в-в можно представить схемой:
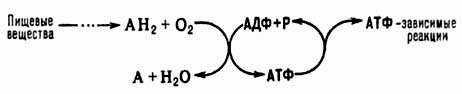
АН2-орг. в-ва, окисляемые в дыхат. цепи (т. наз. субстраты окисления, или дыхания), АДФ-аденозиндифосфат, Р-неорг. фосфат.
Поскольку АТФ необходим для осуществления мн. процессов, требующих затраты энергии (биосинтез, совершение мех. работы, транспорт в-в и др.), О.ф. играет важнейшую роль в жизнедеятельности аэробных организмов. Образование АТФ в клетке происходит также благодаря др. процессам, напр. в ходе гликолиза и разл. типов брожения, протекающих без участия кислорода. Их вклад в синтез АТФ в условиях аэробного дыхания составляет незначит. часть от вклада О.ф. (ок. 5%).
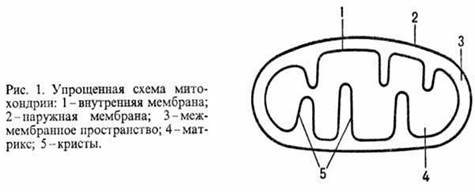
У животных, растений и грибов О.ф. протекает в спе-циализир. субклеточных структурах-митохондриях (рис. 1); у бактерий ферментные системы, осуществляющие этот процесс, находятся в клеточной мембране.
Митохондрии окружены белково-фосфолипидной мембраной. Внутри митохондрий (в т. наз. матриксе) идет ряд метаболич. процессов распада пищ. в-в, поставляющих субстраты окисления АН2 для О.ф. Наиб. важные из этих процессов -трикарбоновых кислот цикл и т. наз.![]() -окисление жирных к-т (окислит. расщепление жирной к-ты с образованием ацетил-кофермента А и к-ты, содержащей на 2 атома С меньше, чем исходная; вновь образующаяся жирная к-та также может подвергаться
-окисление жирных к-т (окислит. расщепление жирной к-ты с образованием ацетил-кофермента А и к-ты, содержащей на 2 атома С меньше, чем исходная; вновь образующаяся жирная к-та также может подвергаться ![]() -окислению). Интермедиаты этих процессов подвергаются дегидрированию (окислению) при участии ферментов дегидрогеназ; затем электроны передаются в дыхат. цепь митохондрий-ансамбль окислит.-восстановит. ферментов, встроенных во внутр. митохонд-риальную мембрану. Дыхат. цепь осуществляет многоступенчатый экзэргонич. перенос электронов (сопровождается уменьшением своб. энергии) от субстратов к кислороду, а высвобождающаяся энергия используется расположенным в той же мембране ферментом АТФ-синтетазой, для фос-форилирования АДФ до АТФ. В интактной (неповрежденной) митохондриальной мембране перенос электронов в дыхат. цепи и фосфорилирование тесно сопряжены между собой. Так, напр., выключение фосфорилирования по исчерпании АДФ либо неорг. фосфата сопровождается торможением дыхания (эффект дыхат. контроля). Большое число повреждающих митохондриальную мембрану воздействий нарушает сопряжение между окислением и фосфори-лированием, разрешая идти переносу электронов и в отсутствие синтеза АТФ (эффект разобщения).
-окислению). Интермедиаты этих процессов подвергаются дегидрированию (окислению) при участии ферментов дегидрогеназ; затем электроны передаются в дыхат. цепь митохондрий-ансамбль окислит.-восстановит. ферментов, встроенных во внутр. митохонд-риальную мембрану. Дыхат. цепь осуществляет многоступенчатый экзэргонич. перенос электронов (сопровождается уменьшением своб. энергии) от субстратов к кислороду, а высвобождающаяся энергия используется расположенным в той же мембране ферментом АТФ-синтетазой, для фос-форилирования АДФ до АТФ. В интактной (неповрежденной) митохондриальной мембране перенос электронов в дыхат. цепи и фосфорилирование тесно сопряжены между собой. Так, напр., выключение фосфорилирования по исчерпании АДФ либо неорг. фосфата сопровождается торможением дыхания (эффект дыхат. контроля). Большое число повреждающих митохондриальную мембрану воздействий нарушает сопряжение между окислением и фосфори-лированием, разрешая идти переносу электронов и в отсутствие синтеза АТФ (эффект разобщения).
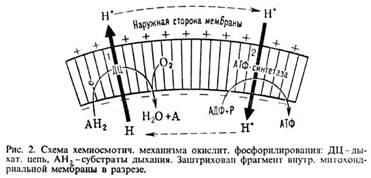
Механизм О.ф. можно представить схемой: Перенос электронов (дыхание) ![]() А ~ В
А ~ В![]() АТФ А ~ В-высокоэнергетич. интермедиат. Предполагалось, что А ~ В - хим. соед. с макроэргич. связью, напр. фосфорилир. фермент дыхат. цепи (хим. гипотеза сопряжения), или напряженная конформация к.-л. белка, участвующего в О.ф. (конформац. гипотеза сопряжения). Однако эти гипотезы не получили эксперим. подтверждения. Наиб. признанием пользуется хемиосмотич. концепция сопряжения, предложенная в 1961 П. Митчеллом (за развитие этой концепции в 1979 ему присуждена Нобелевская премия). Согласно этой теории, своб. энергия транспорта электронов в дыхат. цепи затрачивается на перенос из митохондрий через митохондриальную мембрану на ее наружную сторону ионов Н+ (рис. 2, процесс 1). В результате на мембране возникает разность электрич. потенциалов
АТФ А ~ В-высокоэнергетич. интермедиат. Предполагалось, что А ~ В - хим. соед. с макроэргич. связью, напр. фосфорилир. фермент дыхат. цепи (хим. гипотеза сопряжения), или напряженная конформация к.-л. белка, участвующего в О.ф. (конформац. гипотеза сопряжения). Однако эти гипотезы не получили эксперим. подтверждения. Наиб. признанием пользуется хемиосмотич. концепция сопряжения, предложенная в 1961 П. Митчеллом (за развитие этой концепции в 1979 ему присуждена Нобелевская премия). Согласно этой теории, своб. энергия транспорта электронов в дыхат. цепи затрачивается на перенос из митохондрий через митохондриальную мембрану на ее наружную сторону ионов Н+ (рис. 2, процесс 1). В результате на мембране возникает разность электрич. потенциалов ![]() и разность хим. активностей ионов Н+
и разность хим. активностей ионов Н+ ![]() (внутри митохондрий рН выше, чем снаружи). В сумме эти компоненты дают трансмембранную разность электрохим. потенциалов ионов водорода
(внутри митохондрий рН выше, чем снаружи). В сумме эти компоненты дают трансмембранную разность электрохим. потенциалов ионов водорода ![]() между матриксом митохондрий и внеш. водной фазой, разделенными мембраной:
между матриксом митохондрий и внеш. водной фазой, разделенными мембраной:
![]()
где R-универсальная газовая постоянная, T-абс. т-ра, F- число Фарадея. Величина ![]() обычно составляет ок. 0,25 В, причем осн. часть (0,15-0,20 В) представлена электрич. составляющей
обычно составляет ок. 0,25 В, причем осн. часть (0,15-0,20 В) представлена электрич. составляющей ![]() . Энергия
. Энергия ![]() , выделяющаяся при движении протонов внутрь митохондрий по электрич. полю в сторону меньшей их концентрации (рис. 2, процесс 2), используется АТФ-синтетазой для синтеза АТФ. Т. обр., схему О.ф., согласно этой концепции, можно представить в след. виде:
, выделяющаяся при движении протонов внутрь митохондрий по электрич. полю в сторону меньшей их концентрации (рис. 2, процесс 2), используется АТФ-синтетазой для синтеза АТФ. Т. обр., схему О.ф., согласно этой концепции, можно представить в след. виде:
Перенос электронов (дыхание) ![]() АТФ
АТФ
Сопряжение окисления и фосфорилирования через![]() позволяет объяснить, почему О.ф., в отличие от гликолитич. ("субстратного") фосфорилирования, протекающего в р-ре, возможно лишь в замкнутых мембранных структурах, а также почему все воздействия, снижающие электрич. сопротивление и увеличивающие протонную проводимость мембраны, подавляют ("разобщают") О.ф. Энергия
позволяет объяснить, почему О.ф., в отличие от гликолитич. ("субстратного") фосфорилирования, протекающего в р-ре, возможно лишь в замкнутых мембранных структурах, а также почему все воздействия, снижающие электрич. сопротивление и увеличивающие протонную проводимость мембраны, подавляют ("разобщают") О.ф. Энергия ![]() , помимо синтеза АТФ, может непосредственно использоваться клеткой для др. целей - транспорта метаболитов, движения (у бактерий), восстановления нико-тинамидных коферментов и др.
, помимо синтеза АТФ, может непосредственно использоваться клеткой для др. целей - транспорта метаболитов, движения (у бактерий), восстановления нико-тинамидных коферментов и др.
В дыхат. цепи имеется неск. участков, к-рые характеризуются значит. перепадом окислит.-восстановит. потенциала ![]() и сопряжены с запасанием энергии (генерацией
и сопряжены с запасанием энергии (генерацией ![]() ). Таких участков, наз. пунктами или точками сопряжения, обычно три: НАДН: убихинон-редуктазное звено (
). Таких участков, наз. пунктами или точками сопряжения, обычно три: НАДН: убихинон-редуктазное звено (![]() 0,35-0,4 В), убихинол: цитохром-c-редуктазное звено (
0,35-0,4 В), убихинол: цитохром-c-редуктазное звено (![]() ~ ~ 0,25 В) и цитохром-с-оксидазный комплекс (
~ ~ 0,25 В) и цитохром-с-оксидазный комплекс (![]() ~ 0,6 В)-пункты сопряжения 1, 2 и 3 соотв. (рис. 3). Каждый из пунктов сопряжения дыхат. цепи м.б. выделен из мембраны в виде индивидуального ферментного комплекса, обладающего окислит.-восстановит. активностью. Такой комплекс, встроенный в фосфолипидную мембрану, способен функционировать как протонный насос.
~ 0,6 В)-пункты сопряжения 1, 2 и 3 соотв. (рис. 3). Каждый из пунктов сопряжения дыхат. цепи м.б. выделен из мембраны в виде индивидуального ферментного комплекса, обладающего окислит.-восстановит. активностью. Такой комплекс, встроенный в фосфолипидную мембрану, способен функционировать как протонный насос.
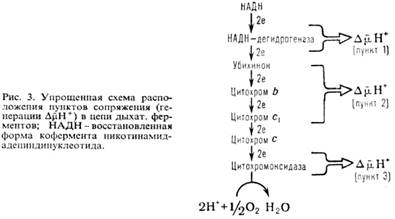
Обычно для характеристики эффективности О.ф. используют величины Н+/2е или q/2e, указывающие сколько протонов (либо электрич. зарядов) переносится через мембрану при транспорте пары электронов через данный участок дыхат. цепи, а также отношение Н+/АТФ, показывающее, сколько протонов нужно перенести снаружи внутрь митохондрий через АТФ-синтетазу для синтеза 1 молекулы АТФ. Величина q/2e составляет для пунктов сопряжения 1, 2 и 3 соотв. 3-4, 2 и 4. Величина Н+/АТФ при синтезе АТФ внутри митохондрий равна 2; однако еще один Н+ может тратиться на вынос синтезированного АТФ4- из матрикса в цитоплазму переносчиком адениновых нуклеотидов в обмен на АДФ -3 . Поэтому кажущаяся величина Н+ / АТФнаружн равна 3.
В организме О.ф. подавляется мн. токсичными в-вами, к-рые по месту их действия можно разделить на три группы: 1) ингибиторы дыхат. цепи, или т. наз. дыхат. яды. 2) Ингибиторы АТФ-синтетазы. Наиб. распространенные ингибиторы этого класса, употребляемые в лаб. исследованиях, - антибиотик олигомицин и модификатор карбоксильных групп белка дициклогексилкарбодиимид. 3) Т. наз. разобщители О.ф. Они не подавляют ни перенос электронов, ни собственно фосфорилирование АДФ, но обладают способностью уменьшать величину ![]() на мембране, благодаря чему нарушается энергетич. сопряжение между дыханием и синтезом АТФ. Разобщающее действие проявляет большое число соед. самой разнообразной хим. структуры. Классич. разобщители - в-ва, обладающие слабыми кислотными св-вами, способные проникать через мембрану как в ионизованной (депротонированной), так и в нейтральной (протонированной) формах. К таким в-вам относят, напр., 1-(2-дицианометилен)гидразино-4-трифтор-метоксибензол, или карбонилцианид-n-трифторметокси-фенилгидразон, и 2,4-динитрофенол (соотв. ф-лы I и II; показаны протонир. и депротонир. формы).
на мембране, благодаря чему нарушается энергетич. сопряжение между дыханием и синтезом АТФ. Разобщающее действие проявляет большое число соед. самой разнообразной хим. структуры. Классич. разобщители - в-ва, обладающие слабыми кислотными св-вами, способные проникать через мембрану как в ионизованной (депротонированной), так и в нейтральной (протонированной) формах. К таким в-вам относят, напр., 1-(2-дицианометилен)гидразино-4-трифтор-метоксибензол, или карбонилцианид-n-трифторметокси-фенилгидразон, и 2,4-динитрофенол (соотв. ф-лы I и II; показаны протонир. и депротонир. формы).
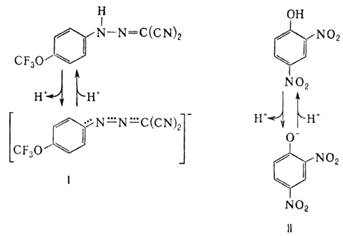
Двигаясь через мембрану в электрич. поле в ионизованной форме, разобщитель уменьшает![]() ; возвращаясь обратно в протонир. состоянии, разобщитель понижает
; возвращаясь обратно в протонир. состоянии, разобщитель понижает ![]() (рис. 4). Т. обр., такой "челночный" тип действия разобщителя приводит к уменьшению
(рис. 4). Т. обр., такой "челночный" тип действия разобщителя приводит к уменьшению![]()
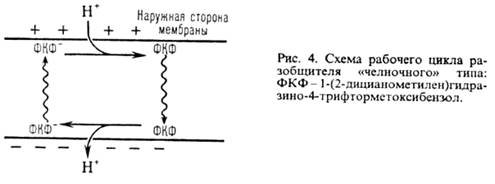
Разобщающим действием обладают также ионофоры (напр., грамицидин), повышающие электропроводность мембраны в результате образования ионных каналов или в-ва, разрушающие мембрану (напр., детергенты).
О.ф. открыто В. А. Энгельгардтом в 1930 при работе с эритроцитами птиц. В 1939 В. А. Белицер и Е. Т. Цыбакова показали, что О.ф. сопряжено с переносом электронов в процессе дыхания; к такому же заключению несколько позднее пришел Г. М. Калькар.
Вопрос 64. Напишите фрагменты молекул клетчатки, крахмала. Укажите какой углевод не переваривается в ЖКТ человека и почему?
Ответ. Крахмал и клетчатка являются природными полимерами глюкозы. Крахмал - полимера-глюкозы, клетчатка - полимер --> глюкозы. Крахмал имеет как линейные цепи (амилоза), в которых связь между а-глюкозами -а, а—1, 4-0-гликозидная, так и разветвленные цепи (амилопектин) с дополнительными а,ос-1,6-а-гликозидными связями в местах ветвления, n = 1000 до 6000.
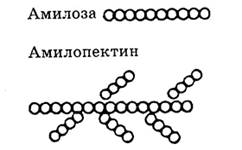
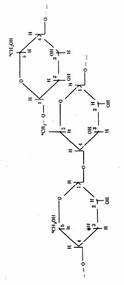
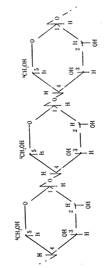
Фрагмент молекулы крахмала. Фрагмент молекулы клетчатки.
М. крахмала до 1 млн. Д.
Клетчатка имеет только линейное строение. Этим объясняется, что целлюлоза (клетчатка) образует такие волокнистые материалы, как хлопок, лен. Между параллельными макромолекулами образуются водородные связи, которые обусловливают высокую прочность этих волокон.
М. клетчатки до 20 млн. Д; n до 40.000.
В клетчатке 1,4-B,B-гликозидные связи.
Углеводы служат важнейшим источником энергии, обеспечивая детям старше года до 50-60% общей калорийности рациона. Способность углеводов служить высокоэффективным источником энергии лежит в основе их «сберегающего белок» действия.
При поступлении с пищей достаточного количества углеводов аминокислоты лишь в незначительной степени используются в организме как энергетический материал и используются в основном для различных пластических (строительных) нужд организма.
При этом углеводы играют ведущую роль в процессах клеточной рецепции гормонов и других биологически активных соединений и межклеточном взаимодействии, имеющем существенное значение для нормального хода процессов клеточного роста, дифференцировки и иммунитета.
Хотя углеводы не принадлежат к числу незаменимых факторов питания и могут образовываться в организме из аминокислот и глицерина, их полное исключение из рациона ведет к резким нарушениям метаболических процессов (процессам, в ходе которых организм расщепляет содержащиеся в продуктах питательные вещества, чтобы произвести энергию, необходимую для жизнедеятельности).
Самые важные из углеводов - сахароза, глюкоза, фруктоза, а также крахмал и гликоген, клетчатка. Они быстро усваиваются ("сгорают") в организме. Исключение составляет клетчатка (целлюлоза или растительные волокна), которой особенно много в растительной пище. Она практически не усваивается организмом, но имеет большое значение: выступает в роли балласта и помогает пищеварению, механически очищая слизистые оболочки желудка и кишечника.
Вопрос 75. Написать в виде схемы реакции синтеза гликогена
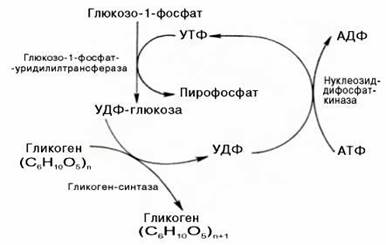
Ответ. Прежде всего, глюкоза подвергается фосфорилированию при участии фермента гексокиназы, а в печени – и глюкокиназы. Далее глюкозо-6-фосфат под влиянием фермента фосфоглюкомутазы переходит в глюкозо-1-фос-фат:

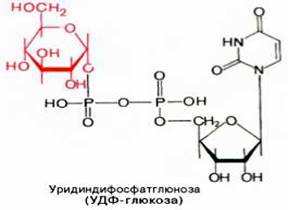
Образовавшийся глюкозо-1-фосфат уже непосредственно вовлекается в синтез гликогена. На первой стадии синтеза глюкозо-1-фосфат вступает во взаимодействие с УТФ (уридинтрифосфат), образуя уридиндифосфатглю-козу (УДФ-глюкоза) и пирофосфат. Данная реакция катализируется ферментом глюкозо-1-фосфат-уридилилтрансферазой (УДФГ-пирофосфорила-за):
Глюкозо-1-фосфат + УТФ < = > УДФ-глюкоза + Пирофосфат.
Приводим структурную формулу УДФ-глюкозы:
На второй стадии – стадии образования гликогена – происходит перенос глюкозного остатка, входящего в состав УДФ-глюкозы, на глюкозидную цепь гликогена («затравочное» количество). При этом образуется α-(1–>4)-связь между первым атомом углерода добавляемого остатка глюкозы и 4-гидроксильной группой остатка глюкозы цепи. Эта реакция катализируется ферментом гликогенсинтазой. Необходимо еще раз подчеркнуть, что реакция, катализируемая гликогенсинтазой, возможна только при условии, что полисахаридная цепь уже содержит более 4 остатков D-глюкозы.
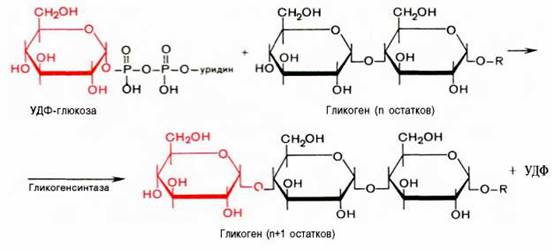
Образующийся УДФ затем вновь фосфорилируется в УТФ за счет АТФ, и таким образом весь цикл превращений глюкозо-1-фосфата начинается сначала.
В целом образование α-1,4-глюкозидной ветви («амилозной» ветви) гликогена можно представить в виде следующей схемы:
Установлено, что гликогенсинтаза неспособна катализировать образование α-(1–>6)-связи, имеющейся в точках ветвления гликогена. Этот процесс катализирует специальный фермент, получивший название гли-когенветвящего фермента, или амило-(1–>4)–>(1–>6)-трансглюкозидазы. Последний катализирует перенос концевого олигосахаридного фрагмента, состоящего из 6 или 7 остатков глюкозы, с нередуцирующего конца одной из боковых цепей, насчитывающей не менее 11 остатков, на 6-гидроксиль-ную группу остатка глюкозы той же или другой цепи гликогена. В результате образуется новая боковая цепь.
Ветвление повышает растворимость гликогена. Кроме того, благодаря ветвлению создается большое количество невосстанавливающих концевых остатков, которые являются местами действия гликогенфосфорилазы и гликогенсинтазы.
Таким образом, ветвление увеличивает скорость синтеза и расщепления гликогена.
Благодаря способности к отложению гликогена (главным образом в печени и мышцах и в меньшей степени в других органах и тканях) создаются условия для накопления в норме некоторого резерва углеводов. При повышении энерготрат в организме в результате возбуждения ЦНС обычно происходят усиление распада гликогена и образование глюкозы.
Помимо непосредственной передачи нервных импульсов к эффекторным органам и тканям, при возбуждении ЦНС повышаются функции ряда желез внутренней секреции (мозговое вещество надпочечников, щитовидная железа, гипофиз и др.), гормоны которых активируют распад гликогена, прежде всего в печени и мышцах.
Как отмечалось, эффект катехоламинов в значительной мере опосредован действием цАМФ, который активирует протеинкиназы тканей. При участии последних происходит фосфорилирование ряда белков, в том числе гликогенсинтазы и фосфорилазы b – ферментов, участвующих в обмене углеводов. Фосфорилированный фермент гликогенсинтаза сам по себе малоактивен или полностью неактивен, но в значительной мере активируется положительным модулятором глюкозо-6-фосфатом, который увеличивает Vmaxфермента. Эта форма гликогенсинтазы называется D-формой, или зависимой (dependent) формой, поскольку ее активность зависит от глюкозо-6-фосфата. Дефосфорилированная форма гликоген-синтазы, называемая также I-формой, или независимой (independent) формой, активна и в отсутствие глюкозо-6-фосфата.
Таким образом, адреналин оказывает двойное действие на обмен углеводов: ингибирует синтез гликогена из УДФ-глюкозы, поскольку для проявления максимальной активности D-формы гликогенсинтазы нужны очень высокие концентрации глюкозо-6-фосфата, и ускоряет распад гликогена, так как способствует образованию активной фосфорилазы а. В целом суммарный результат действия адреналина состоит в ускорении превращения гликогена в глюкозу.
Вопрос 84. Какие вещества необходимы для синтеза жирных кислот на синтаве жирных кислот. Как связан процесс синтеза жирных кислот с обменом углеводов?
Ответ. Из приведенной общей схемы (рис. 4) видно также, что имеются различные пути взаимопревращений жиров и углеводов. Практика откорма сельскохозяйственных животных давно подтвердила возможность синтеза жиров из углеводов пищи. С энергетической точки зрения, превращение углеводов в жиры следует рассматривать как накопление и депонирование энергии, хотя синтез жира сопровождается затратой энергии, которая вновь освобождается при окислении жиров в организме.
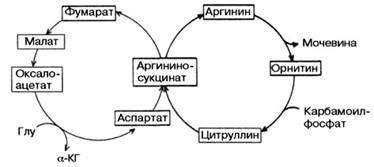
Рис. 4. The "Krebs Bicycle".
Глицерин, входящий в состав триацилглицеролов и фосфоглицеринов, может легко образоваться из промежуточных метаболитов гликолиза, в частности из глицераль-дегид-3-фосфата. Следует, однако, подчеркнуть, что основным путем превращения углеводов в жиры является путь образования высших жирных кислот из ацетил-КоА, который образуется при окислительном декар-боксилировании пирувата. Последняя реакция практически необратима, поэтому образования углеводов из высших жирных кислот почти не происходит. Таким образом, синтез углеводов из жиров в принципе может происходить только из глицерина, хотя в обычных условиях реакция протекает в обратную сторону, т.е. в сторону синтеза жиров из глицерина, образующегося при окислении углеводов. Ацетил-КоА, образующийся в процессе обмена углеводов, жиров и ряда аминокислот, служит пусковым субстратом как для синтеза жирных кислот (а следовательно, и липидов вообще), так и для цикла трикарбоновых кислот. Для окисления ацетил-КоА в этом цикле требуется оксалоацетат, который является вторым ключевым субстратом в цикле Кребса. Оксалоацетат может синтезироваться из пировиноградной кислоты и СО2 благодаря реакции карбокси-лирования или образоваться из аспарагиновой кислоты в процессе транс-аминирования с α-кетоглутаратом. Две молекулы ацетил-КоА, конденсируясь, образуют ацетоуксусную кислоту (ацетоацетат), которая является источником других кетоновых тел в организме, в частности β-оксимасляной кислоты (β-оксибутирата) и ацетона. Следует подчеркнуть, что ацетоуксусная и β-оксимасляная кислоты часто рассматриваются как транспортные формы активной уксусной кислоты, доставляющие ее для окисления в цикле Кребса в периферических тканях. Эти же реакции конденсации двух молекул ацетил-КоА составляют начальные этапы синтеза холестерина, в свою очередь являющегося предшественником гормонов стероидной природы, витамина D3, а также желчных кислот. Последние в виде парных желчных кислот выполняют важную функцию эмульгаторов при переваривании липидов пищи в кишечнике, а также функцию транспортеров, способствуя всасыванию высших жирных кислот. Следует указать также на использование галактозы и частично глюкозы для биосинтеза цереброзидов и гликолипидов, выполняющих важные и специфические функции в деятельности ЦНС. В этом синтезе участвуют не свободные моносахариды, а гексозамины (галактозамин и глюкозамин), биосинтез которых в свою очередь требует доставки амидного азота глутамина, интегрируя тем самым обмен углеводов, липидов и белков.
Вопрос 88. Какими липопротеинами крови доставляются жиры из печени и кишечника к жировым депо? Каков состав этих липопротеинов?
Ответ. У человека основная масса β-липопротеинов (липопротеины низкой плотности – ЛПНП) образуется в плазме крови из ЛПОНП при действии липопротеинлипазы. В ходе этого процесса образуются сначала промежуточные короткоживущие липопротеины (Пр.ЛП), а затем формируются частицы, обедненные триглицеридами и обогащенные холестерином, т.е. ЛПНП.
При высоком содержании жирных кислот в плазме их поглощение печенью возрастает, усиливается синтез триглицеридов, а также окисление жирных кислот, что может привести к повышенному образованию кетоновых тел.
Следует подчеркнуть, что кетоновые тела образуются в печени в ходе так называемого β-гидрокси-β-метилглутарил-КоА пути. Однако существует мнение, что ацетоацетил-КоА, являющийся исходным соединением при кетогенезе, может образоваться как непосредственно в ходе β-окисле-ния жирных кислот, так и в результате конденсации ацетил-КоА [Марри Р. и др., 1993]. Из печени кетоновые тела током крови доставляются в ткани и органы (мышцы, почки, мозг и др.), где они быстро окисляются при участии соответствующих ферментов, т.е. по сравнению с другими тканями печень является исключением.
В печени происходит интенсивный распад фосфолипидов, а также их синтез. Помимо глицерина и жирных кислот, которые входят в состав нейтральных жиров, для синтеза фосфолипидов необходимы неорганические фосфаты и азотистые соединения, в частности холин, для синтеза фосфатидхолина. Неорганические фосфаты в печени имеются в достаточном количестве. При недостаточном образовании или недостаточном поступлении в печень холина синтез фосфолипидов из компонентов нейтрального жира становится либо невозможным, либо резко снижается и нейтральный жир откладывается в печени. В этом случае говорят о жировой инфильтрации печени, которая может затем перейти в ее жировую дистрофию. Иными словами, синтез фосфолипидов лимитируется количеством азотистых оснований, т.е. для синтеза фосфоглицеридов необходим либо холин, либо соединения, которые могут являться донорами метильных групп и участвовать в образовании холина (например, метионин). Такие соединения получили название липотропных веществ. Отсюда становится ясным, почему при жировой инфильтрации печени весьма полезен творог, содержащий белок казеин, в составе которого имеется большое количество остатков аминокислоты метионина.
В последние годы достигнут определенный прогресс в выяснении химической природы и структуры липопротеинов (ЛП). Этот класс сложных белков состоит из белка и простетической группы, представленной каким-либо липидом. В частности, в составе липопротеинов открыты нейтральные жиры, свободные жирные кислоты, фосфолипиды, холестериды. Липопротеины широко распространены в природе: в растениях, тканях животных и у микроорганизмов – и выполняют разнообразные биологические функции. Они входят в состав клеточной мембраны и внутриклеточных биомембран ядра, митохондрий, микросом (структурированные липопро-теины), а также присутствуют в свободном состоянии (главным образом в плазме крови). К липопротеинам относятся, кроме того, тромбопластический белок ткани легких, липовителлин желтка куриного яйца, некоторые фосфолипиды молока и т.д. Установлено, что липопротеины участвуют в структурной, комплексной организации миелиновых оболочек, нервной ткани, хлоропластов, фоторецепторной и электронно-транспортной систем, палочек и колбочек сетчатки и др.
Большинство ЛП синтезируется в печени или в слизистой оболочке кишечника. Они содержат гидрофобное липидное ядро, окруженное полярными липидами и оболочкой из белков, получивших название апобелки. Различают 8 типов апобелков: апо-AI, АII, В, CI, СII, CIII, D и Е. Обычно ЛП содержат до 5% углеводов (глюкоза, галактоза, гексозамины, фукоза, сиаловая кислота), поэтому некоторые из них являются и гликопротеинами.
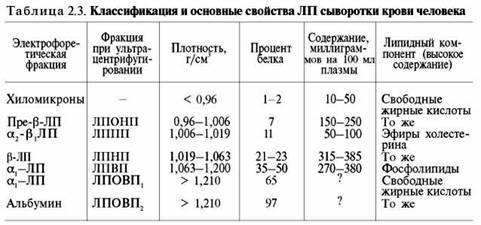
Липопротеины сыворотки крови подразделяют на отдельные классы в зависимости от электрофоретической подвижности (с белками крови) и от плотности при ультрацентрифугировании. Различают ЛП низкой плотности (ЛПНП), очень низкой плотности (ЛПОНП), высокой плотности (ЛПВП), очень высокой плотности (ЛПОВП) и ЛП промежуточной плотности (ЛППП) (табл. 2.3).
Механизм связывания белкового компонента с липидами. Имеются данные, что в образовании липопротеинов участвуют нековалентные силы различной природы, определяемые наличием или отсутствием у липидного компонента ионизированных групп атомов. Если в образовании липопро-теина участвуют фосфолипиды, то между ними и белковой молекулой возникает ионный тип связи (рис. 2.3).
Доказано также существование гидрофобных взаимодействий между неполярными группами липидного компонента (например, радикалы жирных кислот) и белковой молекулы. Чаще в липопротеинах действуют комбинированно разные нековалентные силы, способствуя образованию в высшей степени упорядоченной двойной белково-липидной структуры биомембран.
Вопрос 96. В состав пищи входили пептиды следующего состава: ала-сер-гли-дис-сен-лиз-три-вал-лей. Назовите ферменты и соответствующие связи, которые они расщепляют: а) в желудке, б) в просвете тонкой кишки, в) в слизистой оболочке тонкой кишки
Ответ. Строительный материал для мышц и энергию, необходимую для жизнедеятельности, организм получает исключительно из пищи. Получение энергии из пищи - вершина эволюционного механизма потребления энергии. В процессе переваривания пища превращается в составные элементы, которые могут быть использованы организмом.
При высоких физических нагрузках потребность в пищевых веществах может быть настолько велика, что даже здоровый желудочно-кишечный тракт не способен будет обеспечить организм достаточным количеством пластического и энергетического материала. В связи с этим, возникает противоречие между потребностью организма в пищевых веществах и способностью желудочно-кишечного тракта эту потребность удовлетворить.
Попробуем рассмотреть способы решения этой проблемы.
Для того, чтобы понять, каким образом лучше всего повысить переваривающую способность желудочно-кишечного тракта, необходимо сделать краткий экскурс в физиологию.
В химических преобразованиях пищи самую важную роль играет секреция пищеварительных желез. Она строго координирована. Пища, передвигаясь по желудочно-кишечному тракту, подвергается поочередному воздействию различных пищеварительных желез.
Понятие “пищеварение” неразрывно связано с понятием пищеварительных ферментов. Пищеварительные ферменты – это узкоспециализированная часть ферментов, основная задача которых - расщепление сложных пищевых веществ в желудочно-кишечном тракте до более простых, которые уже непосредственно усваиваются организмом.
Рассмотрим основные компоненты пищи:
Углеводы. Простые углеводы сахара (глюкоза, фруктоза) переваривания не требуют. Они благополучно всасываются в ротовой полости, 12-и перстной кишке и тонком кишечнике.
Сложные углеводы - крахмал и гликоген требуют переваривания (расщепления) до простых сахаров.
Частичное расщепление сложных углеводов начинается уже в ротовой полости, т.к. слюна содержит амилазу - фермент, расщепляющий углеводы. Амилаза слюны L-амилаза, осуществляет лишь первые фазы распада крахмала или гликогена с образованием декстринов и мальтозы. В желудке действие слюнной L -амилазы прекращается из-за кислой реакции содержимого желудка (рН 1,5-2,5). Однако в более глубоких слоях пищевого комка, куда не сразу проникает желудочный сок, действие слюнной амилазы некоторое время продолжается и происходит расщепление полисахаридов с образованием декстринов и мальтозы.
Когда пища попадает в 12-и перстную кишку, там осуществляется самая важная фаза превращения крахмала (гликогена), рН возрастает до нейтральной среды и L -амилаза максимально активизируется. Крахмал и гликоген полностью распадаются до мальтозы. В кишечнике мальтоза очень быстро распадается на 2 молекулы глюкозы, которые быстро всасываются.
Дисахариды.
Сахароза (простой сахар), попавшая в тонкий кишечник, под действием фермента сахарозы быстро превращается в глюкозу и фруктозу.
Лактоза, молочный сахар, который содержится только в молоке, под действием фермента лактозы.
В конце концов, все углеводы пищи распадаются на составляющие их моносахариды (преимущественно глюкоза, фруктоза и галактоза), которые всасываются кишечной стенкой и затем попадают в кровь. Свыше 90% всосавшихся моносахаридов (главным образом глюкозы) через капилляры кишечных ворсинок попадают в кровеносную систему и с током крови доставляются прежде всего в печень. В печени большая часть глюкозы превращается в гликоген, который откладывается в печеночных клетках.
Итак, теперь мы с вами знаем, что основными ферментами, расщепляющими углеводы, являются амилаза, сахароза и лактоза. Причем более 90% удельного веса занимает амилаза. поскольку большая часть потребляемых нами углеводов являются сложными, то и амилаза соответственно - основной пищеварительный фермент, расщепляющий углеводы (сложные).
Белки. Белки пищи не усваиваются организмом, они не будут расщеплены в процессе переваривания пищи до стадии свободных аминокислот. Живой организм обладает способностью использовать вводимый с пищей белок только после его полного гидролиза в желудочно-кишечном тракте до аминокислот, из которых затем в клетках организма строятся свойственные для данного вида специфические белки.
Процесс переваривания белков и является многоступенчатым. Ферменты, расщепляющие белки называются “протиолитическими”. Примерно 95-97% белков пищи (те, что подверглись расщеплению) всасываются в кровь в виде свободных аминокислот.
Ферментный аппарат желудочно-кишечного тракта расщепляет пептидные связи белковых молекул поэтапно, строго избирательно. При отсоединении от белковой молекулы одной аминокислоты получается аминокислота и пептид. Затем от пептида отщепляется еще одна аминокислота, затем еще и еще. И так до тех пор, пока вся молекула не будет расщеплена до аминокислот.
Основной протеолитический фермент желудка - пепсин. Пепсин расщепляет крупные белковые молекулы до пептидов и аминокислот. Активен пепсин только в кислой среде, поэтому для его нормальной активности необходимо поддерживать определенный уровень кислотности желудочного сока. При некоторых заболеваниях желудка (гастрит и т.д.) кислотность желудочного сока значительно снижается.
В желудочном соке содержится также ренин. Это протеолитический фермент, который вызывает створаживание молока. Молоко в желудке человека должно сначала превращаться в кефир, а уж затем подвергаться дальнейшему усвоению. При отсутствии ренина (считается, что он присутствует в желудочном соке только до 10-13 летнего возраста) молоко не будет створоженным, проникает в толстый кишечник и там подвергается процессам гниения (лактаальбумины) и брожения (галактоза). Утешением служит тот факт, что у 70% взрослых людей функцию ренина берет на себя пепсин. 30% взрослых людей молоко все-таки не переносит. Оно вызывает у них вздутие кишечника (брожение галактозы) и послабление стула. Для таких людей предпочтительны кисломолочные продукты, в которых молоко находится уже в створоженном виде.
В 12-и перстной кишке пептиды и белки подвергаются уже более сильной “агрессии” протеолитичекими ферментами. Источником этих ферментов служит внешнесекреторный аппарат поджелудочной железы. Итак, 12-и перстная кишка содержит такие протеолитические ферменты, как трипсин, химотрипсин, коллагеназа, пептидаза, эластаза. А отличие от протеолитичеких ферментов желудка, ферменты поджелудочной железы разрывают большую часть пептидных связей и превращают основную массу пептидов в аминокислоты.
В тонком кишечнике полностью завершается распад еще имеющихся пептидов до аминокислот. Происходит всасывание основного количества аминокислот путем пассивного транспорта. Всасывание путем пассивного транспорта означает, что чем больше аминокислот будет находиться в тонком кишечнике, тем больше их всосется в кровь.
Тонкий кишечник содержит большой набор различных пищеварительных ферментов, которые объединяются под общим названием пептидазы. Здесь завершается в основном пищеварение белков.
Следы пищеварительных процессов можно отыскать еще и в толстом кишечнике, где под влиянием микрофлоры происходит частичный распад трудноперевариваемых молекул. Однако этот механизм носит рудиментарный характер и серьезного значения в общем процессе пищеварения не имеет.
Заканчивая рассказ о гидролизе белков, следует упомянуть, что все основные процессы пищеварения протекают на поверхности слизистой оболочки кишечника (пристеночное пищеварение по А.М.Уголеву).
Жиры (липиды). Слюна не содержит ферментов, расщепляющих жиры. В полости рта жиры не подвергаются никаким изменениям. Желудок человека содержит некоторое количество липазы. Липаза - фермент, расщепляющий жиры. В желудке человека, однако, липаза малоактивна из-за очень кислой желудочной среды. Только у грудных детей липаза расщепляет жиры грудного молока.
Расщепление жиров у взрослого человека происходит в основном в верхних отделах тонкого кишечника. Липаза не может воздействовать на жиры, если они не эмульгированы. Эмульгирование жиров происходит в 12-и перстной кишке, сразу же, как только туда попадает содержимое желудка. Основное эмульгирующее действие на жиры оказывают соли желчных кислот, которые попадают в 12-и перстную кишку из желчного пузыря. Желчные же кислоты синтезируются в печени из холестерина. Желчные кислоты не только эмульгируют жиры, но и активизируют липазу 12-и перстной кишки и кишечника. Эта липаза вырабатывается в основном внешнесекреторным аппаратом поджелудочной железы. Причем поджелудочная железа вырабатывает несколько видов липаз, которые расщепляют нейтральный мир на глицерин и свободные жирные кислоты.
Частично жиры в виде тонкой эмульсии могут всасываться в тонком кишечнике в неизменном виде, однако основная часть жира всасывается лишь после того, как липаза поджелудочной железы расщепит его на жирные кислоты и глицерин. Жирные кислоты с короткой цепью всасываются легко. Жирные же кислоты с длинной цепью всасываются плохо. Для всасывания им приходится соединиться с желчными кислотами, фосфолипидами и холестерином, образуя так называемые мицеллы - жировые шарики.
При необходимости ассимилировать большие, чем обычно, количества пищи и ликвидировать противоречие между потребностью организма в пищевых вещевых и способностью желудочно-кишечного тракта обеспечить эту потребность, чаще всего используют ведение извне фармакологических препаратов, содержащих пищеварительные ферменты.
Вопрос 106. Декарбоксилирование аминокислот. Строение кофермента декарбоксилаз. Биологическая роль образовавшихся соединений
Ответ. Процесс отщепления карбоксильной группы аминокислот в виде СО2 получил название декарбоксилирования. Несмотря на ограниченный круг аминокислот и их производных, подвергающихся декарбоксилированию в животных тканях, образующиеся продукты реакции – биогенные амины – оказывают сильное фармакологическое действие на множество физиологических функций человека и животных. В животных тканях установлено декарбоксилирование следующих аминокислот и их производных: тирозина, триптофана, 5-окситриптофана, валина, серина, гистидина, глу-таминовой и γ-оксиглутаминовой кислот, 3,4-диоксифенилаланина, цис-теина, аргинина, орнитина, S-аденозилметионина и α-аминомалоновой кислоты. Помимо этого, у микроорганизмов и растений открыто де-карбоксилирование ряда других аминокислот.
В живых организмах открыты 4 типа декарбоксилирования аминокислот:
1. α-Декарбоксилирование, характерное для тканей животных, при котором от аминокислот отщепляется карбоксильная группа, стоящая по соседству с α-углеродным атомом. Продуктами реакции являются СО2 и биогенные амины:
![]()
2. ω-Декарбоксилирование, свойственное микроорганизмам. Например, из аспарагиновой кислоты этим путем образуется α-аланин:
![]()
3. Декарбоксилирование, связанное с реакцией трансаминирования:
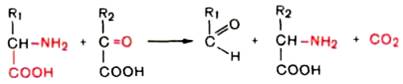
В этой реакции образуются альдегид и новая аминокислота, соответствующая исходной кетокислоте.
4. Декарбоксилирование, связанное с реакцией конденсации двух молекул:
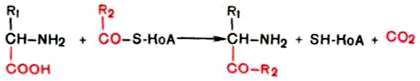
Эта реакция в тканях животных осуществляется при синтезе δ-амино-левулиновой кислоты из глицина и сукцинил-КоА и при синтезе сфинголипидов, а также у растений при синтезе биотина.
Реакции декарбоксилирования в отличие от других процессов промежуточного обмена аминокислот являются необратимыми. Они катализируются специфическими ферментами – декарбоксилазами аминокислот, отличающимися от декарбоксилаз α-кетокислот как белковым компонентом, так и природой кофермента. Декарбоксилазы аминокислот состоят из белковой части, обеспечивающей специфичность действия, и простетической группы, представленной пиридоксальфосфатом (ПФ), как и у трансаминаз.
Таким образом, в двух совершенно различных процессах обмена аминокислот участвует один и тот же кофермент. Исключение составляют две декарбоксилазы: гистидиндекарбоксилаза Micrococcus и Lactobacilus и аденозилметионин-декарбоксилаза Е. coli, содержащие вместо ПФ остаток пировиноградной кислоты.
Механизм реакции декарбоксилирования аминокислот в соответствии с общей теорией пиридоксалевого катализа сводится к образованию ПФ-субстратного комплекса, представленного, как и в реакциях трансаминирования, шиффовым основанием ПФ и аминокислоты:

Образование подобного комплекса в сочетании с некоторым оттягиванием электронов белковой частью молекулы фермента сопровождается лабилизацией одной из трех связей при α-углеродном атоме, благодаря чему аминокислота способна вступать в реакции трансаминирования (а), декарбоксилирования (b) и альдольного расщепления (с).
Далее представлены отдельные примеры декарбоксилирования аминокислот, в частности тех, продукты реакции которых оказывают сильное фармакологическое действие. Одним из хорошо изученных ферментов является декарбоксилаза ароматических аминокислот. Она не обладает строгой субстратной специфичностью и катализирует декарбок-силирование L-изомеров триптофана, 5-окситриптофана и 3,4-диоксифе-нилаланина (ДОФА); продуктами реакций, помимо СО2, являются соответственно триптамин, серотонин и диоксифенилэтиламин (дофамин).
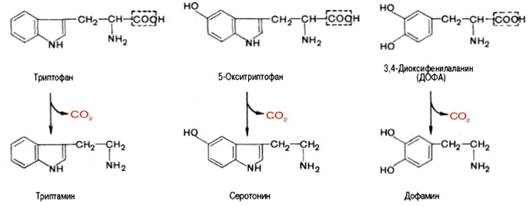
Декарбоксилаза ароматических аминокислот получена в чистом виде (мол. масса 112000), кофермент – ПФ. В больших количествах она содержится в надпочечниках и ЦНС, играет важную роль в регуляции содержания биогенных аминов. Образующийся из 5-окситриптофана серотонин оказался высокоактивным биогенным амином сосудосуживающего действия. Серотонин регулирует артериальное давление, температуру тела, дыхание, почечную фильтрацию и является медиатором нервных процессов в ЦНС. Некоторые авторы считают серотонин причастным к развитию аллергии, демпинг-синдрома, токсикоза беременных, карциноидного синдрома и геморрагических диатезов.
Продукт декарбоксилазной реакции дофамин является предшественником катехоламинов (норадреналина и адреналина). Источником ДОФА в организме является тирозин, который под действием специфической гидроксилазы превращается в 3,4-диоксифенилаланин. Тиро-зин-3-монооксигеназа открыта в надпочечниках, ткани мозга и периферической нервной системы. Простетической группой тирозин-моноокси-геназы, как и дофамин-монооксигеназы (последняя катализирует превращение дофамина в норадреналин) является тетрагидробиоптерин, имеющий следующее строение:
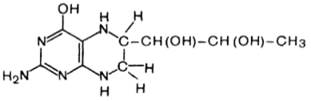
Физиологическая роль тирозин-3-монооксигеназы чрезвычайно велика, поскольку катализируемая этим ферментом реакция определяет скорость биосинтеза катехоламинов, регулирующих деятельность сердечно-сосудистой системы. В медицинской практике широко используются ингибиторы декарбоксилазы ароматических аминокислот, в частности α-метилдофа (альдомет), вызывающий снижение артериального давления.
В животных тканях с высокой скоростью протекает декарбоксилиро-вание гистидина под действием специфической декарбоксилазы.
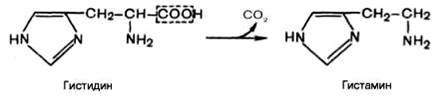
Гистамин оказывает широкий спектр биологического действия. По механизму действия на кровеносные сосуды он резко отличается от других биогенных аминов, так как обладает сосудорасширяющим свойством. Большое количество гистамина образуется в области воспаления, что имеет определенный биологический смысл. Вызывая расширение сосудов в очаге воспаления, гистамин тем самым ускоряет приток лейкоцитов, способствуя активации защитных сил организма. Кроме того, гистамин участвует в секреции соляной кислоты в желудке, что широко используется в клинике при изучении секреторной деятельности желудка (гистаминовая проба). Он имеет прямое отношение к явлениям сенсибилизации и десенсибилизации. При повышенной чувствительности к гистамину в клинике используют антигистаминные препараты (санорин, димедрол и др.), оказывающие влияние на рецепторы сосудов. Гистамину приписывают также роль медиатора боли. Болевой синдром – сложный процесс, детали которого пока не выяснены, но участие в нем гистамина не подлежит сомнению.
В клинической практике широко используется, кроме того, продукт α-декарбоксилирования глутаминовой кислоты – γ-аминомасляная кислота (ГАМК). Фермент, катализирующий эту реакцию (глутаматдекарбокси-лаза), является высокоспецифичным.
![]()
Интерес к ГАМК объясняется ее тормозящим действием на деятельность ЦНС. Больше всего ГАМК и глутаматдекарбоксилазы обнаружено в сером веществе коры большого мозга, в то время как белое вещество мозга и периферическая нервная система их почти не содержат. Введение ГАМК в организм вызывает разлитой тормозной процесс в коре (центральное торможение) и у животных приводит к утрате условных рефлексов. ГАМК используется в клинике как лекарственное средство при некоторых заболеваниях ЦНС, связанных с резким возбуждением коры большого мозга. Так, при эпилепсии хороший эффект (резкое сокращение частоты эпилептических припадков) дает введение глутаминовой кислоты. Как оказалось, лечебный эффект обусловлен не самой глутаминовой кислотой, а продуктом ее декарбоксилирования – ГАМК.
В животных тканях с высокой скоростью декарбоксилируются также два производных цистеина – цистеиновая и цистеинсульфиновая кислоты. В процессе этих специфических ферментативных реакций образуется таурин, который используется в организме для синтеза парных желчных кислот.
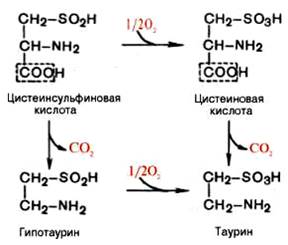
Следует указать еще на два недавно открытых в тканях животных фермента, катализирующих декарбоксилирование орнитина и S-аденозил-метионина: орнитиндекарбоксилазу и аденозилметиониндекарбоксилазу.
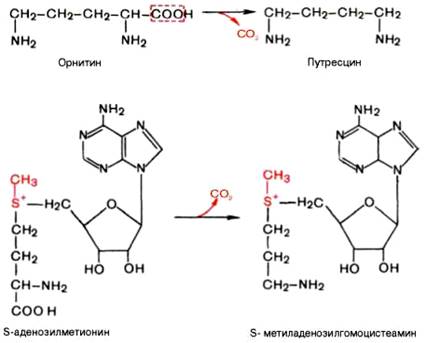
Значение этих реакций для тканей животных огромно, поскольку продукты реакций используются для синтеза полиаминов – спермидина и спермина.
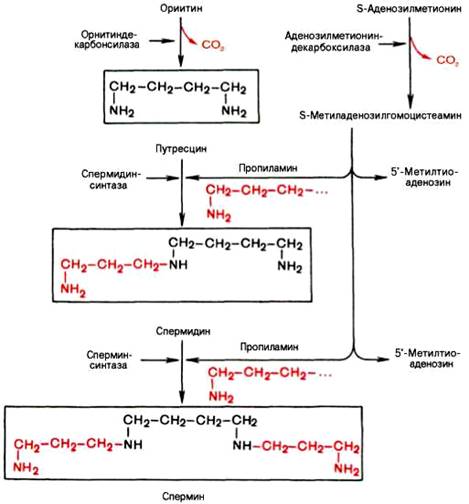
Полиамины, к которым относят также диамин путресцин, играют важную роль в процессах клеточного роста и дифференцировки, в регуляции синтеза ДНК, РНК и белка, стимулируя транскрипцию и трансляцию, хотя конкретный механизм участия их в указанных процессах не всегда ясен.
Таким образом, биогенные амины являются сильными фармакологически активными веществами, оказывающими разностороннее влияние на физиологические функции организма. Некоторые биогенные амины нашли широкое применение в качестве лекарственных препаратов.
Распад биогенных аминов. Накопление биогенных аминов может отрицательно сказываться на физиологическом статусе и вызывать ряд существенных нарушений функций в организме. Однако органы и ткани, как и целостный организм, располагают специальными механизмами обезвреживания биогенных аминов, которые в общем виде сводятся к окислительному дезаминированию этих аминов с образованием соответствующих альдегидов и освобождением аммиака:
![]()
Ферменты, катализирующие эти реакции, получили название моноамин-и диаминоксидаз. Более подробно изучен механизм окислительного дез-аминирования моноаминов. Этот ферментативный процесс является необратимым и протекает в две стадии:
R-CH2-NH2+ Е-ФАД + H20-> R-CHO + NH3+ Е-ФАДН2 (1)
Е-ФАДН2 +02-> Е-ФАД + Н202 (2)
Первая (1), анаэробная, стадия характеризуется образованием альдегида, аммиака и восстановленного фермента. Последний в аэробной фазе окисляется молекулярным кислородом. Образовавшаяся перекись водорода далее распадается на воду и кислород. Моноаминоксидаза (МАО), ФАД-содержащий фермент, преимущественно локализуется в митохондриях, играет исключительно важную роль в организме, регулируя скорость биосинтеза и распада биогенных аминов. Некоторые ингибиторы моно-аминоксидазы (ипраниазид, гармин, паргилин) используются при лечении гипертонической болезни, депрессивных состояний, шизофрении и др.
Вопрос 108. Напишите орнитиновый цикл мочевинообразования. Укажите локализацию процесса
Ответ. Основным механизмом обезвреживания аммиака в организме является биосинтез мочевины. Последняя выводится с мочой в качестве главного конечного продукта белкового, соответственно аминокислотного, обмена. На долю мочевины приходится до 80–85% от всего азота мочи. Основным и, возможно, единственным местом синтеза мочевины является печень. Впервые Г. Кребс и К. Гензеляйт в 1932 г. вывели уравнения реакций синтеза мочевины, которые представлены в виде цикла, получившего в литературе название орнитинового цикла мочевинообразования Кребса. Следует указать, что в биохимии это была первая циклическая система метаболизма, описание которой почти на 5 лет опеределило открытие Г. Кребсом другого метаболического процесса – цикла трикарбоновых кислот. Дальнейшие исследования в основном подтвердили циклический характер биосинтеза мочевины в печени. Благодаря исследованиям Г. Коена, С. Ратнер и сотр. были уточнены промежуточные этапы и ферментные системы, катализирующие образование мочевины.
Таким образом, весь цикл мочевинообразования может быть представлен следующим образом. На первом этапе синтезируется макроэрги-ческое соединение карбамоилфосфат – метаболически активная форма аммиака, используемая в качестве исходного продукта для синтеза пи-римидиновых нуклеотидов (соответственно ДНК и РНК) и аргинина (соответственно белка и мочевины):
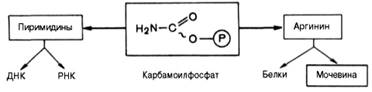
К настоящему времени открыты три разных пути синтеза карбамоил-фосфата de novo, катализируемые тремя разными ферментами. Первую необратимую реакцию катализирует регуляторный фермент – аммиакзави-симая карбамоилфосфатсинтетаза (КФ 6.3.4.16):
![]()
Реакция требует затраты двух молекул АТФ, открыта в митохондриях клеток печени и используется преимущественно для синтеза аргинина и мочевины. В этой реакции в качестве активного стимулирующего ал-лостерического эффектора действует N-ацетилглутамат.
Вторую, также необратимую, реакцию катализирует глутаминзависимая карбамоилфосфатсинтетаза (КФ 6.3.5.5):
![]()
Данная реакция открыта в цитозоле клеток животных и требует наличия ионов Mg2+. Следует указать, что благодаря включению гидролитической стадии она используется преимущественно для синтеза пиримидиновых нуклеотидов (см. далее). Фермент широко распространен в клетках животных.
Третью обратимую реакцию катализирует карбаматкиназа (КФ 2.7.2.2):
![]()
Реакция открыта у разных микроорганизмов и, возможно, используется скорее для ресинтеза АТФ, чем для синтеза карбамоилфосфата.
На втором этапе цикла мочевинообразования происходит конденсация карбамоилфосфата и орнитина с образованием цитруллина; реакцию катализирует орнитин-карбамоилтрансфераза (КФ 2.1.3.3).
На следующей стадии цитруллин превращается в аргинин в результате двух последовательно протекающих реакций. Первая из них, энергозави-симая,– это конденсация цитруллина и аспарагиновой кислоты с образованием аргининосукцината (эту реакцию катализирует аргининосукцинат-синтетаза). Аргининосукцинат распадается в следующей реакции на аргинин и фумарат при участии другого фермента – аргининосукцинатлиазы. На последнем этапе аргинин расщепляется на мочевину и орнитин под действием аргиназы.
Необходимо подчеркнуть, что аргиназа содержится в печени тех животных, которые экскретируют с мочой мочевину как основной и конечный продукт азотистого обмена. В печени птиц, например, аргиназа отсутствует, поскольку птицы вместо мочевины выделяют мочевую кислоту. Орни-тиновый цикл мочевинообразования с учетом новых данных представлен на рис. 6.
Суммарная реакция синтеза мочевины без учета всех промежуточных продуктов может быть представлена в следующем виде:

Данная реакция сопровождается снижением свободной энергии (ΔG0 = –40 кДж), поэтому процесс всегда протекает в направлении синтеза мочевины. Следует указать, что синтез мочевины энергетически дорого обходится организму.
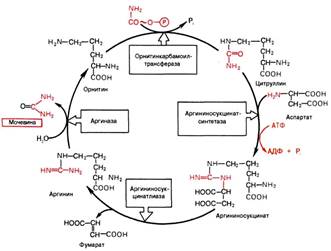
Рис. 6. Орнитиновый цикл синтеза мочевины в печени.
На синтез одной молекулы мочевины требуется затрата четырех высокоэнергетических фосфатных групп: две молекулы АТФ расходуются на синтез карбамоилфосфата и одна – на образование аргининоянтарной кислоты, при этом АТФ расщепляется на АМФ и РРi, который при гидролизе также образует две молекулы Рi.
Из приведенной схемы процесса мочевинообразования нетрудно видеть, что один из атомов азота мочевины имеет своим источником свободный аммиак (через карбамоилфосфат); второй атом азота поступает из ас-партата. Аммиак образуется главным образом в процессе глутаматде-гидрогеназной реакции. В процессе пополнения запасов аспартата участвуют три сопряженные реакции: сначала фумарат под действием фумаразы присоединяет воду и превращается в малат, который окисляется при участии малатдегидрогеназы с образованием оксалоацетата; последний в реакции трансаминирования с глутаматом вновь образует аспартат.
Учитывая известные фактические данные о механизмах обезвреживания аммиака в организме, можно сделать следующее заключение. Часть аммиака используется на биосинтез аминокислот путем восстановительного аминирования α-кетокислот по механизму реакции трансаминирования. Аммиак связывается при биосинтезе глутамина и аспарагина. Некоторое количество аммиака выводится с мочой в виде аммонийных солей. В форме креатинина, который образуется из креатина и креатинфосфата, выделяется из организма значительная часть азота аминокислот. Наибольшее количество аммиака расходуется на синтез мочевины, которая выводится с мочой в качестве главного конечного продукта белкового обмена в организме человека и животных. Подсчитано, что в состоянии азотистого равновесия организм взрослого здорового человека потребляет и соответственно выделяет примерно 15 г азота в сутки; из экскретируемого с мочой количества азота на долю мочевины приходится около 85%, креатинина – около 5%, аммонийных солей – 3%, мочевой кислоты – 1% и на другие формы – около 6%.
В процессе эволюции живые организмы выработали различные типы азотистого обмена. Это аммониотелический тип, при котором главным конечным продуктом азотистого обмена является аммиак; он свойствен преимущественно рыбам. При уреотелическом типе обмена основным конечным продуктом обмена белков является мочевина; такой тип характерен для человека и животных. Урикотелический тип характерен для птиц и рептилий; главным конечным продуктом данного типа обмена является мочевая кислота.
Вопрос 121. Через какие активные соединения клетки гормоны реализуют свое действие?
Ответ. Высшей формой гуморальной регуляции является гормональная. Термин "гормон" был впервые применен в 1902 г. Старлингом и Бейлиссом в отношении открытого ими вещества, продуцирующегося в двенадцатиперстной кишке, - секретина. Термин "гормон" в переводе с греческого означает "побуждающий к действию", хотя не все гормоны обладают стимулирующим эффектом.
Гормоны - это биологически высокоактивные вещества, синтезирующиеся и выделяющиеся во внутреннюю среду организма эндокринными железами, или железами внутренней секреции, и оказывающие регулирующее влияние на функции удаленных от места их секреции органов и систем организма. Эндокринная железа - это анатомическое образование, лишенное выводных протоков, единственной или основной функцией которого является внутренняя секреция гормонов. К эндокринным железам относятся гипофиз, эпифиз, щитовидная железа, надпочечники (мозговое и корковое вещество), паращитовидные железы.
В отличие от внутренней секреции, внешняя секреция осуществляется экзокринными железами через выводные протоки во внешнюю среду. В некоторых органах одновременно присутствуют оба типа секреции. Инкреторная функция осуществляется эндокринной тканью, т.е. скоплением клеток с инкреторной функцией в органе, обладающем функциями, не связанными с продукцией гормонов. К органам со смешанным типом секреции относятся поджелудочная железа и половые железы. Одна и та же железа внутренней секреции может продуцировать неодинаковые по своему действию гормоны. Так, например, щитовидная железа продуцирует тироксин и тирокальцитонин. В то же время продукция одних и тех же гормонов может осуществляться разными эндокринными железами. Например, половые гормоны продуцируются и половыми железами, и надпочечниками.
Продукция биологически активных веществ - это функция не только желез внутренней секреции, но и других традиционно неэндокринных органов: почек, желудочно-кишечного тракта, сердца. Не все вещества, образующиеся специфическими клетками этих органов, удовлетворяют классическим критериям понятия "гормоны". Поэтому наряду с термином "гормон" в последнее время используются также понятия гормоноподобные и биологически активные вещества (БАВ), гормоны местного действия. Так, например, некоторые из них синтезируются так близко к своим органам-мишеням, что могут достигать их диффузией, не попадая в кровоток. Клетки, вырабатывающие такие вещества, называют паракринными. Трудность точного определения термина "гормон" особенно хорошо видна на примере катехоламинов - адреналина и норадреналина. Когда рассматривается их выработка в мозговом веществе надпочечников, их обычно называют гормонами, если речь идет об их образовании и выделении симпатическими окончаниями, их называют медиаторами.
Регуляторные гипоталамические гормоны - группа нейропептидов, включая недавно открытые энкефалины и эндорфины, действуют не только как гормоны, но и выполняют своеобразную медиаторную функцию. Некоторые из регуляторных гипоталамических пептидов обнаружены не только в нейронах головного мозга, но и в особых клетках других органов, например кишечника: это вещество Р, нейротензин, соматостатин, холецистокинин и др. Клетки, вырабатывающие эти пептиды, образуют согласно современным представлениям диффузную нейроэндокринную систему, состоящую из разбросанных по разным органам и тканям клеток.
Клетки этой системы характеризуются высоким содержанием аминов, способностью к захвату предшественников аминов и наличием декарбоксилазы аминов. Отсюда название системы по первым буквам английских слов Amine Precursors Uptake and Decarboxylating system - APUD-система - система захвата предшественников аминов и их декарбоксилирования. Поэтому правомерно говорить не только об эндокринных железах, но и об эндокринной системе, которая объединяет все железы, ткани и клетки организма, выделяющие во внутреннюю среду специфические регуляторные вещества.
Химическая природа гормонов и биологически активных веществ различна. От сложности строения гормона зависит продолжительность его биологического действия, например, от долей секунды у медиаторов и пептидов до часов и суток у стероидных гормонов и йодтиронинов. Анализ химической структуры и физико-химических свойств гормонов помогает понять механизмы их действия, разрабатывать методы их определения в биологических жидкостях и осуществлять их синтез.
Классификация гормонов и БАБ по химической структуре:
Производные аминокислот: производные тирозина: тироксин, трийодтиронин, дофамин, адреналин, норадреналин; производные триптофана: мелатонин, серотонин; производные гистидина: гистамин.
Белково-пептидные гормоны: полипептиды: глюкагон, кортикотропин, меланотропин, вазо-прессин, окситоцин, пептидные гормоны желудка и кишечника; простые белки (протеины): инсулин, соматотропин, пролактин, паратгормон, кальцитонин; сложные белки (гликопротеиды): тиреотропин, фоллитропин, лютропин.
Стероидные гормоны: кортикостероиды (альдостерон, кортизол, кортикостерон); половые гормоны: андрогены (тестостерон), эстрогены и прогестерон.
Производные жирных кислот: арахидоновая кислота и ее производные: простагландины, простациклины, тромбоксаны, лейкотриены.
Несмотря на то, что гормоны имеют разное химическое строение, для них характерны некоторые общие биологические свойства.
Общие свойства гормонов:
1. Строгая специфичность (тропность) физиологического действия.
2. Высокая биологическая активность: гормоны оказывают свое физиологическое действие в чрезвычайно малых дозах.
3. Дистантный характер действия: клетки-мишени располагаются обычно далеко от места образования гормона.
4. Многие гормоны (стероидные и производные аминокислот) не имеют видовой специфичности.
5. Генерализованность действия.
6. Пролонгированность действия.
Установлены четыре основных типа физиологического действия на организм: кинетическое, или пусковое, вызывающее определенную деятельность исполнительных органов; метаболическое (изменения обмена веществ); морфогенетическое (дифференциация тканей и органов, действие на рост, стимуляция формообразовательного процесса); корригирующее (изменение интенсивности функций органов и тканей).
Гормональный эффект опосредован следующими основными этапами: синтезом и поступлением в кровь, формами транспорта, клеточными механизмами действия гормонов. От места секреции гормоны доставляются к органам-мишеням циркулирующими жидкостями: кровью, лимфой. В крови гормоны циркулируют в нескольких формах: 1) в свободном состоянии; 2) в комплексе со специфическими белками плазмы крови; 3) в форме неспецифического комплекса с плазменными белками; 4) в адсорбированном состоянии на форменных элементах крови. В состоянии покоя 80% приходится на комплекс со специфическими белками. Биологическая активность определяется содержанием свободных форм гормонов. Связанные формы гормонов являются как бы депо, физиологическим резервом, из которого гормоны переходят в активную свободную форму по мере необходимости.
Обязательным условием для проявления эффектов гормона является его взаимодействие с рецепторами. Гормональные рецепторы представляют собой особые белки клетки, для которых характерны: 1) высокое сродство к гормону; 2) высокая избирательность; 3) ограниченная связывающая емкость; 4) специфичность локализации рецепторов в тканях. На одной и той же мембране клетки могут располагаться десятки разных типов рецепторов. Количество функционально активных рецепторов может изменяться при различных состояниях и в патологии. Так, например при беременности в миометрии исчезают М-холинорецепторы, и возрастает количество окситоциновых рецепторов. При некоторых формах сахарного диабета имеет место функциональная недостаточность инсулярного аппарата, т.е. уровень инсулина в крови высокий, но часть инсулиновых рецепторов оккупирована аутоантителами к этим рецепторам. В 50% случаев рецепторы локализуются на мембранах клетки-мишени; 50% - внутри клетки.
Механизмы действия гормонов. Существуют два основных механизма действия гормонов на уровне клетки: реализация эффекта с наружной поверхности клеточной мембраны и реализация эффекта после проникновения гормона внутрь клетки.
В первом случае рецепторы расположены на мембране клетки. В результате взаимодействия гормона с рецептором активируется мембранный фермент - аденилатциклаза. Этот фермент способствует образованию из аденозинтрифосфорной кислоты (АТФ) важнейшего внутриклеточного посредника реализации гормональных эффектов - циклического 3,5-аденозинмонофос-фата (цАМФ). цАМФ активирует клеточный фермент протеинкиназу, реализующую действие гормона. Установлено, что гормонозависимая аденилатциклаза - это общий фермент, на который действуют различные гормоны, в то время как рецепторы гормонов множественны и специфичны для каждого гормона. Вторичными посредниками кроме цАМФ могут быть циклический 3,5-гуанозинмонофосфат (цГМФ), ионы кальция, инозитол-трифосфат. Так действуют пептидные, белковые гормоны, производные тирозина - катехоламины. Характерной особенностью действия этих гормонов является относительная быстрота возникновения ответной реакции, что обусловлено активацией предшествующих уже синтезированных ферментов и других белков.
Во втором случае рецепторы для гормона находятся в цитоплазме клетки. Гормоны этого механизма действия в силу своей липофильности легко проникают через мембрану внутрь клетки-мишени и связываются в ее цитоплазме специфическими белками-рецепторами. Гормон-рецепторный комплекс входит в клеточное ядро. В ядре комплекс распадается, и гормон взаимодействует с определенными участками ядерной ДНК, следствием чего является образование особой матричной РНК. Матричная РНК выходит из ядра и способствует синтезу на рибосомах белка или белка-фермента. Так действуют стероидные гормоны и производные тирозина - гормоны щитовидной железы. Для их действия характерна глубокая и длительная перестройка клеточного метаболизма.
Инактивация гормонов происходит в эффекторных органах, в основном в печени, где гормоны претерпевают различные химические изменения путем связывания с глюкуроновой или серной кислотой либо в результате воздействия ферментов. Частично гормоны выделяются с мочой в неизмененном виде. Действие некоторых гормонов может блокироваться благодаря секреции гормонов, обладающих антагонистическим эффектом.
Гормоны выполняют в организме следующие важные функции:
1. Регуляция роста, развития и дифференцировки тканей и органов, что определяет физическое, половое и умственное развитие.
2. Обеспечение адаптации организма к меняющимся условиям существования.
3. Обеспечение поддержания гомеостаза.
Функциональная классификация гормонов:
1. Эффекторные гормоны - гормоны, которые оказывают влияние непосредственно на орган-мишень.
2. Тройные гормоны - гормоны, основной функцией которых является регуляция синтеза и выделения эффекторных гормонов. Выделяются аденогипофизом.
3. Рилизинг-гормоны - гормоны, регулирующие синтез и выделение гормонов аденогипофиза, преимущественно тройных. Выделяются нервными клетками гипоталамуса.
Виды взаимодействия гормонов. Каждый гормон не работает в одиночку. Поэтому необходимо учитывать возможные результаты их взаимодействия.
Синергизм - однонаправленное действие двух или нескольких гормонов. Например, адреналин и глюкагон активируют распад гликогена печени до глюкозы и вызывают увеличение уровня сахара в крови.
Антагонизм всегда относителен. Например, инсулин и адреналин оказывают противоположные действия на уровень глюкозы в крови. Инсулин вызывает гипогликемию, адреналин - гипергликемию. Биологическое же значение этих эффектов сводится к одному - улучшению углеводного питания тканей.
Пермиссивное действие гормонов заключается в том, что гормон, сам не вызывая физиологического эффекта, создает условия для ответной реакции клетки или органа на действие другого гормона. Например, глюкокортикоиды, не влияя на тонус мускулатуры сосудов и распад гликогена печени, создают условия, при которых даже небольшие концентрации адреналина увеличивают артериальное давление и вызывают гипергликемию в результате гликогенолиза в печени.
Вопрос 126. Охарактеризуйте механизм действия адреналина и глюкагона на липидный обмен
Ответ. Обмен липидов регулируется ЦНС. Кора большого мозга оказывает трофическое влияние на жировую ткань либо через нижележащие отделы ЦНС – симпатическую и парасимпатическую системы, либо через эндокринные железы. В настоящее время установлен ряд биохимических механизмов, лежащих в основе действия гормонов на липидный обмен.
Известно, что длительный отрицательный эмоциональный стресс, сопровождающийся увеличением выброса катехоламинов в кровяное русло, может вызвать заметное похудание. Уместно напомнить, что жировая ткань обильно иннервируется волокнами симпатической нервной системы, возбуждение этих волокон сопровождается выделением норадреналина непосредственно в жировую ткань. Адреналин и норадреналин увеличивают скорость липолиза в жировой ткани; в результате усиливается мобилизация жирных кислот из жировых депо и повышается содержание неэстерифи-цированных жирных кислот в плазме крови. Как отмечалось, тканевые липазы (триглицеридлипаза) существуют в двух взаимопревращающихся формах, одна из которых фосфорилирована и каталитически активна, а другая – нефосфорилирована и неактивна. Адреналин стимулирует через аденилатциклазу синтез цАМФ. В свою очередь цАМФ активирует соответствующую протеинкиназу, которая способствует фосфорилированию липазы, т.е. образованию ее активной формы. Следует заметить, что действие глюкагона на липолитическую систему сходно с действием кате-холаминов.
Не подлежит сомнению, что секрет передней доли гипофиза, в частности соматотропный гормон, оказывает влияние на липидный обмен. Гипофункция железы приводит к отложению жира в организме, наступает гипофизарное ожирение. Напротив, повышенная продукция СТГ стимулирует липолиз, и содержание жирных кислот в плазме крови увеличивается. Доказано, что стимуляция липолиза СТГ блокируется ингибиторами синтеза мРНК. Кроме того, известно, что действие СТГ на липолиз характеризуется наличием лаг-фазы продолжительностью около 1 ч, тогда как адреналин стимулирует липолиз почти мгновенно. Иными словами, можно считать, что первичное действие этих двух типов гормонов на липолиз проявляется различными путями. Адреналин стимулирует активность аденилатциклазы, а СТГ индуцирует синтез данного фермента. Конкретный механизм, с помощью которого СТГ избирательно увеличивает синтез аденилатциклазы, пока неизвестен.
Вопрос 130. Клинико-диагностическое значение определения активности амилазы сыворотки крови и мочи
Ответ. У человека альфа-амилаза секретируется поджелудочной и слюнной железами, небольшая ее активность обнаруживается в тканях печени и скелетной мускулатуры. Молекулярная масса альфа-амилазы относительно низка, в отличие от большинства ферментов она фильтруется в клубочках почек и содержится в моче. Альфа-амилаза состоит из двух изоферментов: панкреатического типа (Р - тип) и слюнного (S ? тип). У здоровых людей в сыворотке крови приходится около 70% амилолитической активности приходится на слюнной изофермент, в моче приблизительно такой же процент приходится на панкреатическую изоамилазу.
Гиперамилаземия и гиперамилазурия наблюдаются при многих заболеваниях, но наиболее выражены при остром панкреатите, при котором активность увеличивается в основном (до 90% и более) за счет панкреатического изофермента. При данном заболевании наибольший объем содержания амилазы в крови и моче отмечен в первые 1-3 сут. Гиперамилазурию панкреатического происхождения вызывают также вирусный гепатит, рак поджелудочной железы.
К гиперамилаземии непанкреатического происхождения относят поражение слюнных желез, почечную недостаточность. Причинами повышения α-амилазы в крови являются нарушение секреции желез, содержащих α-амилазу, недостаточность выделения почками амилазы из организма.
Гиперамилаземию вызывают многие фармакологические вещества, кортикостероидные препараты, салицилаты, тетрациклин, фуросемид, гистамин. Для α-амилазы крови характерны широкие внутри- и межиндивидуальные вариации. Наиболее информативным является определение панкреатической изоамилазы.
Изменение активности альфа-амилазы при патологии.
Определение активности альфа-амилазы приобретает клинико-диагностическое значение при диагностике и мониторинге заболеваний поджелудочной железы, пищеварительной системы, поражения органов брюшной полости. В ряде случаев (например, при заболеваниях поджелудочной железы) диагностическое значение имеет активность только одного изофермента - панкреатической альфа-амилазы; при мониторинге заболеваний пищеварительного тракта необходимо проводить сравнение активности фермента в сыворотке (секреторная активность и дополнительное всасывание из органов брюшной полости) и моче (зачастую степень амилазурии является более стабильным и длительно сохраняющимся показателем, чем амилаземия).
Повышенные значения активности альфа-амилазы наблюдаются при:
Заболеваниях поджелудочной железы воспалительного характера (острый, отечный, хронический, реактивный панкреатит). Гиперамилаземия обычно носит острый (повышение в 10-40 раз), но зачастую кратковременный характер, гиперамилазурия также значительна, но уровень альфа-амилазы в моче снижается гораздо медленнее, чем в плазме (сыворотке) крови. Это свойство удобно использовать при "запоздалом" диагностическом обследовании пациента с клиническими признаками панкреатита, однако для мониторинга течения процесса обычно измеряют активность фермента в обеих биологических жидкостях. Следует отметить, что тяжелые формы поражения ПЖ с угрозой некроза тканей, а также панкреатит, сопровождающийся серьезными нарушениями липидного обмена, могут сопровождаться реальным или ложным (за счет ингибирования амилазы триацилглицеридами) снижением активности фермента. Поэтому при подозрении на панкреатит любой формы диагностически важно не только определять активность альфа-амилазы в моче и сыворотке крови, но и рассчитывать амилазокреатининовый клиренс по формуле:
[(Активность альфа-амилазы в моче х Креатинин сыворотки крови)/(Креатинин мочи х Активность альфа-амилазы сыворотки крови)]*100%.
При остром панкреатите эта расчетная величина превышает 6%. Расчет клиренса позволяет не делать поправку на объем и время сбора мочи и делает результат менее зависимым от используемых способов определения.
Заболеваниях органов брюшной полости, имеющих сходные симптомы с острым панкреатитом: остром аппендиците, перитоните, перфоративной язве желудка и двенадцатиперстной кишки. Активность альфа-амилазы увеличивается примерно в 3-5, редко - в 10 раз, основной причиной гиперамилазиемии является не столько повышенная секреция фермента, сколько вторичное всасывание ее из органов брюшной полости в кровь. Для дифференцирования диагноза проводят определение альфа-амилазы и креатинина в моче и расчет клиренса по креатинину с использованием приведенной выше формулы. При постоянном значении клиренса от 1 до 4 % следует заподозрить именно заболевания органов брюшной полости, маскирующихся под острый панкреатит.
По той же причине активность альфа-амилазы достоверно повышается при перфорации пищевода, гастрите, острой кишечной непроходимости, ишемии и инфаркте тонкой кишки, разрыве маточных труб, сальпингите, аневризме аорты.
При заболеваниях внутренних органов, не связанных напрямую со системой пищеварения, возможно умеренное повышение активности общей альфа-амилазы и/или ее изоферментов. К таким заболеваниям относятся: холецистит, холелитиаз, поражение паренхимы почек и воспалительные заболевания этого органа, почечная недостаточность, простатит, диабетический кетоацидоз, метастазирование злокачественных опухолей в ткани и органы (в особенности, в легкие, поджелудочную железу, поперечно-ободочную кишку). При подозрении на описанные выше патологии проводят сравнительный анализ общей и панкреатической альфа-амилазы, поскольку при описанных заболеваниях в основном повышается активность селиварного изофермента (при онкологическом поражении ПЖ и поперечно-ободочной кишки повышается активность обоих - P- и S-изоферментов, что позволяет отличить его от воспалительных заболеваний тех же органов).
Гиперамилаземия непанкреатического происхождения возникает также при заболеваниях слюнных желез и некоторых патологиях мозга. В этом случае повышение общей альфа-амилазной активности также возникает за счет увеличения активности слюнного изофермента и для дифференцирования диагноза следует провести параллельное измерение общей и панкреатической альфа-амилазы (при нормальной активности Р-амилазы можно заподозрить указанную выше патологию).
Сравнительный анализ активности альфа-амилазы в моче и сыворотке крови является также ценным диагностическим параметром при оценке патологии почек. При поражении функции почечного фильтра (при нефротическом синдроме, гломерулонефритах и др.) активность альфа-амилазы в моче резко снижена, тогда как в крови отмечается умеренная или значительная гиперамилаземия, поэтому коэффициент "Амилаза сыворотки крови / амилаза мочи" является одним из основных диагностических параметров при данной патологии.
Послеоперационные состояния (системный воспалительный ответ на хирургическое вмешательство, развитие ОРДС и др.) также могут явиться причиной умеренной гиперамилаземии.
Понижение активности альфа-амилазы обычно сопутствует понижению функциональной активности поджелудочной железы, вызванной поражением паренхимы органа и/или нарушением экзокринной регуляции, и чаще всего возникает при:
Некротических изменениях в поджелудочной железе, связанных с гемморагическим панкреатитом и тотальным панкреонекрозом.
При заболеваниях печени (гепатиты, цирроз). Злокачественных опухолях, в особенности - при метастазировании печени.
Тяжелых нарушениях эндокринного баланса: гипотиреозе, сахарном диабете, гиперлипемии (особенно, с избытком триацилглицеридов, за счет ингибрования активного центра фермента), при общем расстройстве питания, кахексии, при токсикозе беременных.
В ряде случаев при описанных выше патологиях сывороточная активность альфа-амилазы может не снизиться, а остаться неизменной.
Основные соотношения при дифференциальной диагностике патологий со сходными симптомами.
Наиболее распространенные коэффициенты соотношений :
Общая альфа-амилаза /панкреатическая составляющая. Используется при дифференциальной диагностике воспалительных и злокачественных процессов в органах брюшной полости и некоторых других тканей. При воспалительных процессах, не задевающих поджелудочную железу, основным источником активности общей альфа-амилазы является селиварный фермент, поэтому соотношение "Общая амилаза/панкреатическая изоформа" > 1. При злокачественном поражении органов брюшной полости вклад обоих изоферментов примерно одинаков, и указанный коэффициент приблизительно равен 1,8-2,4. При вовлечении ПЖ в воспалительный процесс или при наличии в ней метастазов опухоли соотношение активности общей и панкреатической амилазы не превышает 1,5.
Общая альфа-амилаза/креатинин в моче и в сыворотке крови. Используется расчетный клиренс (формула приведена в пункте 3), позволяет дифференцировать воспалительный процесс в ПЖ (коэффициент превышает 6%) и органах брюшной полости (1-4%).
Общая альфа-амилаза сыворотки крови / Общая альфа-амилаза мочи. Используется при оценке функциональной активности почечного фильтра. При нефротическом синдроме и гломерулонефритах коэффициент выше 2-2,5.
Общая альфа-амилаза сыворотки крови / Триглицериды сыворотки крови. Соотношение носит "обратный" характер, поскольку избыток триацилглицеридов может ингибировать активность альфа-амилазы. Позволяет дифференцировать патологическое снижение продукции фермента и интерференцию с гиперипемией.
Общая альфа-амилаза сыворотки крови/ Панкреатическая липаза. Биологический и диагностический смысл коэффициента аналогичен таковому у предыдущего соотношения, поскольку наличие свободных триацилглицеридов в сыворотке крови напрямую связано с деятельностью панкреатической липазы. Пропорция прямая.
Изменение активности альфа-амилазы у практически здоровых людей. Интерференция с лекарственными препаратами.
Активность альфа-амилазы в плазме крови здоровых людей существенно зависит от характера питания и превалирующих в диете продуктов. Кроме того, этот параметр сильно зависит от интенсивности обменных процессов у конкретного индивидуума, а также от мозговой активности обследуемого. Поэтому активность фермента у одного и того же индивидуума в различные сроки (даже в течение одних и тех же суток) может варьировать в пределах 25-50% в обе стороны.
Гиперамилазиемия у здоровых людей может быть вызвана эмоциональным состоянием (стресс, усиление умственной активности), употреблением (даже однократным) алкоголя, а также связан с диетой, обогащенной белком и обедненной углеводами и сахарами. Гипоамилаземия возникает при увеличении утилизации глюкозы, при голодании, активной физической нагрузке, а также у людей с гиперлипидемией даже в отсутствие ее клинических проявлений.
При беременности сроком до 19 недель активность альфа-амилазы понижена в сравнении с нормой, в сроки до 33-34 недели беременности уровень активности повышается, а к 37-40 неделе стабилизируется на уровне, на 10-20% превышающем активность фермента у здоровых небеременных женщин.
На активность альфа-амилазы и ее панкреатического изофермента существенное влияние оказывает также применение целого ряда распространенных лекарственных препаратов, употребляемых практически здоровыми людьми при возникновении преходящих заболеваний инфекционного и воспалительного характера - например, антибиотиков тетрациклинового ряда, фуросемида, кортикостероидных противовоспалительных препаратов, салицилатов.
Также, значительное влияние на активность альфа-амилазы оказывают препараты, приводящие к сокращению сфинктера Одди, в том числе - гормоны (адреналин, гистамин, секретин), наркотические вещества (морфин, пантопон, опий, кодеин) и другие препараты, повышающие секрецию фермента и подавляющие отток секрета из поджелудочной железы.
Вопрос 133. Нарушение прямого и соотношения непрямого билирубина при желтухах. Охарактеризуйте гемолитическую, паренсиматозную и обтурационную желтухи
Ответ. Желтухой называется синдром, развивающийся вследствие накопления в крови избыточного количества билирубина. В клинике он диагностируется по окрашиванию кожи и слизистой в различные оттенки желтого цвета.
Интенсивность желтухи зависит от кровоснабжения органа или ткани. В начале обнаруживается желтое окрашивание склер, несколько позднее – кожных покровов. Накапливаясь в коже и слизистой, билирубин в сочетании с другими пигментами прокрашивает их в светло-желтый цвет с красноватым оттенком. В дальнейшем происходит окисление билирубина в биливердин, и желтуха приобретает зеленоватый оттенок. При длительном существовании желтухи кожные покровы приобретают черновато-бронзовую окраску. Таким образом, осмотр больного позволяет решить вопрос о длительности желтухи, что имеет большое дифференциально-диагностическое значение.
Для того, чтобы классификацироать различные формы желтухи, необходимо знать основные данные физиологии билирубина.
Эритроциты разрушаются в селезенке или ретикуло-эндотелиальной системе. При этом гемоглобин расщепляется на глобин, железосодержащий гемосидерин и не содержащий железа гематоидин. Глобин распадается на аминокислоты и снова идет на построение белков организма. Железо подвергается окислению и снова использется организмом в виде ферритина. Гематоидин (порфириновое кольцо) превращается через стадию биливердина в билирубин. Свободный билирубин захватывается плазмой крови. Он совершенно не растворим в воде и соединяется с белками плазмы. В этом состоянии он задерживается печенью, где под влиянием фермента глюкоруноилтрансферазы переходит в билирубин - глюкуроновую кислоту. В отличие от свободного билирубина эта кислота растворима в воде. Таким образом в печени образуется прямой билирубин, который растворим в воде и в дальнейшем выделяется с желчью в кишечник. В кишечнике часть прямого билирубина превращается в уробилиноген, который, реабсорбируясь, частично возвращается в печень, а частично выделяется с мочой в виде уробилина (около 4 мг в сутки). Другая асть прямого билирубина, попавшего в кишечник, под действием кишечной флоры, превращается в стеркобилин, который выделяется с калом (60-80 мг в сутки).
Повышение в крови уровня непрямого билирубина может быть следствием повышенного гемолиза, при снижении способности гепатоцитов захватывать билирубин из плазмы крови, обусловленного недостаточностью трансферазной активности клеток печени (синдромы Жильбера и Криглера - Найара - наследственная недостаточность глюкуронилтрансферазы). Повышение в крови уровня прямого билирубина, как правило, обусловлено нарушением оттока желчи по внепеченочным желчным протокам (подпеченочная или механическая желтуха) или холангиолам (внутрипечечный холестаз).
Различают три основных вида желтух:
1). Рассмотрим механизмы развития гемолитической желтухи. Для этого необходимо кратко вспомнить определение гемолиза и его механизмы. Гемолизом называется усиленное разрушение эритроцитов. В норме эритроциты циркулируют в крови в течение 120 дней. При гемолитических состояниях продолжительность жизни эритроцитов значительно укорачивается, иногда до нескольких часов. Гемолиз бывает внутрисосудистым, когда эритроцитоц лизируются в крови, и внесосудистым - эритроциты подвергаются деструкции и перевариваются системой макрофагов. Внутрисосудистый гемолиз встречается относительно редко. Он может возникнуть вследствие механического разрушения эритроцитов при травмировании в малых кровеносных сосудах (маршевая гемоглобинурия), турбулентном кровяном потоке (дисфункция протезированных клапанов сердца), при прохождении через депозиты фибрина в артериолах (тромботическая тромбоцитопеническая пурпура, гемолитико-уремический синдром, ДВС-синдром), в результате несовместимости эритроцитов по системе АВ0, резус или какой-либо другой системе (острая посттрансфузионная реакция), под влиянием комплемента (пароксизмальная ночная гемоглобинурия, пароксизмальная холодовая гемоглобинурия), вследствие прямого токсического воздействия (яд змеи кобры, инфекции), при тепловом ударе.
Внесосудистый гемолиз осуществляется преимуещственно в селезенке и печени. Селезенка способна захватывать и разрушать малоизмененные эритроциты, печень различает лишь эритроциты с большими нарушениями, однако поскольку ток крови к печени примерно в 7 раз больше, чем к селезенке, печень может играть существенную роль в гемолизе.
Удаление эритроцитов из крови макрофагами происходит в основном двумя путями. Первый механизм связан с распознаванием макрофагами эритроцитов, покрытых IgG-АТ и С3, к которым на поверхности макрофагов имеются специфические рецепторы - иммуногемолитические анемии. Фагоцитоз эритроцитов сопровождается усиленной клеточной пролиферацией в селезенке (и в меньшей степени печени) с гиперплазией органа, увеличением тока крови в нем, повышением его функционально активности.
Второй механизм обусловлен изменениями физических свойств эритроцитов, снижением их пластичности - способности изменять свою форму при прохождении через узкую фильтрационную сеть селезенки. Нарушение пластичности отмечается при мембранных дефектах эритроцитов (наследственный микросфероцитоз), изменении липидного состава мембран (при циррозе печени), аномалиях гемоглобина, ведущих к повышению вязкости внутриклеточной среды эритроцита (серповидно-клеточная анемия), наличии включений в цитоплазме эритроцитов (бета-талассемия, альфа-талассемия), прециптированного гемоглобина в виде телец Гейнца (дефицит глюкозо-6-фосфатдегидрогеназы). Накопление эритроцитов в селезеночной пульпе, независимо от его мхеанизма, вызывает дальнейшую редукцию эритроцитарной поверхности – процесс "кондиционирования", что еще больше ухудшает условия продвижения клеток через синусы селезенки, приводит к необратимой задержке и деструкции эритроцитов путем их фрагментации и фагоцитоза макрофагами.
Наиболее важным признаком в начальной диагностике гемолиза является увеличение числа ретикулоцитов. Ретикулоцитоз отражает активацию костномозговой продукции эритроцитов в ответ на их разрушение на периферии. Кроме того, ускоренный выброс из костного мозга недозрелых эритроцитов, содержащих РНК, обуславливает феномен полихроматофилиии эритроцитов в фиксированных окрашенных азур-эозином мазках крови. Уровень непрямого билирубина в сыворотке крови при гемолизе повышен. Уровень его повышения зависит от активности гемолиза и способности гепатоцитов к образованию прямого билирубина и его экскреции. Уровень прямого билирубина (связанного) в сыворотке крови нормальный и билирубинурия отсутствует, так как через неповрежденный почечный фильтр проходит только прямой билирубин. В моче и кале содержится увеличенное количество уробилина и стеркобилина.
Таким образом, повышенный гемолиз независимо от его этиологии всегда приводит к характерной клинической триаде:
1. Анемия
2. Желтуха с лимонным оттенком
3. Спленомегалия
Несколько практических советов: как запомнить, что прямой билирубин – это связанный, а непрямой - свободный.
1. Быть прямым и свободным одновременно невозможно, следовательно, прямой билирубин связанный.
2. Не свой. Непрямой билирубин свободный.
2). При паренхиматозной желтухе повышается уровень прямого билирубина. В норме прямой билирубин выделяется в желчный капилляр и далее по билиарному дереву поступает в двенадцатиперстную кишку. Этот путь транспорта может нарушаться на внутриклеточном участке. Таким образом, при паренхиматозной желтухе страдает сама печеночная клетка. Этот вид желтухи возникает, например, при вирусных гепатитах. Вирус гепатита А обладает прямым повреждающим действием на клетки печени. Иммунная реакция в ответ на попадание вируса обычно адекватна, что и обусловливает редкость тяжелых форм заболевания и отсутствие перехода в хронический гепатит. Вирус гепатита В не обладает прямым цитопатогенным действием, но инкорпорирование вирусов или их частиц в мембраны печеночных клеток вызывает развитие клеточной цитотоксической реакции, направленной против клеточной оболочки, которая может приводить к некрозу печеночных клеток. Полный зрелый вирус гепатита В имеет 3 антигенные системы. В наружной оболочке вируса содержится поверхностный антиген (HbsAg), во внутренней оболочке - 2 антигена, HbеAg локализуется в цитоплазме гепатоцита, но не проникает в ядро, HbcAg (от латинского соr - сердцевина, ядро) проникает в ядро гепатоцита. Последний никогда не обнаруживается в крови, где можно выявить лишь антитела к нему. HbsAg способен образовывать с иммунные комплексы, которые могут циркулировать в крови (ЦИКи). Именно наличие циркулирующих иммунных комплексов при хроническом гепатите обусловливает наличие целого ряда системных проявлений. Кроме вирусных гепатитов, печеночные клетки поражаются и при гепатитах другой этиологии (острые токсические, лекарственные гепатиты, алкогольные гепатиты).
3).Обтурационная (механическая) желтуха. При этом виде желтухи также увеличивается уровень прямого билирубина, что обусловлено нарушением оттока желчи по внепеченочным желчным протока. Таким образом, обтурационная желтуха является следствием закупорки желчных протоков. Это патологическое состояние, которое не имеет однородных этиологических факторов. В патогенезе ключевым моментов является нарушение оттока желчи. Желчь не втекает в желудочно-кишечный тракт, следовательно не происходит нормального всасывания и переваривания.
Обтурация происходит за счет:
- камня
- опухоли
- рубцовой стриктуры протоков
При опухоли наблюдается ахолия, холемия. При закупорке камней присоединяется инфекция (холангит), что отягощает течение обтурационной желтухи. Наблюдается синдром Шарко или, другое название триада Шарко: желтуха, сопровождающаяся увеличением печени, боли, лихорадка. Это острое состояние, связанное с закупоркой. Застой желчи, закупорка и инфекция являются ведущими моментами. В тяжелых случаях может развиться сепсис, при котором появится пентада Рейнольда: триада Шарко + гипотония и спутанное сознание. Таким образом, речь идет о билиарном септическом шоке. Чаще всего камни вентильные. В крови лейкоцитоз, ускорение СОЭ, сдвиг формулы влево, анемия. В моче - желчные пигменты. Кал не окрашен, но при вентильном камне кал то окрашен, то нет. В биохимическом анализе крови щелочная фосфатаза не увеличена. Паренхиматозных поражений нет. Будут гнойная инфекция и синдром холестаза.
При закупорке опухолью клиника будет скудная, отмечается симптом Курвуазье - увеличенный, безболезненный желчный пузырь. Этот симптом связан с закупоркой дистальных отделов желчных путей. Этот симптом встречается при раке головки поджелудочной железы. Однако, если опухоль высокой локализации, то симптома Курвуазье не будет. Высокая локализация: рак желчного пузыря, пузырного протока. При высокой локализации опухоли будет синдром блокады доли печени: длительная время обтурационная желтуха, а билирубинемия не более 170 (норма 17-20). Происходит закупорка одного из печеночных протоков (правого или левого). Если рак головки поджелудочной железы, то билирубинемия достигает 300-400-500 Ед. В клиническом анализе крови при опухоли может быть незначительное увеличение СОЭ. Повышен прямой билирубин. Щелочная фосфатаза и пировиноградная фосфатаза не изменены.
Вопрос 134. С какими свойствами белков плазмы крови связана их способность регулировать объем крови и ее онкотическое давление? Какая белковая фракция играет наиболее существенную роль в обеспечении этой функции?
Ответ. Из 9–10% сухого остатка плазмы крови на долю белков приходится 6,5–8,5%. Используя метод высаливания нейтральными солями, белки плазмы крови можно разделить на три группы: альбумины, глобулины и фибриноген. Нормальное содержание альбуминов в плазме крови составляет 40–50 г/л, глобулинов – 20–30 г/л, фибриногена – 2,4 г/л. Плазма крови, лишенная фибриногена, называется сывороткой.
Синтез белков плазмы крови осуществляется преимущественно в клетках печени и ретикулоэндотелиальной системы. Физиологическая роль белков плазмы крови многогранна.
1. Белки поддерживают коллоидно-осмотическое (онкотическое) давление и тем самым постоянный объем крови. Содержание белков в плазме значительно выше, чем в тканевой жидкости. Белки, являясь коллоидами, связывают воду и задерживают ее, не позволяя выходить из кровяного русла. Несмотря на то что онкотическое давление составляет лишь небольшую часть (около 0,5%) от общего осмотического давления, именно оно обусловливает преобладание осмотического давления крови над осмотическим давлением тканевой жидкости. Известно, что в артериальной части капилляров в результате гидростатического давления безбелковая жидкость крови проникает в тканевое пространство. Это происходит до определенного момента – «поворотного», когда падающее гидростатическое давление становится равным коллоидно-осмотическому. После «поворотного» момента в венозной части капилляров происходит обратный ток жидкости из ткани, так как гидростатическое давление стало меньше, чем коллоидно-осмотическое. При иных условиях в результате гидростатического давления в кровеносной системе вода просачивалась бы в ткани, что вызвало бы отек различных органов и подкожной клетчатки.
2. Белки плазмы принимают активное участие в свертывании крови. Ряд белков, в том числе фибриноген, являются основными компонентами системы свертывания крови.
3. Белки плазмы в известной мере определяют вязкость крови, которая, как отмечалось, в 4–5 раз выше вязкости воды и играет важную роль в поддержании гемодинамических отношений в кровеносной системе.
4. Белки плазмы принимают участие в поддержании постоянного рН крови, так как составляют одну из важнейших буферных систем крови.
5. Важна также транспортная функция белков плазмы крови: соединяясь с рядом веществ (холестерин, билирубин и др.), а также с лекарственными средствами (пенициллин, салицилаты и др.), они переносят их к тканям.
6. Белки плазмы играют важную роль в процессах иммунитета (особенно это касается иммуноглобулинов).
7. В результате образования с белками плазмы недиализируемых комплексов поддерживается уровень катионов в крови. Например, 40–50% кальция сыворотки связано с белками, значительная часть железа, магния, меди и других элементов также связана с белками сыворотки.
8. Наконец, белки плазмы крови могут служить резервом аминокислот. Современные физико-химические методы позволили открыть и описать около 100 различных белковых компонентов плазмы крови. Особое значение приобрело электрофоретическое разделение белков плазмы (сыворотки) крови.
В сыворотке крови здорового человека при электрофорезе на бумаге можно обнаружить 5 фракций: альбумины, α1-, α2-, β-, γ-глобулины. Методом электрофореза в агаровом геле в сыворотке крови выделяют 7– 8 фракций, а при электрофорезе в крахмальном или полиакриламидном геле – до 16–17 фракций. Следует помнить, что терминология белковых фракций, получаемых при различных видах электрофореза, еще окончательно не установилась. При изменении условий электрофореза, а также при электрофорезе в различных средах (например, в крахмальном или полиак-риламидном геле) скорость миграции и, следовательно, порядок белковых зон могут меняться.
Еще большее число белковых фракций (свыше 30) можно получить методом иммуноэлектрофореза (рис. 17.1). Этот метод представляет собой своеобразную комбинацию электрофоретического и иммунологического методов анализа белков. Иными словами, термин «иммуноэлектрофорез» подразумевает проведение электрофореза и реакции преципитации в одной среде, т.е. непосредственно на гелевом блоке. При данном методе с помощью серологической реакции преципитации достигается значительное повышение аналитической чувстительности электрофоретического метода.
Вопрос 143. Приведите схему строения антител. Обозначьте на схеме участки взаимодействия с антигеном
Ответ. Основной функцией иммунной системы является защита генетической целостности организма от проникновения чужеродных веществ. Эта защита обеспечивается сложной системой органов, клеток и растворимых факторов. В механизмах устойчивости организма к генетически чужеродной информации участвуют два основных феномена: неспецифическая резистентность и приобретенный иммунитет.
Приобретенный антиинфекционный иммунитет отражает специфическую устойчивость, возникающую в организме в течение его жизни против конкретных видов микроорганизмов.
При этом приобретенный антиинфекционный иммунитет не обособлен от неспецифической резистентности организма, которую обеспечивают системы фагоцитов, комплемента, естественных киллеров, лизоцима, интерферонов и других медиаторов взаимодействия клеток, вызванных неспецифическими раздражителями; белков острой фазы воспаления и других веществ, участвующих в механизмах развития воспаления.
В развитии противовирусного иммунитета участвуют гуморальные и клеточные факторы. Особенности противовирусного иммунитета обусловлены своеобразием строения и биологии вирусов. Иммунитет направлен на нейтрализацию и удаление из организма вируса, его антигенов и зараженных вирусом клеток. Приобретенный противовирусный иммунитет, как и другие виды антиинфекционного иммунитета начинает развиваться со стадии представления антигена Т-хелперам. Напряженность противовирусного иммунитета зависит от уровня циркулирующих антител и образования цитотоксических лимфоцитов. Цитотоксические лимфоциты вызывают лизис инфицированных вирусом клеток.
Антитела, образующиеся при вирусных инфекциях, действуют непосредственно на вирус или на клетки, инфицированные вирусом. В связи с этим можно выделить две основные формы участия антител в развитии противовирусного иммунитета. Одна из них - нейтрализация вируса антителами. Такая нейтрализация препятствует рецепции вируса на клетке и проникновению его в клетку. Вторая форма участия антител - иммунный лизис инфицированных клеток.
Основную массу противовирусных антител составляют иммуноглобулины класса G. Антитела класса M свидетельствуют о свежеперенесенной инфекции, они появляются раньше и раньше исчезают по сравнению с IgG-антителами.
Схематическое строение антитела.
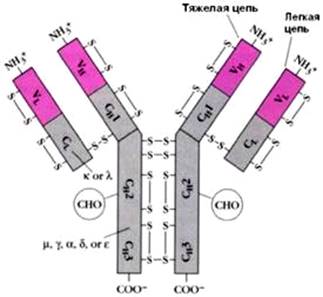
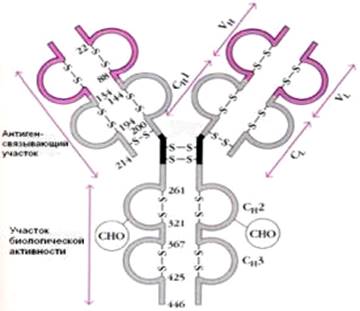
Прочность иммунитета при различных вирусных инфекциях значительно варьирует. При некоторых инфекциях (ветряная оспа, корь, паротит, краснуха) иммунитет достаточно стойкий. Повторные заболевания в этом случае редки. Менее стойкий иммунитет развивается при инфекциях дыхательных путей и кишечного тракта. Например, при гриппе иммунитет сохраняется в течение нескольких месяцев. Повторное заболевание гриппом объясняется прежде всего тем, что происходит постоянный дрейф поверхностных антигенных вирусных белков и смена циркулирующих штаммов.
Список используемой литературы
1. Березов Т.Т., Коровкин Б.Ф. «Биологическая химия». М., 2003
2. Комов Б.В. «Биохимия. Учебник для ВУЗов». М., 2004
3. Филиппович Ю.Б. «Биохимия». М., 2003
Похожие рефераты:
Влияние глауконита на продуктивные качества гусят-бройлеров
Шпоры по анатомии и физиологии человека