| Скачать .docx | Скачать .pdf |
Реферат: Строение и функции клеток
СОДЕРЖАНИЕ
1. ЖИВАЯ КЛЕТКА……………………………………………..3
2. ОСНОВНЫЕ ОСОБЕННОСТИ КЛЕТКИ…………………6
3. ОБМЕН МАТЕРИАЛАМИ МЕЖДУ КЛЕТКОЙ И ОКРУЖАЮЩЕЙ СРЕДОЙ……………………………………8
4. КЛЕТОЧНОЕ ЯДРО……………………………………………15
5. ЦЕНТРИОЛИ И МИТОТИЧЕСКОЕ ВЕРЕТЕНО. ……………19
6. МИТОХОНДРИИ…………………………………………………21
7. ХЛОРОПЛАСТЫ…………………………………………………24
8. РИБОСОМЫ И ДРУГИЕ ОРГАНЕЛЛЫ ЦИТОПЛАЗМЫ…25
9. СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ…………....29
ЖИВАЯ КЛЕТКА.
Поскольку биологию обычно определяют как «науку о живых организмах», мы должны прежде всего уметь дифференцировать «живое» и «неживое». Мы называем организмом любой живой объект, будь то растение, животное или бактерия. Сравнительно легко видеть, что человек, дуб, розовый куст, лев или дождевой червь — живые, а скалы и камни — неживые. Но считать ли живыми такие образования, как вирусы, — это уже зависит от того, как мы определим понятие «жизнь».
Почти все организмы построены из обособленных единиц, называемых клетками. Каждая клетка представляет собой самостоятельную функциональную единицу, а протекающие в организме процессы слагаются из совокупности координированных функций его клеток. Клетки могут быть весьма различны по своим размерам, форме и функции. У некоторых мельчайших организмов все тело состоит из одной клетки. Другие организмы, например человек или дуб, построены из многих миллиардов клеток, пригнанных друг к другу.
В 1839 году чешский физиолог Пуркинье ввел для обозначения живого содержимого клетки термин протоплазма. Когда исследователи лучше изучили структуру и функции клеток, стало ясно, что живое содержимое клетки представляет собой неимоверно сложную систему разнородных компонентов (рис. 1). Термин «протоплазма» не имеет четкого физического или химического значения, но им до сих пор можно пользоваться для обозначения всех организованных компонентов клетки.
Чтобы получить представление о том, как выглядит протоплазма, мы можем исследовать какой-нибудь просто устроенный организм, вроде амебы или миксомицета (слизистого гриба), у которого этот живой материал ничем не прикрыт и поэтому хорошо виден под микроскопом. Протоплазма подобного организма полупрозрачна, причем она либо бесцветна, либо имеет слегка желтоватую, красноватую или зеленоватую окраску. Она обладает вязкой консистенцией густого сиропа и показалась бы слизистой на ощупь. При помощи обычного микроскопа в ней можно иногда различить зернышки или волоконца из более плотного материала, капельки жировых веществ или наполненные жидкостью пузырьки (вакуоли); все это взвешено в прозрачном однородном полужидком «основном веществе». Однако в том материале, который при исследовании в обычном микроскопе казался более или менее гомогенным, электронный микроскоп открывает удивительно сложные структуры (рис. 1, В и Г). Как показал рентгеноструктурный анализ, клеточные мембраны и разнообразные внутриклеточные образования обладают еще более тонкой структурой, определяемой, по-видимому, структурой больших молекул, из которых они состоят.
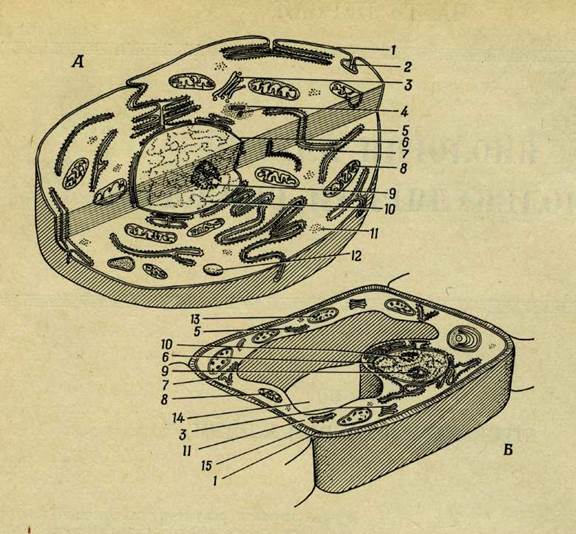
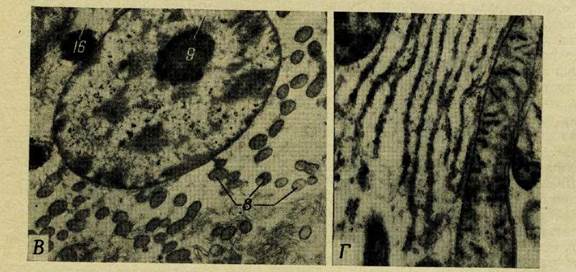
Рис. 1. Строение клетки.
А. Схема типичной животной клетки. Б. Схема типичной растительной клетки. В. Электронная микрофотография ядра и окружающей его цитоплазмы в клетке печени лягушки (X 16 500). Г. Электронная микрофотография митохондрий и микросом в клетке печени крысы (X 65 000); на микросомах видны зернышки нуклеопротеида, а в верхнем левом углу и справа в митохондриях можно видеть структуры с двойными мембранами. 1 — клеточная мембрана; 2 — пиноцитозный пузырек; 3 —тельце Гольджи; 4 — центриоли; 5 — рибосома; 6 — ядерная мембрана; 7 — эндоплазматическая сеть; 8— митохондрии; 9 — ядрышко; 10 — ядро; 11 — цитоплазма; 12 — лизосома; 13 — хлоропласт; 14 — вакуоль; 15 — клеточная стенка; 16 — липидное включение.
ОСНОВНЫЕ ОСОБЕННОСТИ КЛЕТКИ.
Каждая клетка содержит ядро и окружена плазматической мембраной. Эритроциты млекопитающих и клетки ситовидных трубок флоэмы в процессе своего созревания теряют ядро, а в поперечнополосатых мышцах и у многих грибов и водорослей на каждую клетку приходится по нескольку ядер, У простейших растений и животных весь живой материал заключен в одну плазматическую мембрану. Такие организмы можно считать одноклеточными или бесклеточными (т. е. имеющими тело, не разделенное на клетки). Однако их единственная клетка может быть высокоспециализирована как морфологически, так и функционально и может иметь очень большие размеры — крупнее, чем все тело некоторых многоклеточных организмов. Поэтому неверно было бы думать, что одноклеточный организм непременно должен быть мельче и проще многоклеточного.
У разных растений и животных и в различных органах одного и того же растения или животного клетки поразительно разнообразны по своим размерам, форме, окраске и внутреннему строению. Однако все они имеют ряд общих особенностей: каждая клетка окружена плазматической мембраной, имеет ядро и содержит различного рода внутриклеточные органеллы. К последним относятся митохондрии, шероховатая (гранулярная) и гладкая (агранулярная) эндоплазматическая сеть, комплекс Гольджи, лизосомы и центриоли.
Все организмы и составляющие их клетки имеют более или менее определенные размеры и форму. В них происходят метаболические реакции. Они обладают раздражимостью, способны к движению, росту, размножению и приспособлению к изменениям внешней среды. Хотя этот перечень свойств кажется вполне четким и определенным, граница между живым и и неживым довольно условна. Вирусам, например, свойственны лишь некоторые, но не все черты, характерные для живых организмов. Если мы поймем, что мы не в состоянии обоснованно ответить на вопрос, являются ли вирусы живыми, а можем лишь решать, следует ли называть их живыми, то проблема эта предстанет перед нами в правильном ракурсе. Неживые объекты могут обладать одним или несколькими из перечисленных выше свойств, но не всеми одновременно. Кристаллы в насыщенном растворе могут «расти», кусочек металлического натрия начинает быстро «бегать» по поверхности воды, а капля масла, плавающая в смеси глицерина и спирта, выпускает псевдоподии и передвигается наподобие амебы.
Тот или иной род живых организмов всегда можно распознать по характерным для него форме и внешнему виду; взрослые особи каждого рода организмов, как правило, имеют определенные размеры. В отличие от этого размеры и форма неживых объектов гораздо менее постоянны. Живые организмы не гомогенны, а состоят из различных частей, выполняющих те или иные специальные функции; таким образом, они характеризуются специфической сложной организацией. Структурной и функциональной единицей как у растений, так и у животных служит клетка, которая в свою очередь тоже имеет специфическую организацию; каждый вид клеток обладает характерными размерами и формой, по которым его можно распознать.
Совокупность осуществляемых клеткой биохимических процессов, обеспечивающих ее рост, поддержание и восстановление, называется обменом веществ, или метаболизмом. Протоплазма каждой клетки непрерывно изменяется: она поглощает новые вещества, подвергает их разнообразным химическим изменениям, строит новую протоплазму и превращает в кинетическую энергию и тепло потенциальную энергию, заключенную в молекулах белков, жиров и углеводов, по мере того как эти вещества превращаются в другие, более простые соединения. Это постоянное расходование энергии представляет собой одну из характерных особенностей живых организмов, свойственных им одним. Некоторые типы клеток, например бактериальные клетки, отличаются высокой интенсивностью обмена. Другие клетки, например семена и споры, имеют столь низкий уровень обмена, что его с трудом удается обнаружить даже с помощью самых чувствительных приборов. Даже в пределах одного вида организмов или у одной особи интенсивность обмена может меняться в зависимости от таких факторов, как возраст, пол, общее состояние организма, активность эндокринных желез, беременность.
Процессы обмена веществ принято разделять на анаболические и катаболические. Анаболиз мом называют те химические процессы, при которых более простые вещества соединяются между собой с образованием более сложных веществ, что приводит к накоплению энергии, построению новой протоплазмы и росту. Катаболиз мом же называют расщепление этих сложных веществ, приводящее к освобождению энергии; при этом происходит разрушение протоплазмы и расходование составляющих ее веществ. Процессы того и другого типа протекают непрерывно, и взаимозависимость между ними столь велика, что их трудно разграничить. Сложные соединения расщепляются, и их составные части соединяются друг с другом в новых комбинациях, образуя другие сложные вещества. Примером сочетания катаболизма с анаболизмом могут служить взаимные превращения углеводов, белков и жиров, непрерывно происходящие в клетках нашего тела. Поскольку большинство анаболических процессов требует затраты энергии, необходимы какие-то катаболические процессы, которые поставляли бы энергию для реакций, связанных с построением новых молекул. Клетки зеленых растений обладают способностью синтезировать свои собственные органические соединения из минеральных веществ, которые они получают из почвы и воздуха; животные же зависят в своем питании от растений.
Другая особенность живых организмов — это их способность к движению. Подвижность большинства животных совершенно очевидна: они ползают, плавают, бегают или летают. Движения растений гораздо более медленны и не так заметны, но они все же происходят. Некоторые животные — губки, кораллы, устрицы, многие паразиты — сами не передвигаются с места на место, но у большинства из них имеются реснички или жгутики, приводящие в движение окружающую жидкую среду, которая доставляет этим животным пищу и все необходимое для жизни. Движение может быть результатом мышечного сокращения, биения ресничек или жгутиков (рис. 2, А) и, наконец, медленного течения массы протоплазмы — амебоидное движение (рис. 2, Б). Течение протоплазмы в клетках листьев растений известно под названием циклоза (рис. 2, В).
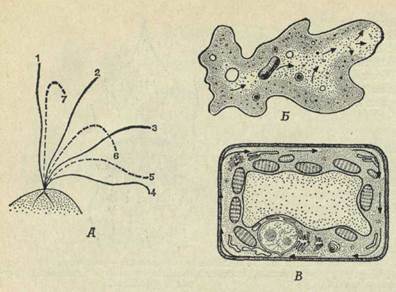 Рис. 2. Схема, иллюстрирующая различные типы клеточных движений.
Рис. 2. Схема, иллюстрирующая различные типы клеточных движений.
А. Движение жгутика. Б. Амебоидное движение. В. Циклоз.
Живые организмы обладают раздражимос тью: они реагируют на раздражители (стимулы), т. е. на физические или химические изменения в непосредственно окружающей их среде. Раздражители, вызывающие реакцию у большинства животных и растений,— это изменения цвета, интенсивности или направления световых лучей; изменения температуры, давления или звука; изменения в химическом составе почвы, воды или атмосферы, окружающей организм. Некоторые высокоспециализированные клетки тела обладают особой чувствительностью к раздражителям определенного типа: палочки и колбочки в сетчатке глаза реагируют на свет, определенные клетки в носу и во вкусовых почках языка — на химические стимулы, а специальные клетки кожи — на изменения температуры или давления. У низших животных и у растений такие специализированные клетки могут отсутствовать, но организм в целом реагирует на раздражение. Одноклеточные животные и растения отвечают на воздействие тепла или холода, некоторых веществ, света или на прикосновение микроиглы движением по направлению к раздражителю или от него.
Раздражимость растительных клеток не всегда столь очевидна, как раздражимость животных клеток, но и растительные клетки чувствительны к изменениям окружающей среды. Так, при изменении освещения течение протоплазмы в клетках растений иногда ускоряется или прекращается. Некоторые растения (например, венерина мухоловка, растущая на болотах) чрезвычайно чувствительны к прикосновению и благодаря этому могут ловить насекомых: листья таких растений способны перегибаться вдоль средней жилки, а края их снабжены волосками; в ответ на раздражение, производимое насекомым, лист складывается, его края сближаются, а волоски, переплетаясь, не дают добыче выскользнуть; затем лист выделяет жидкость, которая убивает и переваривает насекомое. Способность ловить насекомых развилась как приспособление, позволяющее таким растениям получать часть необходимого для их роста азота из «поедаемой» добычи.
Рост живой ткани, т. е. увеличение клеточной массы, может происходить за счет увеличения размеров отдельных клеток, за счет увеличения их числа или же за счет того и другого. Увеличение размеров клеток может быть вызвано просто поглощением воды, но такого рода набухание обычно не рассматривают как рост. Ростом принято называть лишь те процессы, при которых увеличивается количество живого вещества организма, измеряемое количеством азота или белка. (Как вы думаете, почему показателем роста служит количество азота или белка, а не количество углеводов, жиров, серы или натрия?) Рост различных частей организма может происходить либо равномерно, либо одни части растут быстрее других, так что пропорции тела во время роста изменяются. Некоторые организмы могут расти в течение неопределенно долгого времени, тогда как другие имеют ограниченный период роста, заканчивающийся после достижения определенных размеров. Одна из замечательных особенностей процесса роста состоит в том, что всякий растущий орган, так же как и любая растущая клетка, продолжает в то же время функционировать.
Если есть какое-либо свойство, которое можно считать совершенно обязательным атрибутом жизни, так это способность к воспроизведению. Наиболее простые вирусы лишены обмена веществ, не двигаются и не растут, и все-таки, поскольку они способны воспроизводить себя, а также мутировать, большинство биологов считает их живыми. Так как все живое происходит только от живого и не может возникать путем самозарождения, эта способность воспроизводить самих себя является важнейшей особенностью живых организмов.
Процесс размножения может сводиться к простому делению одного индивидуума на два. Однако у большей части организмов размножение связано с образованием специализированных клеток — яйцеклеток и сперматозоидов, которые, соединяясь между собой, образуют оплодотворенное яйцо, или зиготу, развивающуюся в новый организм. У некоторых паразитических червей процесс развития слагается из нескольких совершенно различных форм, сменяющих одна другую, пока цикл не завершится образованием взрослой особи.
Способность растения или животного приспосабливаться (адаптироваться)
к окружающим условиям позволяет ему выжить в мире, полном неожиданных перемен. Тот или иной вид может либо отыскивать пригодную для его жизни среду, либо претерпевать изменения, делающие его лучше приспособленным к существующим в данный момент внешним условиям. Адаптация может осуществляться путем немедленного изменения, основанного на раздражимости, или путем длительного процесса мутирования и отбора. Конечно, отдельное растение или животное не может приспособиться ко![]() всем возможным условиям среды, а это означает, что существуют определенные области, где оно не сможет выжить. Перечень факторов, которые могут ограничивать распространение вида, почти бесконечен: вода, свет, температура, пища, хищники, конкуренты, паразиты и т. п.
всем возможным условиям среды, а это означает, что существуют определенные области, где оно не сможет выжить. Перечень факторов, которые могут ограничивать распространение вида, почти бесконечен: вода, свет, температура, пища, хищники, конкуренты, паразиты и т. п.
ОБМЕН МАТЕРИАЛАМИ МЕЖДУ КЛЕТКОЙ И ОКРУЖАЮЩЕЙ СРЕДОЙ
Снаружи каждая клетка одета нежным эластичным покровом, который составляет неотъемлемый функциональный компонент клетки и называется плазматической мембра ной. Эта мембрана играет чрезвычайно важную роль в регулировании состава клеточного содержимого, так как через нее в клетку поступают все питательные вещества и выходят наружу все отходы или продукты секреции. Мембрана задерживает проникновение в клетку одних веществ и облегчает поступление других. Клетки почти всегда окружены водной средой; это может быть пресная или морская вода (в случае простейших организмов), тканевый сок (высшие растения), плазма или внеклеточная жидкость (высшие животные).
Плазматическая мембрана действует так, как будто она пронизана ультрамикроскопическими порами, через которые проходят определенные вещества, причем величиной этих пор определяется максимальная величина способных пройти через них молекул. Возможность прохождения вещества через мембрану зависит не только от величины молекул, но и от электрического заряда диффундирующей частицы (если она им обладает), от присутствия и числа молекул воды, связанных с поверхностью этих частиц, и от растворимости частиц в липидах. Химическая и физическая природа мембраны выяснена еще не до конца, но, по-видимому, это трехслойная пленка толщиной около 12 нм. Наружный и внутренний слои, каждый толщиной около 3 нм, состоят из белка, а между ними лежит слой фосфолипидных молекул, имеющий толщину 60 нм.
Такого рода трехслойную структуру можно видеть на электронных микрофотографиях, полученных при высоком разрешении. Интересно, что все плазматические мембраны животных, растительных и бактериальных клеток, так же как и мембраны разнообразных внутриклеточных органелл, имеют, видимо, сходное трехслойное строение. Два слоя белка, разделенные слоем липида, так называемая элементарная мембрана, по-видимому, представляет собой широко распространенную основную единицу мембранной структуры.
У растений почти все клетки имеют, кроме того, толстую клеточную стенку, состоящую из целлюлозы и лежащую кнаружи от плазматической мембраны (у большинства животных клеток ее нет). Клеточная стенка во многих местах пронизана мельчайшими отверстиями, через которые протоплазма одной клетки соединяется с протоплазмой других, соседних с ней клеток; через эти отверстия вещества могут переходить из одной клетки в другую. Плотные, прочные клеточные стенки создают опору телу растения.
Для того чтобы понять механизмы, лежащие в основе обмена материалами между клеткой и окружающей средой, мы должны прежде всего учитывать, что для всех молекул в жидкостях и газах характерна тенденция диффундировать, т. е. перемещаться во всех направлениях до тех пор, пока они не распределятся равномерно по всему доступному пространству. Диффузию можно определить как распространение молекул из области их высокой концентрации в область более низкой концентрации, обусловленное их тепловым движением (рис. 19). Скорость диффузии зависит от величины молекул и от температуры. Молекулы, из которых построены все вещества, в том числе и твердые, находятся в постоянном движении. Основное различие между тремя состояниями вещества — твердым, жидким и газообразным — определяется степенью свободы движения молекул. Молекулы твердого тела упакованы сравнительно плотно, и силы притяжения между ними позволяют им совершать колебания, но не допускают свободного передвижения. В жидкости расстояния между молекулами несколько больше, межмолекулярные силы слабее и молекулы обладают значительной свободой передвижения. Наконец, в газообразном веществе молекулы настолько удалены друг от друга, что межмолекулярные силы ничтожны и свободу движения молекул ограничивают только внешние препятствия.
Если исследовать каплю воды под микроскопом, то движение молекул обнаружить не удастся, но если добавить к ней капельку туши (которая содержит мелкие частицы угля), то можно наблюдать непрерывное беспорядочное движение частиц угля. Каждая частица угля непрерывно подвергается ударам молекул воды, и толчки от этих ударов приводят эти частицы в движение. Такое движение мелких частиц называют броуновским движением по имени Роберта
Броуна — английского ботаника, который его впервые заметил, рассматривая под микроскопом пыльцевые зерна в капле воды. В процессе диффузии каждая молекула движется по прямой линии, пока не столкнется с чем-нибудь, например с другой молекулой или со стенкой сосуда; тогда она отскакивает и начинает перемещаться в другом направлении. Молекулы продолжают двигаться и после того, как они равномерно распределились по всему доступному пространству; однако, в то время как одни молекулы переходят, например, слева направо, другие переходят справа налево, и равновесие сохраняется. Различные вещества (сколько бы их ни было), находящиеся в одном и том же растворе, диффундируют независимо друг от друга. Скорость движения отдельных молекул может достигать нескольких сот метров в секунду, но каждая молекула до столкновения с другой, от которой она отскакивает, проходит лишь малую долю нанометра. Поэтому перемещение молекулы в каком-нибудь одном направлении происходит очень медленно. В этом можно убедиться, положив на дно стеклянного цилиндра, наполненного водой, кусочек какой-нибудь краски. Через несколько дней можно заметить, что красящее вещество постепенно поднимается вверх, но пройдут месяцы, прежде чем краска равномерно распределится по всему цилиндру. Таким образом, хотя диффузия на очень малые расстояния происходит весьма быстро, для того чтобы пройти расстояние в несколько сантиметров, молекулам требуется много времени.
Этот факт имеет большое биологическое значение, так как из него следует, что число молекул кислорода или питательных веществ, которые могут достигнуть организма путем одной только диффузии, весьма ограничено. Лишь очень небольшой организм, которому ежесекундно необходимо относительно небольшое число молекул питательных веществ или кислорода, может выжить, сидя на одном месте и дожидаясь, когда эти молекулы дойдут до него путем диффузии. Более крупный организм должен иметь возможность либо передвигаться с места на место, либо приводить в движение окружающую среду и таким путем доставлять себе нужные молекулы, либо, наконец, он может обитать в таком месте, где сама среда непрерывно движется, например в реке или на морском побережье в зоне приливов. Крупные наземные растения — деревья и кустарники — разрешили эту проблему особым образом: у них имеется чрезвычайно сильно разветвленная корневая система, при помощи которой они получают необходимые им вещества с большого участка окружающей среды.
Могут ли молекулы данного вещества проходить через ту или иную мембрану, зависит от ее структуры и от величины имеющихся в ней пор. Мембрану называют проницаемой, если через нее проходит любое вещество, непроницаемой — если она не пропускает ни одно вещество, избирательно проницаемой, или полу проницаемой, — если через нее могут диффундировать некоторые, но не все вещества. Все клеточные мембраны (окружающие саму клетку, ядра, вакуоли и различные субклеточные структуры) обладают дифференциальной проницаемостью.
Диффузия растворенного вещества через полупроницаемую мембрану называется диализом. Для того чтобы продемонстрировать процесс диализа, можно взять мешочек из коллодия, целлофана или пергамента, наполнить его раствором сахара и поместить в сосуд с водой. Если поры мембраны не слишком мелки, то молекулы сахара будут проходить сквозь нее. Со временем концентрация сахара в воде, окружающей мешочек, станет равна концентрации его в мешочке. Диффузия молекул будет продолжаться и после этого, но концентрация изменяться не будет, так как диффузия в обоих направлениях будет происходить с одинаковой скоростью.
Но если взять мешочек с более мелкими порами, с тем чтобы он был проницаем для молекул воды, но непроницаем для более крупных молекул сахара, то наблюдается другое явление. Нальем в мешочек раствор сахара, снабдим мешочек пробкой с проходящей через нее стеклянной трубкой и поместим его в сосуд с водой. Молекулы сахара не проходят через мембрану и поэтому остаются внутри мешочка. Молекулы воды, однако, диффундируют через мембрану в раствор сахара. Жидкость внутри мешочка содержит 5% сахара, а следовательно, лишь 95% воды. Жидкость же, окружающая мешочек снаружи, представляет собой чистую воду. Поэтому молекулы воды переходят из области более высокой концентрации (100% — снаружи) в область более низкой концентрации (95% — внутри мешочка). Такая диффузия молекул воды или другого растворителя через мембрану называется осмосом.
По мере того как происходит осмос, вода поднимается по стеклянной трубке. Если через мембраны пройдет такое количество воды, какое первоначально содержалось в мешочке, то раствор сахара окажется разбавленным до соотношения 2,5% сахара: 97,5% воды, но концентрация воды снаружи все еще будет выше, чем концентрация ее внутри, и осмос будет продолжаться. В конце концов уровень воды в стеклянной трубке поднимется настолько что давление, производимое водой в трубке, будет равно силе, заставляющей воду входить в мешочек. После этого изменение количества воды в мешочке прекратится; осмос через полупроницаемую мембрану будет продолжаться с одинаковой скоростью в обоих направлениях.
Давление столба воды в трубке служит мерой осмотического давления раствора сахара. Осмотическое давление вызывается стремлением молекул воды пройти сквозь полупроницаемую мембрану и выравнять концентрацию воды по обе стороны мембраны. Более концентрированный раствор сахара имел бы еще более высокое осмотическое давление и «насасывал» бы воду в трубку до более высокого уровня. При 10%-ной концентрации раствора сахара вода в трубке поднялась бы приблизительно вдвое выше, чем при 5%-ной.
Диализ и осмос — всего лишь две особые формы диффузии. Диффузия — общий термин для обозначения движения молекул из области высокой концентрации в область более низкой концентрации под действием тепловой энергии этих молекул. Диализом называется диффузия молекул растворенного вещества через полупроницаемую мембрану, а осмосом — диффузия через такую же мембрану молекул растворителя. В живых системах растворителем служит вода.
В жидком содержимом всякой живой клетки растворены соли, сахара и другие вещества, благодаря которым оно обладает известным осмотическим давлением. Если поместить клетку в жидкость с таким же осмотическим давлением, как и давление в клетке, вода не входит в клетку и не выходит из нее, а клетка соответственно не набухает и не съеживается; такую жидкость называют изотонической или изоосмоти ческой по отношению к внутриклеточной жидкости. В норме плазма крови и все жидкости организма изотоничны; они содержат такое же количество растворенного материала, как и клетки.
Если концентрация растворенных веществ в окружающей жидкости выше, чем внутри клетки, то вода стремится выйти наружу и клетка съеживается. Такую жидкость называют гипертонической по отношению к клетке. Если в жидкости меньше растворенного материала, чем в клетке, ее называют гипотониче ской; в этом случае вода стремится входить в клетку и вызывает ее набухание. 0,9%-ный раствор хлористого натрия (иногда называемый «физиологическим» раствором) изотоничен по отношению к клеткам человека. Эритроциты, помещенные в 0,6%-ный раствор хлористого натрия, набухают и лопаются (рис. 21), в 1,3%-ном растворе они сжимаются, а в 0,9%-ном растворе с ними не происходит ни того, ни другого.
Если клетку поместить в раствор, не изотонический по отношению к ее содержимому, то она может иногда приспособиться к такой среде, изменяя содержание воды в своей протоплазме (набухая или сжимаясь) до тех пор, пока концентрация растворенных веществ в клетке и в окружающей среде не станет одинаковой. Многие клетки способны активно насасывать воду или некоторые растворенные вещества через плазматическую мембрану и откачивать их наружу, в результате чего поддерживается осмотическое давление, отличное от осмотического давления окружающей среды. У простейших, живущих в сильно гипотонической пресной воде, выработались сократительные вакуоли, которые вбирают в себя воду из протоплазмы и выводят ее наружу. Перед растениями, обитающими в пресной воде, тоже встает проблема: как быть с водой, проникающей в клетки из окружающей гипотонической среды путем осмоса? Клетки растений не имеют сократительных вакуолей, чтобы «откачивать» воду, но зато прочная целлюлозная стенка клетки предохраняет ее от чрезмерного набухания. По мере поступления в клетку воды в ней возникает внутреннее давление, называемое тургором, которое препятствует дальнейшему проникновению воды. Тургор вообще характерен для растительных клеток; именно он «поддерживает» тело растения. Когда тургорное давление в клетках понижается вследствие недостатка воды, цветок увядает.
Многие организмы, обитающие в море, обладают феноменальной способностью избирательно накапливать некоторые вещества из морской воды. Морские водоросли могут накапливать йод в таком количестве, что концентрация его в клетках становится в 2 миллиона раз выше, чем в окружающей среде. Примитивные хордовые — оболочники — способны накапливать ванадий, и концентрация этого элемента в их клетках может превышать его концентрацию в морской воде тоже примерно в два миллиона раз. Перенос воды или растворенных веществ в клетку или из клетки против градиента концентрации представляет собой физическую работу и требует затраты энергии. Клетка способна перемещать молекулы против градиента концентрации лишь пока она жива и пока в ней происходят метаболические процессы, доставляющие энергию. Если на клетку воздействовать каким-нибудь метаболическим ядом («отравляющим» обмен веществ), например цианистым калием, она утрачивает способность создавать и поддерживать разность концентраций по обе стороны плазматической мембраны.
КЛЕТОЧНОЕ ЯДРО
Каждая клетка содержит небольшую, обычно шаровидную или овальную органеллу, называемую ядром. В одних клетках ядро занимает относительно постоянное положение и располагается примерно в центре клетки; в других оно свободно перемещается и может оказаться почти в любом участке. Ядро играет важную роль в регулировании протекающих в клетке процессов; оно содержит наследственные факторы, или гены, определяющие признаки данной клетки и всего организма, и прямо или косвенно регулирует многие стороны клеточной активности. Ядро отделено от окружающей цитоплазмы ядерной мембраной, состоящей из двух элементарных мембран; ядерная мембрана регулирует передвижение веществ из ядра в цитоплазму и обратно. В электронном микроскопе видно, что ядерная мембрана состоит из двух слоев и что в ней имеются поры (рис. 22), через которые содержимое ядра сообщается с цитоплазмой; возможно, что через эти поры проходят информационные макромолекулы. По-видимому, наружный из двух слоев ядерной мембраны без перерыва переходит в мембраны эндоплазматической сети и комплекса Гольджи.
В ядре клетки, убитой путем фиксации в соответствующих химических веществах и окрашенной подходящими красителями, выявляются различные структуры. В живой клетке их трудно увидеть с помощью обычного светового микроскопа, но они хорошо видны в фазово-контрастном микроскопе (рис. 3). В кариоплазме — полужидком основном веществе ядра — взвешено строго определенное число вытянутых нитевидных образований, называемых хромосомами; они состоят из ДНК и белка и содержат единицы наследственности — гены. На окрашенном срезе (рис. 3) клетки в состоянии покоя (в период между делениями) хромосомы обычно неразличимы, а вместо них
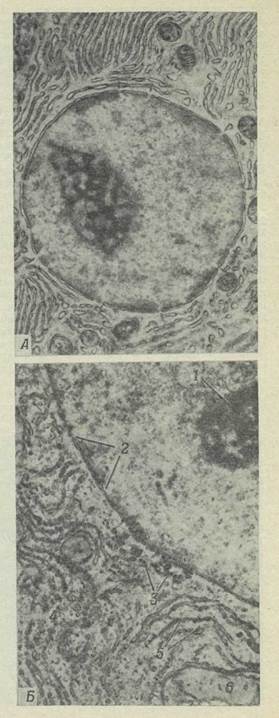 Рис. 22.
Рис. 22.
А. Электронная микрофотография ядра и окружающей его эндоплазматической сети; поры показаны стрелками (Х 20 000). Б. Часть того же препарата при большем увеличении (Х 50 000). 1 — ядрышко; 2 — поры; 3 — гликоген; 4 — рибосомы; 5 —эндоплазматическая сеть; е — митохондрия.
видна сеть из темных тяжей и зернышек, в совокупности называемых хроматином. Перед началом деления ядра эти тяжи уплотняются в компактные палочковидные хромосомы, которые впоследствии строго поровну распределяются между двумя дочерними клетками. Для каждого организма характерно строго определенное число хромосом, содержащееся в каждой из составляющих его клеток. У плодовой мушки (дрозофилы) 8 хромосом, у сорго их 10, у садового гороха 14, у кукурузы 20, у жабы 22, у томата 24, у вишни 32, у крысы 42, у человека 46, у картофеля 48, у козы 60, а у утки 80. Приведенные цифры относятся к соматическим клеткам высших растений и животных, в которых хромосома каждого типа представлена в двойном числе; клетку с двумя полными наборами хромосом называют диплоидной. Сперматозоиды и яйцеклетки, в которых каждая хромосома представлена лишь в единственном числе (т. е. имеется один полный набор хромосом), называют гаплоидными клетками. Число хромосом в них вдвое меньше, чем в соматических клетках того же организма. При оплодотворении яйцеклетки сперматозоидом два гаплоидных набора хромосом объединяются и таким образом восстанавливается их диплоидное число. В ядре имеется сферическое тельце, называемое ядрышком. В большинстве клеток ядрышко чрезвычайно изменчиво: оно меняет свою форму и структуру, появляется и исчезает. Ядро может содержать и несколько ядрышек, но обычно клетки каждого вида животных или растений содержат определенное число ядрышек. Ядрышки исчезают, когда клетка готовится к делению, а затем появляются вновь; они, по-видимому, участвуют в синтезе рибонуклеиновой кислоты, входящей в состав рибосом. Если ядрышко разрушить сфокусированным пучком рентгеновских или ультрафиолетовых лучей, то клеточное деление подавляется. Если же подвергнуть облучению другой участок ядра, не затрагивая ядрышко, то этого не происходит.
Для того чтобы выяснить роль ядра, можно удалить его из клетки и наблюдать последствия такой операции. Если с помощью микроиглы удалить ядро у одноклеточного животного — амебы, то клетка продолжает жить и двигаться, но не может расти и через несколько дней погибает. Следовательно, ядро необходимо для метаболических процессов (в первую очередь — для синтеза нуклеиновых кислот и белков), обеспечивающих рост и размножение клеток.
Можно возразить, что к гибели приводит не утрата ядра, а сама операция. Для того чтобы выяснить это, необходимо поставить опыт с контролем, т. е. подвергнуть две группы амеб одной и той же операции, с той разницей, что в одном случае ядро действительно удаляют, а в другом в амебу вводят микроиглу, передвигают ее в клетке подобно тому, как это делается при удалении ядра, и выводят, оставив ядро в клетке; это называется «мнимой» операцией. После такой процедуры амебы оправляются, растут и делятся; это показывает, что гибель амеб первой группы вызывалась не операцией как таковой, а именно удалением ядра.
Классическую серию опытов, доказывающих важную роль ядра в регуляции клеточного роста, провел Геммерлинг на одноклеточном (неклеточном) растении Acetahularia mediter -тапеа. Эта морская водоросль, которая может достигать высоты 5 см, внешне несколько напоминает гриб и имеет «корни» и «стебель», заканчивающийся вверху большой дисковидной «шляпкой». Все растение представляет собой одну-единственную клетку и содержит лишь одно ядро, находящееся у основания стебля. Геммерлинг установил, что если перерезать стебель, то нижняя часть остается живой, регенерирует шляпку и полностью оправляется после операции. Верхняя же часть, лишенная ядра, живет в течение некоторого времени, но в конце концов погибает, не будучи в состоянии восстановить нижнюю часть. Следовательно, ацетабулярии, так же как и амебе, ядро необходимо для метаболических процессов, лежащих в основе роста; регенерация — это, конечно, одна из форм роста. В последующих экспериментах Геммерлинг сначала перерезал стебель непосредственно над ядром, а затем вторично перерезал его чуть ниже шляпки. Изолированный отрезок стебля, будучи снова помещен в морскую воду, частично или полностью восстанавливал шляпку. Казалось бы, это говорит о том, что ядро не является необходимым для регенерации; однако если Геммерлинг удалял и вторую шляпку, то третья шляпка уже не развивалась.
На основании таких экспериментов Геммерлинг заключил, что ядро вырабатывает какое-то вещество, необходимое для образования шляпки. Это вещество распространяется путем диффузии вверх по стеблю и стимулирует рост шляпки. В только что описанных опытах после перерезок 1 и 2 в стебле оставалось достаточно этого вещества, чтобы вызвать образование еще одной шляпки. Однако после того, как это вещество было израсходовано на образование одной новой шляпки, регенерация второй шляпки при отсутствии ядра оказалась уже невозможной.
У другого вида, Acetabularia crenulata , шляпка ветвистая, а не дисковидная. Если кусочек стебля этого вида (без ядра) пересадить на нижнюю часть стебля A . mediterranea (с ядром A . mediterranea ), то на верхушке стебля образуется новая шляпка, но форма ее определяется не пересаженным кусочком стебля, а нижней частью, на которую он пересажен. Ядро благодаря содержащимся в нем генам доставляет специфическую информацию, определяющую форму регенерирующей шляпки, и его влияние оказывается сильнее тенденции пересаженного кусочка стебля образовать шляпку, характерную для вида A . crenulata . Ядро может контролировать активность других частей клетки благодаря тому, что в его хромосомах закодированы «инструкции», необходимые для синтеза белков и других веществ, от которых зависят структурные особенности и функции клетки. Очевидно, всякий раз, когда клетка делится, весь этот набор инструкций удваивается и каждая из дочерних клеток получает по одному его экземпляру.
ЦЕНТРИОЛИ И МИТОТИЧЕСКОЕ ВЕРЕТЕНО.
В клетках животных и некоторых низших растений около ядра находятся два небольших интенсивно окрашивающихся тельца — центриоли. Центриоли играют важную роль в клеточном делении: в начале деления они отходят друг от друга, направляясь к противоположным полюсам клетки, и между ними образуется веретено. В электронном микроскопе каждая центриоль имеет вид полого цилиндра, в стенку которого заложены 9 групп продольных трубочек, по 3 трубочки в каждой группе (рис. 25). В типичном случае длинные оси цилиндров двух центриолей расположены перпендикулярно друг другу.
Когда клетка начинает делиться, центриоли расходятся к ее противоположным концам. От каждой центриоли отходят тонкие нити в виде лучей, образующие звезду, а между расходящимися центриолями протягиваются белковые нити, свойства которых сходны со свойствами сократимого белка мышц — актомиозина. Эти нити расположены в виде двух конусов, обращенных друг к другу основаниями, образуя веретено, сужающееся к концам, или полюсам (около центриолей), и расширяющееся к середине, или экватору. Нити веретена тянутся от экватора к полюсам, так что веретено представляет собой единую внутриклеточную структуру: можно ввести в клетку тонкую иглу и перемещать с ее помощью все веретено. Применяя специальную методику, можно извлечь веретено из клетки (рис. 120). Изолированные веретена содержат белок (в основном — один вид белка), а также небольшое количество РНК. Некоторые из нитей веретена прикрепляются к центромерам хромосом, и создается впечатление, что во время митоза они толкают или тянут хромосомы к полюсам.
В электронном микроскопе при сильном увеличении нити веретена имеют вид тонких прямых полых трубочек. Во время клеточного деления они сначала удлиняются, а затем укорачиваются, но, по-видимому, не становятся при этом тоньше или толще. Это позволяет предполагать, что изменение размеров веретена происходит не за счет растягивания или сокращения, а за счет добавления к ним нового материала или его удаления. Пробовали метить движущуюся нить веретена, «прижигая» какой-нибудь ее участок ультрафиолетовыми лучами; при этом можно было видеть, что метка перемещается из области экватора к полюсу и, дойдя до конца нити, исчезает; очевидно, белковый материал добавляется к нити у экватора, движется к полюсу и там элиминируется.
Свободная поверхность некоторых клеток покрыта ресничками, и у основания каждой реснички находится базалъное тельце. Это образование очень похоже на центриоль, так как в нем тоже имеется 9 параллельных трубочек. Каждая ресничка содержит 9 продольных нитей, расположенных по периферии, и еще 2 нити, расположенные центрально. Так же как и центриоли, базальные тельца могут удваиваться.
МИТОХОНДРИИ.
Материал, находящийся внутри плазматической мембраны, но вне ядра, называется цитоплазмой. В обычном микроскопе он выглядит как полужидкое основное вещество, в котором взвешены разнообразные капельки, вакуоли, гранулы и палочковидные или нитевидные структуры. Электронная микроскопия показала, что цитоплазма представляет собой чрезвычайно сложный лабиринт из мембран (рис. 26) и заключенных в эти мембраны пространств. При исследовании тонкого среза клетки в электронном микроскопе эти мембраны имеют вид массы червеобразных полых тяжей, образующих так называемую эндоплазматич ескую сеть. В трехмерном пространстве она представляет собой систему пластинчатых мембран, заполняющих большую часть цитоплазмы. Остальная ее часть заполнена другими специализированными структурами, несущими специфические функции; это митохондрии, аппарат Гольджи, центриоли и пластиды.
Все живые клетки, как растительные, так и животные, содержат митохондрии — тельца величиной 0,2—5 мкм, форма которых варьирует от сферической до палочковидной и нитевидной. Число митохондрий в одной клетке может быть очень разным: от нескольких штук до более чем тысячи. При исследовании живых клеток можно видеть, что митохондрии движутся, изменяют свои размеры и форму, сливаются между собой в более длинные образования или распадаются на относительно короткие структуры. Обычно они сосредоточены в той части клетки, где обмен веществ наиболее интенсивен.
Более крупные митохондрии видны в обычный микроскоп, но детали их внутреннего строения можно выявить только с помощью электронного микроскопа. Каждая митохондрия ограничена двойной мембраной; наружный слой образует гладкую наружную поверхность, а от внутреннего слоя отходят многочисленные складки в виде параллельных, направленных к центру митохондрии выступов, которые могут встречаться, а иногда и сливаться со складками, отходящими от противоположной стороны (рис. 4). Каждый из этих слоев представляет собой элементарную мембрану и состоит в свою очередь из двойного слоя молекул фосфолипида, покрытого с обеих сторон слоем белковых молекул. Внутренние складки, называемые кри стами, содержат ферменты, участвующие в
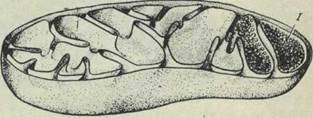
Рис. 4. Схема строения митохондрии, показывающая расположение наружной и внутренней мембран.
Внутренняя поверхность внутренней мембраны покрыта регулярно расположенными частицами, имеющими форму многогранников и соединенными с мембраной тонкой ножкой. Полагают что эти образования, называемые «субмитохондриальными частицами» (1), содержат ферменты, участвующие в окислительном фосфорилировании.
системе переноса электронов, которая играет важнейшую роль в превращении потенциальной энергии пищевых веществ в биологически полезную энергию, необходимую для осуществления клеточных функций. Полужидкое внутреннее содержимое митохондрии — матрикс — содержит ферменты цикла лимонной кислоты, или цикла Кребса. Митохондрии, главная функция которых — преобразование энергии в биологически полезную форму, иногда называют «электростанциями» клетки.
Биохимики научились гомогенизировать клетки, а затем отделять митохондрии от других внутриклеточных органелл путем дифференциального центрифугирования на больших скоростях. Эти очищенные митохондрии при инкубации их in vitro расщепляют углеводы и жирные кислоты до двуокиси углерода и воды, используя кислород и высвобождая богатые энергией фосфаты. Во время этих процессов митохондрии набухают и сжимаются.
Биологи выдвигали различные гипотезы об эволюционном происхождении митохондрий. В клетках бактерий митохондрий нет, но в них имеются мембраны, в которые как бы встроены ферменты, образующие систему переноса электронов. У некоторых бактерий эти мембраны лежат под самой плазматической мембраной. Другие бактерии, например некоторые морские формы, имеют сложную систему тонких параллельных пластинок, пересекающих центральную область клетки. В этих пластинчатых мембранах также находятся ферменты, участвующие в системе переноса электронов. Можно было бы предположить, что, по мере того как клетки становились более крупными и сложными, эти мембраны образовывали складки и в конце концов отделялись, превращаясь в обособленные органеллы — предшественники современных митохондрий. Некоторые исследователи высказывали даже мысль о том, что целые бактериальные клетки вместе со своими мембранами из ферментов, переносящих электроны, проникли в более крупные клетки и перешли к симбиотическому существованию в качестве их митохондрий.
ХЛОРОПЛАСТЫ
В клетках большинства растений имеются пластиды — небольшие тельца, в которых происходит синтез или накопление органических веществ. Наиболее важные пластиды — хлоропласты — содержат зеленый пигмент хлорофилл, который придает растению зеленую окраску и играет важнейшую роль в фотосинтезе, улавливая энергию солнечного света. Типичные хлоропласты — это дисковидные образования диаметром около 5 мкм и толщиной 1 мкм. При изучении в электронном микроскопе видно, что хлоропласты построены из мембран, плотно уложенных параллельно друг другу. Каждая клетка содержит от 20 до 100 хлоропластов, которые могут расти и делиться, образуя новые, дочерние хлоропласты. Внутри каждого хлоропласта находится множество мелких телец, называемых гранами ; эти тельца содержат хлорофилл.
Хлоропласт — это не просто мешочек, наполненный хлорофиллом. Самая способность этого пигмента улавливать энергию света зависит от его упаковки в гранах. Слой молекул хлорофилла и слой фосфолипидных молекул лежат здесь между слоями белка. Благодаря этому молекулы хлорофилла распределены по большой площади; кроме того, слоистая структура, возможно, облегчает перенос энергии с одной молекулы на другую — соседнюю с ней — во время фотосинтеза. Материал, в который погружены граны, называется стромой. Многочисленные граны одного хлоропласта соединены между собой листками мембран, проходящих через строму.
Другой тип пластид — это бесцветные лейкопласты, которые служат центрами накопления крахмала и других веществ. Пластиды третьего типа — хромопласты — содержат различные пигменты, обусловливающие окраску цветков и плодов.
РИБОСОМЫ И ДРУГИЕ ОРГАНЕЛЛЫ ЦИТОПЛАЗМЫ.
Клетки с особенно активным синтезом секретируемых белков (так называемый «синтез на экспорт»), например клетки поджелудочной железы, заполнены мембранами эндоплазматической сети, образующими сложный лабиринт; в других клетках таких мембран может быть мало. Эндоплазматическая сеть бывает двух типов: агранулярная, или гладкая, состоящая из одних только мембран, и грану лярная, или шероховатая, к мембранам которой прикреплено множество рибосом. Рибосо мы — это мелкие нуклеопротеидные частицы, на которых происходит синтез белка; они могут быть прикреплены к мембранам эндоплазматической сети или свободно взвешены в основном веществе цитоплазмы. Одна и та же клетка может содержать как гранулярную, так и гладкую эндоплазматическую сеть. Функция гладкой сети неясна; возможно, что она участвует в секреторной деятельности клетки. Плотно упакованные мембраны эндоплазматической сети иногда образуют канальцы диаметром около 50—100 нм. В других частях клетки промежутки между мембранами могут быть расширены, так что получаются уплощенные мешочки, называемые цистернами. Все эти мембраны разделяют цитоплазму на множество относительно обособленных отсеков, в которых, возможно, протекают различные химические реакции. Эндоплазматическая сеть служит также для переноса субстратов этих реакций и их продуктов через цитоплазму к наружной поверхности клетки и к ядру.
После осаждения митохондрий из гомогенизированных клеток путем дальнейшего центрифугирования можно осадить гетерогенную группу более мелких частиц, так называемых микросом; для этого нужна центробежная сила, примерно в 100 000 раз превышающая силу тяжести. Из этой микросомной фракции с помощью специальных методов (обработка соответствующими детергентами) можно выделить рибосомы. Изолированные рибосомы способны синтезировать белки in vitro, если снабдить их «инструкциями» в форме информационной РНК, набором нужных аминокислот, источником энергии, некоторыми ферментами и необходимыми транспортными РНК. Рибосомы вездесущи: их можно найти в бактериальных, растительных и животных клетках всех типов. Они содержат РНК и белок и состоят из двух субъединиц почти сферической формы, при соединении которых образуется активная структура, способная синтезировать белки. Сами рибосомы синтезируются в ядре и переходят в цитоплазму, где и выполняют свою функцию.
Во многих клетках эффективными функциональными единицами, синтезирующими белок, служат группы из 5—6 рибосом, называемые полирибосомами, или полисомами . Принято считать, что бактериальная клетка, например клетка кишечной палочки, содержит около 6000 рибосом, а ретикулоцит кролика — около 100 000. Рибосомы всех организмов, от бактерий до млекопитающих, поразительно однородны по величине, строению и химическому составу. Они содержат примерно равные количества белка и РНК и совсем или почти совсем не содержат липидов. Белок рибосом отличается довольно высоким содержанием основных аминокислот.
Одна из структурных субъединиц рибосомы имеет молекулярную массу около 1 300 000, а другая — около 600 000. Белковые компоненты рибосом из различных клеток удивительно сходны по аминокислотному составу; однако нуклеотидный состав рибосомной РНК у разных организмов значительно варьирует.
Помимо рибосом, участвующих в синтезе полипептидных цепей, микросомная фракция содержит ряд других, не столь характерных частиц, в которых находятся ферменты, связанные с метаболизмом других химических соединений.
Комплекс Гольджи — еще один компонент цитоплазмы, встречающийся почти во всех клетках, кроме зрелых сперматозоидов и эритроцитов,— представляет собой неупорядоченную сеть канальцев, выстланных мембранами. Обычно комплекс Гольджи расположен около ядра и окружает центриоли. Функция его еще не вполне ясна, но ему давно уже приписывают важную роль в секреции некоторых клеточных продуктов. Судя по некоторым данным, белки, синтезируемые в цистернах эндоплазматической сети, как бы расфасовываются в небольшие пакеты из ее мембран и направляются к комплексу Гольджи; здесь происходит переупаковка их в более крупные пузырьки, образованные из мембран этого комплекса. В этих новых «пакетах» они транспортируются к плазматической мембране, которая затем сливается с мембраной пузырька, так что, когда пузырек вскрывается, содержимое его выходит из клетки наружу. В электронном микроскопе видно, что комплекс Гольджи состоит из групп параллельно расположенных мембран, лишенных гранул; в отдельных участках промежутки между мембранами могут быть растянуты, образуя небольшие пузырьки или вакуоли, наполненные каким-то материалом. По мнению некоторых цитологов, комплекс Гольджи служит для временного хранения веществ, вырабатываемых в эндоплазматической сети, а канальцы его соединены с плазматической мембраной, что облегчает секрецию этих клеточных продуктов.
Возможно, что в растительных клетках комплекс Гольджи секретирует целлюлозу для клеточных стенок. Он обычно имеет вид обособленных, разбросанных по всей клетке телец, каждое из которых состоит из пачки уплощенных пузырьков, слегка растянутых по краям.
Лизосомы — группа внутриклеточных органелл, встречающихся в животных клетках,— сходны по величине с митохондриями, но несколько менее плотны; они представляют собой ограниченные мембраной тельца, которые содержат разнообразные ферменты, способные гидролизовать макромолекулярные компоненты клетки — белки, полисахариды и нуклеиновые кислоты. В неповрежденных клетках эти ферменты, находясь внутри лизосом, изолированы от остальной клетки, по-видимому, для того, чтобы предотвратить переваривание ими клеточного содержимого. При разрыве лизосомной мембраны они высвобождаются; этим можно, по крайней мере частично, объяснить лизис мертвых или умирающих клеток и резорбцию (рассасывание) клеток, происходящую, например, в хвосте головастика во время метаморфоза. Так как лизосомы, разрываясь, освобождают заключенные в них ферменты, способные расщеплять главные химические компоненты клетки, де Дюв назвал их «несессером самоубийцы».
Кроме этих живых элементов, цитоплазма может содержать вакуоли — полости, наполненные водянистой жидкостью и отделенные от остальной цитоплазмы вакуолярной мембраной. Вакуоли весьма обычны в клетках растений и низших животных, но редко встречаются в клетках высших животных. Большинство простейших (Protozoa) имеет пищеварительные вакуоли, которые содержат пищу, находящуюся в процессе переваривания, и сократитель ные вакуоли, «выкачивающие» из клетки избыточную воду. Наконец, цитоплазма может содержать гранулы резервного крахмала или белка, а также капельки масла.
Существует три основных структурных различия между животными и растительными клетками: 1) животные клетки в отличие от клеток высших растений имеют центриоль; 2) растительные клетки в отличие от животных клеток содержат в своей цитоплазме пластиды и 3). клетки растений обладают жесткой клеточной стенкой из целлюлозы, препятствующей изменению их положения или формы, тогда как животные клетки обычно имеют лишь тонкую плазматическую мембрану и благодаря этому способны двигаться и изменять свою форму.
Как растительные, так и животные клетки в большинстве случаев слишком малы, чтобы их можно было видеть невооруженным глазом. Их диаметр варьирует примерно от 1 до 100 мкм, а пятно диаметром 100 мкм уже находится у предела видимости. Некоторые виды амеб имеют 1—2 мм в диаметре; некоторые одноклеточные растения, например ацетабулярия, могут достигать длины 1 см и больше. К самым крупным одиночным клеткам относятся яйцеклетки рыб и птиц. Яйцеклетки крупных птиц могут достигать нескольких сантиметров в поперечнике. В курином яйце собственно клетку представляет собой только желток; белок — это неклеточный материал, выделяемый яйцеводом курицы.
Существование предельных размеров клетки обусловлено тем обстоятельством, что с увеличением размеров шара объем его возрастает пропорционально кубу радиуса, а поверхность увеличивается пропорционально квадрату радиуса. Поскольку для клеточного метаболизма необходимы кислород и питательные вещества, которые могут проникать в клетку только через ее поверхность, ясно, что величина клетки должна иметь известный предел, по достижении которого поверхность оказывается недостаточной для обеспечения метаболической активности протоплазмы. В каждом конкретном случае эта предельная величина зависит от формы клетки и от интенсивности ее метаболизма. Достигнув этой величины, клетка должна либо прекратить
рост, либо разделиться.
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ.
1. Алексеева Н., Книжка-шпаргалка, изд-во «Цитадель», М., 1995.
2. К. Вилле, В. Детье, БИОЛОГИЯ, изд-во «Мир», М., 1974.
3. К. Люцис, Краткий энциклопедический справочник, изд-во «Русское энциклопедическое товарищество», М., 2003.