| Скачать .docx | Скачать .pdf |
Реферат: Везикулярный транспорт
АО «Медицинский университет Астана»
Кафедра биологии с курсом радиобиологии и радиационной медицины.
Реферат
По молекулярной биологии
Тема: Везикулярный транспорт
План.
- Введение
- Пути транспорта белков в клетке
- Сигнальные последовательности белков
4. Транспорт в ядро.Транспорт в митохондрии и пластиды
5. Везикулярный транспорт
6. Транспорт белков из аппарата Гольджи на наружную мембрану. Экзоцитоз и трансцитоз
7. Заключение
8. Использованная литература
Введение.
Синтезируемые в цитоплазме на рибосомах белки должны попадать в разные компартменты клетки — ядро, митохондрии, ЭПР, аппарат Гольджи, лизосомы и др., а некоторые белки должны попасть во внеклеточную среду. Для попадания в определённый компартмент белок должен обладать специфической меткой. В большинстве случаев такой меткой является часть аминокислотной последовательности самого белка (лидерный пептид, или сигнальная последовательность белка). В некоторых случаях меткой служат посттрансляционно присоединённые к белку олигосахариды. Транспорт белков в ЭПР осуществляется по мере их синтеза, так как рибосомы, синтезирующие белки с сигнальной последовательностью для ЭПР, «садятся» на специальные транслокационные комплексы на мембране ЭПР. Из ЭПР в аппарат Гольджи, а оттуда в лизосомы, на внешнюю мембрану или во внеклеточную среду белки попадают путём везикулярного транспорта. В ядро белки, обладающие сигнальной последовательностью для ядра, попадают через ядерные поры. В митохондрии и хлоропласты белки, обладающие соответствующими сигнальными последовательностями, попадают через специфические белковые поры-транслокаторы при участии шаперонов.
Пути транспорта белков в клетке
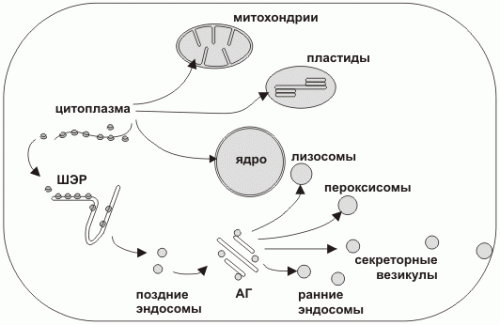
Пути транспорта в клетке
Синтез белка всегда начинается в цитоплазме. Окончание синтеза происходит в цитоплазме либо на шероховатом эндоплазматическом ретикулуме (ШЭР).
Можно условно выделить два пути транспорта белка в клетке:
1. Из цитоплазмы в некоторые органеллы (ядро, пластиды, митохондрии)
2. Большой путь везикулярного транспорта из ШЭР через аппарат Гольджи (АГ) к другим органеллам (лизосомы, пероксисомы) и через секреторные везикулы во внеклеточную среду. Поскольку синтез всех белков начинается в цитоплазме, а конечная локализация каждого белка может быть различна внутри полипептида имеется система сигналов определяющая его транспортный путь. Первичный сигнал определяет путь из цитоплазмы (в ШЭР, в ядро, в митохондрию или в пластиду), вторичный сигнал определяет дальнейшее направление, например, внешняя или внутренняя мембрана митохондрии или матрикс; лизосома, пероксисома или секреторная гранула.
Сигнальные последовательности белков
Сигнальные последовательности имеют длину 3-80 аминокислот узнаются специфическими рецепторами на мембранах различных компартментов клетки.
Сигнальная последовательность ЭР - гидрофобный участок 5-15 аминокислот на N-конце полипептида.
Сигнал митохондриальных белков 20-80 аминокислот состоящий из спирали и торчащих концов - (+)-заряженного и гидрофобного. 5 (+)-заряженных аминокислот для транспортировки в ядро. Пероксисомные белки имеют последовательность на С-конце Ser-Lys-Leu-COOH.
Имеется класс сигнальных последовательностей которые не позволяют белку достигшему определенной локализации транспортироваться дальше. Например, мотив Lys-Asp-Glu-Leu-COOH (KDEL) не позволяет белкам покидать эндоплазматический ретикулум.
Одна из функций гладкого ЭР - удержание кальция готового для выпуска в цитозоль при стимуляции клетки. Кальретикулин - белок удерживающий ионы кальция. Первые 17 аминокислот включают 14 гидрофобных (синие) - сигнальная последовательность для проникновения в ЭР из цитозоля. Последние четыре аминокислоты KDEL удерживают белок в ЭР.
(NH2)MLLSVPLLLGLLGLAVAEPAVYFKEQFLDGDGWTSRWIESKHKSDFGKFVLSSGKF
YGDEEKDKGLQTSQDARFYALSASFEPFSNKGQTLVVQFTVKHEQNIDCGGGYVKLFP
NSLDQTDMHGDSEYNIMFGPDICGPGTKKVHVIFNYKGKNVLINKDIRCKDDEFTHLYTLIVRP
DNTYEVKIDNSQVESGSLEDDWDFLPPKKIKDPDASKPEDWDERAKIDDPTDSKP
EDWDKPEHIPDPDAKKPEDWDEEMDGEWEPPVIQNPEYKGEWKPRQI
DNPDYKGTWIHPEIDNPEYSPDPSIYAYDNF
GVLGLDLWQVKSGTIFDNFLITNDEAYAEEFGNETWGVTKAAEKQMKDKQDEEQRL
KEEEEDKKRKEEEEAEDKEDDEDKDEDEEDEEDKEEDEEEDVPGQAKDEL(COOH)
Некоторые белки имеют различные локализации в клетки. Существует несколько путей транспортировки идентичных полипептидов в различные компартменты клетки [Karniely, 2005]:
1. Несколько сигнальных последовательностей в одном полипептиде преднозначенные для разных компартментов. Каталаза А дрожжей имеет две сигнальные последовательности - для митохондрий и пероксисом, причем количество фермента в этих органеллах зависит от состава среды. Некоторые цитохромы имеют два сигнала - митохондриальный и ЭР. Митохондриальный сигнал запускается после посттрансляционного фосфорилирования белка. Известно, что белок-предшественник амилоида болезни Альцгеймера также имеет два сигнала локализации - ЭР и митохондрий.
2. Одна сигнальная последовательность узнается различными рецепторами на поверхности компартментов. Например, некоторые белки митохондрий и хлоропластов имеют общую сигнальную последовательность, которая более гидрофобна чем специфические сигналы.
3. Сигнал может быть блокирован другим белком. Апуриновая/апиримидиновая эндонуклеаза 1 (Apn1) - основной фермент эксцизионной репарации репарации ДНК в ядре и митохондриях. С-конец имеет сигнал ядерной локализации (NLS), за которым идет сигнал митохондриальной локализации. белок Pir1 взаимодействует с С-концом Apn1 блокируя NLS.
4. Сигнал может быть блокирован специфическим сворачиванием белка. Аденилат-киназа дрожжей Aky2 локализуется в цитоплазме и в небольшом количестве в межмембранном пространстве митохондрий, имеет две сигнальные последовательности, активность которых зависит от конформации белка.
5. Сигнал может быть блокирован после модификации полипептида. Фосфорилированный цитохром CYP2B1, взаимодействует с цитозольным шапероном Hsp70, что приводит к конформационным изменениям и переключает одну сигнальную последовательность на другую.
6. Одна РНК может иметь два сайта инициации трансляции при этом образуются два белка - один с сигнальной последовательностью, другой без нее, что определит различную локализацию белков в клетке. В другом случае может образовываться две различные РНК кодирующие два идентичных белка, но у одного будет сигнальная последовательность, а у другого нет.
Транспорт в ядро
Транспорт в митохондрии и пластиды
Митохондрии и пластиды имеют собственную ДНК и самостоятельно синтезируют некоторые белки. Однако многие из основных белков митохондрий и пластид синтезируются в цитозоле.
Белки проникающие в митохондрии должны нести сигнал, определяющий локализацию - внутрення или наружная мембрана, или матрикс.
Белки преднозначенные для матрикса несут сигнал на N-конце, который узнается рецепторами на внешней мембране. Рецептор связан с комплексом переноса белка, который разворачивает белок и переносит его через мембрану. После переноса белка сигнальная последовательность отрезается и белок снова сворачивается.
Белки шапероны связываются с вновь синтезированным белком предотвращая его сворачивание.
Шаперонины связываются с белком после его транспортировки к месту доставки и способствуют правильному сворачиванию.
В ответ на различные стрессовые воздействия (например повышение температуры) в клетке синтезируются шапероны называемые белками теплового шока - hsp (heat-shock proteins), которые стабилизируют клеточные белки. Hsp обнаружены во всех клеточных компартментах эукариот и у бактерий.
Везикулярный транспорт
Из одной органеллы в другую перемещение происходит в везикуле или на ее поверхности в виде интегральных белков.
Донорый компартмент – органелла от которой отрывается мембрана в составе везикулы, акцепторный компартмент – принимает везикулу.
конститутивная секреция – происходит постоянно и не зависит от внешних сигналов.
регулируемая секреция – под ПМ происходит накопление пузырьков, которые сливаются с ПМ при наличии внешних сигналов – гормоны, нервы – и повышении конц. Ca2+ до 1мкм
ретроградный транспорт – возвращение рецепторных белков и липидов из АГ в Эр - восполнение мембраны ЭР.
антероградный транспорт – растворимые грузовые белки двигаются по секреторному пути ЭР. Окаймленные везикулы - покрыты белками, кот узнают и концентрируют специфич. м-ные белки и отделяют м-ну пузырька, формируют решетку и придают форму везикуле: клатриновые, COPI, COPII:
Клатриновые везикулы – ~0,1мкм, транспорт из АГ и ПМ,клатрин - 3типа, 3 большие и 3 малые субъединицы формирующие трискелетон – собирающиеся на поверхности м-ны со стороны цитоплазмы в пента- и гексагоны, кот спонтанно формируют сферу. Адаптин – связывает клатрин с м-ной и ловит различные трансм-ные белки в том числе грузовые рецепторы, кот. захватывают р-римые грузовые белки, кот попадают внутрь везикулы. Имеетя по крайней мере 4 типа адаптинов
динамин - GTP-аза, р-римый цитоплазматический белок, образует кольцо на отделяющейся клатриновой везикуле – регулирует кол-во клатрина отщепляющееся вместе с м-ной в составе везикулы, ассоциирует другие белки помогающие выпучить м-ну и белки модификаторы липидов, изменяющие локально липидный состав м-ны для выпучивания
После отделения везикулы от м-ны клатрин и адипин отделяют шапероны - ATP-азы hsp70 семейства. Ауксилин – прикрепляется к везикуле и активирует АТФ-азу. Т.к кайма формирующейся везикулы сущ. дольше чем кайма отделенной – имеется стабилизирующий механизм. Клатриновая оболочка обеспечивает значительную силу для изгибания м-ны, т.к. везикулы из внутриклеточных компартментов образуются на уже выпученной м-не
COP-I – транспорт от АГ и ЭР, 8субъединиц, GTP-белок – фактор рибозилирования АДФ –ARF – транспорт
COP-II – транспорт из АГ и ЭР, 5 субъединиц
Везикулы мб не только сферические, часто образуются трубчатые везикулы в которых высокое соотношение S/V
Образование клатриновых и COP везикул регулируется GTP-связывающими белками, которые могут находится в активном GTP- и неактивном GDP-состоянии
Два класса белков обменивают GDP-GTP: GEF-гуанин-нуклеотид-фактор обмена активирует белки заменяя GDF?GTF, GAP- белок активирующий GTP-азы – инактивирует GTP-связывающие белки меняя GTP?GDP.
GTP-азы необходимые для сборки окаймленных везикул перед сборкой пузырьков: мономерные GTP-связывающие белки (GTP-азы):
ARF-белки – необх для клатриновой и COP сборки на пов-ти м-ны АГ. Sar1 белок, необходим для COPII сборки на на ЭР м-не
тримерные (G белки).
GTP-азы находятся в цитозоле в неактивном состоянии, перед сборкой GEF встраивается в м-ну ЭР и связывает цитозольный SarI, кот обменивает GDF?GTP. В GTP состоянии SarI встраивается остатком жирной к-ты в м-ну ЭР. Ассоциирует белки об-ки и инициирует отпочковывание везикулы. GTP-азы попавшие в м-ну активируют фосфолипазу D, кот преобразует фосфолипиды в фосфотидную к-ту, что усиливает связывание оболочных белков. Вместе белок-белковые и белок-липидные взаимодействия изгибают м-ну
SNARE – белки – отвечают за слияние донорной и акцепторной м-н, более 20, каждая на специфич пов-ти м-ны, трансмембранные белки на пов-ти везикулы - v-SNAR, на пов-ти донора – t-SNAR. Взаимодействуя v- и t-SNAR обвиваются др на друга в транс-SNAR-комплекс, обеспечивающий слияние м-н. SNF-белок разрушает транс-SNAR-комплексы – цитозольный шаперон ATP-аза, использует адаптирующие белки для связывания с комплексом-SNAR
Rab-белки – мономерные GTP-азы, более 30, каждая органелла имеет хотя бы один Rab на м-не со стороны цитоплазмы, регулируют стыковку везикул и связывание v-SNAR-ов и t-SNAR-ов необходимых для слияния м-н. В состоянии GDP-не активны, нах в цитозоле, в состоянии GTP-активны и переходят на пов-ть м-ны органеллы или везикулы. В активном состоянии Rap связываются с м-ной липидным якорем и собирают другие белки участвующие в слиянии м-н
неактивный Rab-GDP связан с GDI – GDP-диссоциирующий ингибитор. Rab-GDP связывается с GEF-гуанин нуклеотид меняющий фактор, связанный с м-ной донорного компартмента – меняет GDP на GTP. Rab-GTP связывается с м-ной формирующейся везикулы и ассоциирует v-SNARE, которые в составе везикулы транспортируются к органелле и связываются с Rab-эффекторами и t-SNARE, связанными с м-ной акцепторного компартмента и обеспечивают слияние м-н
белок органелла
Rab1 ЭР и АГ
Rab2 цис-АГ
Rab3A синаптич везикулы, секрет гранулы
Rab4 ранние эндосомы
Rab5A ПМ, клатриновые везикулы
Rab5C ранние эндосомы
Rab6 промежуточный- и транс-АГ
Rab7 поздние эндосомы
Rab8 секреторные везикулы (базолатеральные)
Rab9 поздние эндосомы, trans-АГ
Слияние м-н происходит не только при везикулярном транспорте: слияние спермия с яйцом, слияние миобластов во время развития мышечной клетки.
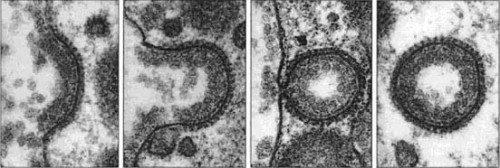 Образование клатринового пузырька. Диаметр клатринового пузырька ~0,3 мкм
Образование клатринового пузырька. Диаметр клатринового пузырька ~0,3 мкм
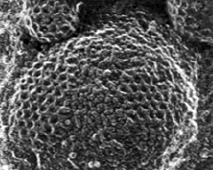 клатриновая везикула
клатриновая везикула
Транспорт белков из аппарата Гольджи на наружную мембрану
Белки, встроившиеся в мембрану ЭПС и попавшие оттуда в составе везикул в АГ, могут перемещаться на наружную мембрану клетки. Их направление к мембране осуществляется благодаря взаимодействию везикул с микротрубочками цитоскелета и благодаря особым стыковочным белкам, которые обеспечивают слияние везикул с мембраной
Экзоцитоз и трансцитоз
Экзоцитоз есть как у эукариот, так иу прокариот. Экзоцитоз (от греч. Έξω — внешний и κύτος — клетка) у эукариот — клеточный процесс, при котором внутриклеточные везикулы (мембранные пузырьки) сливаются с наружной клеточной мембраной. При экзоцитозе содержимое секреторных везикул (экзоцитозных пузырьков) выделяется наружу, а их мембрана сливается с клеточной мембраной. Практически все макромолекулярные соединения (белки, пептидные гормоны и др.) выделяются из клеток эукариот этим способом.
У прокариот везикулярный механизм экзоцитоза не встречается, у них экзоцитозом называют встраивание белков в клеточную мембрану (или в наружную мембрану у грамотрицательных бактерий), выделение белков из клетки во внешнюю среду или в периплазматическое пространство [4] .
Экзоцитоз может выполнять различные задачи:
- доставка на клеточную мембрану липидов, необходимого для роста клетки;
- доставка на клеточную мембрану мембранных белков, таких как рецепторы или белки-транспортёры. При этом часть белка, которая была направлена внутрь секреторной везикулы, оказывается выступающей на наружной поверхности клетки;
- выделение различных веществ из клетки; это могут быть, например, непереваренные остатки пищи у фаготрофных протистов, пищеварительные ферменты у животных с полостным пищеварением, белки межклеточного вещества у животных и материал клеточной стенки у растений, сигнальные молекулы (гормоны или нейромедиаторы).
У эукариот различают два типа экзоцитоза:
- Кальций-независимый конститутивный экзоцитоз встречается практически во всех эукариотических клетках. Это необходимый процесс для построения внеклеточного матрикса и доставки белков на внешнюю клеточную мембрану. При этом секреторные везикулы доставляются к поверхности клетки и сливаются с наружной мембраной по мере их образования.
- Кальций-зависимый неконститутивный экзоцитоз встречается, например, в химических синапсах, где служит для выделения нейромедиаторов. При этом типе экзоцитоза секреторные пузырьки накапливаются в клетке, а процесс их высвобождения запускается по определённому сигналу, опосредованному быстрым повышением концентрации ионов кальция в цитозоле клетки. В пресинаптических мембранах процесс осуществляется специальным кальций-зависимым белковым комплексом [w:[SNARE|]] .
Заключение.
Подготовив работу на тему «везикулярный транспорт» я поняла, что это очень важный и сложный процесс.
Сложная организация эукариотических клеток требует налаженных механизмов внутриклеточного везикулярного транспорта. Новейшие исследования показали, что механизмы, лежащие в основе таких функционально важных процессов как эндо- и экзоцитоз уникальны и, сохранившись в процессе эволюции, эффективно действуют как в клетке дрожжей, так и в нейроне гиппокампа. Как эндоцитоз лиганд-рецепторного комплекса с поверхности плазматической мембраны, так и транспорт вновь синтезируемых секреторных белков из эндоплазматического ретикулума через цис-, медиал-, транс- Гольджи к поверхности плазматической мембраны осуществляются в везикулах. Транспортные везикулы формируются и отпочковываются от донорной мембраны и после осуществления раунда внутриклеточного транспорта сливаются с акцепторной мембраной. Специализированные белки цитоплазмы покрывают вновь образованные везикулы. Согласно современным представлениям, формирование транспортной везикулы на мембране внутриклеточного компартмента начинается после взаимодействия белков, переносимых везикулой, с трансмембранным рецептором. Изменение структурного состояния связанного рецептора может распознаваться цитоплазматическими белками, которые ассоциируются с мембраной и инициируют образование транспортной везикулы.
Используемая литература:
1. Альбертс Б., Брей Д. и др. Молекулярная биология клетки. – М., 1994.
2. Горышина Е.Н., Чага О.СЮ. Сравнительная гистология тканей внутренней среды с основными иммунологами. – Л., 1990.
3. Заварзин А.А. Основы сравнительной гистологии. – Л., 1985.
4. Балахонов А.В. Ошибки развития. - Л., 1990.
5. Гилберт С. Биология развития: в 3-х т. – М., 1993-95.
6. Светлов П.Г. Физиология (механика) развития. - Л., 1978. т.1, 2.
7. Станек И. Эмбриология человека. – Братислава, 1977.
8. Юрина Н.А., Торбек В.Э., Румянцева Л.С. Основные этапы эмбриогенеза позвоночных животных и человека. – М., 1984.