| Скачать .docx | Скачать .pdf |
Курсовая работа: Отдел покрытосеменные цветковые
Кемеровский государственный сельскохозяйственный институт
Кафедра биологии
КУРСОВАЯ РАБОТА
НА ТЕМУ:
Отдел покрытосеменные (цветковые)
КЕМЕРОВО-2009
СОДЕРЖАНИЕ
ВВЕДЕНИЕ
1 ОБЩАЯ ХАРАКТЕРИСТИКА ОТДЕЛА ПОКРЫТОСЕМЕННЫХ
1.1 Происхождение покрытосеменных
1.2 Анатомо-морфологическая характеристика отдела покрытосеменных
2 СИСТЕМАТИКА ОТДЕЛА ПОКРЫТОСЕМЕННЫХ
2.1 Краткая характеристика классов
2.2 Жизненный цикл покрытосеменных
3 РОЛЬ ПОКРЫТОСЕМЕННЫХ В ПРИРОДЕ И ЖИЗНИ ЧЕЛОВЕКА
ЗАКЛЮЧЕНИЕ
СПИСОК ИСПОЛЬЗУЕМОЙ ЛИТЕРАТУРЫ
ПРИЛОЖЕНИЯ
ВВЕДЕНИЕ
Покрытосеменные – самый крупный тип растений, к которому относится более половины всех известных видов, характеризуются рядом четких, резко отграничивающих их признаков.
Наиболее характерно для них наличие пестика, образованного одним или несколькими плодолистиками (макро- и мегаспорофиллами), сросшимися своими краями, так что в нижней части пестика образуется замкнутое полое вместилище – завязь, в которой развиваются семяпочки (макро- и мегаспорангии).
После оплодотворения завязь разрастается в плод, внутри которого находятся развившиеся из семяпочек семена (или одно семя).
Кроме того для покрытосеменных характерны: восьмиядерный, или производный из него, зародышевый мешок, двойное оплодотворение, триплойдный эндосперм, образующийся только после оплодотворения, рыльце у пестика, улавливающее пыльцу, и для подавляющего большинства – более или менее типичный цветок с околоцветником.
Из анатомических признаков для покрытосеменных характерно наличие настоящих сосудов (трахей), тогда как у голосеменных развиты только трахеиды, а сосуды встречаются крайне редко. Ввиду большого количества общих признаков можно предположить монофилетическое происхождение покрытосеменных от какой-то более примитивной группы голосеменных.
Цель курсовой работы – рассмотреть основные аспекты морфологии и биологии отдела покрытосеменные.
Объект исследования – отдел покрытосеменные.
Предмет исследования – основные аспекты морфологии и биологии отдела покрытосеменные.
Поставленная в работе цель исследования обусловила ряд задач, а именно:
- рассмотреть происхождение покрытосеменных;
- изучить анатомо-морфологическую характеристику отдела покрытосеменных;
- рассмотреть краткую характеристику классов покрытосеменных;
- изучить жизненный цикл покрытосеменных;
- рассмотреть роль покрытосеменных в природе и жизни человека.
1 ОБЩАЯ ХАРАКТЕРИСТИКА ОТДЕЛА ПОКРЫТОСЕМЕННЫХ
1.1 Происхождение покрытосеменных
Наиболее ранние и очень отрывочные ископаемые остатки покрытосеменных (пыльца, древесина) известны из юрского геологического периода. Из нижнемеловых отложений известны тоже немногочисленные достоверные остатки покрытосеменных, а в отложениях середины мелового периода они встречаются сразу в больших количествах и в значительном разнообразии форм, которые все принадлежат ко многим различным ныне живущим семействам и даже родам.
В качестве предполагаемых предков покрытосеменных указывали различные группы нижестоящих в системе растений – кейтониевые, семенные папоротники, беннеттиты, гнетовые.
У кейтониевых были завязь, рыльце, но у них завязь формировалась иначе, чем у покрытосеменных; у них не было даже подобия цветков, спорофиллы их простые и, вероятно, они представляют слепую ветвь эволюции. У беннеттитов были обоеполые своеобразные «цветки», но не было пестиков, и семена их были лишь скрыты между бесплодными чешуями, а не находились внутри плодов, образованных мегаспорофиллами. У семенных папоротников не было цветков, не было покрытосемянности.
Теория происхождения покрытосеменных из гнетовых предполагает, что наиболее примитвные покрытосеменные имели мелкие однополые цветки без околоцветника или с невзрачным околоцветником. Но по ряду соображений в настоящее время более примитивными цветками считают крупные, обоеполые цветки.
Поэтому можно предположить, что предками современных покрытосеменных были какие-то вымершие, очень примитивные голосеменные с обоеполыми цветками типа шишки (стробилами), в которых на длинном цветоложе (оси) были спирально расположены свободные (несросшиеся друг с другом) листочки однородного околоцветника, микроспорофиллы (тычинки) и мегоспорофиллы (плодолистики). В системе голосеменных эта группа должна была стоять где-то между семенными папоротниками и уже более специализированными беннеттитами и саговниками.
Покрытосемянность несомненно представляла большое преимущество в смысле защиты семяпочек и развивающихся семян от всяких неблагоприятных внешних воздействий и в первую очередь от сухости воздуха. Но одной покрытосемянностью все же трудно объяснить быстрое мощное развитие покрытосеменных и вытеснение ими господствовавших ранее на земле архегониальных растений.
Русский ботаник М. И. Голенкин высказал (в 1927 г.) интересную гиппотезу о причинах победы покрытосеменных в борьбе за существование. Он предполагает, что в середине мелового периода по каким-то общим космогоническим причинам по всей Земле произошло резкое изменение освещения и влажности воздуха. Густые облака, окутовавшие ранее постоянно Землю, рассеялись и дали доступ ярким солнечным лучам, в связи с чем резко увеличилась сухость воздуха. Громадное большинство высших архегониальных растений того времени, не приспособленных и не сумевших приспособиться к яркому освещению и сухости воздуха, начало вымирать или резко сократило области своего распространения (кроме хвойных, наиболее ксерофитных).
Наоборот, покрытосеменные, имевшие до того очень ограниченное распространение и представление небольшим числом форм, выработали способность хорошо переносить яркий солнечный свет и сухость воздуха. Это обстоятельство, а также чрезвычайная эволюционная пластичность их, способность к возникновению разнообразнейших приспособлений к различным внешним условиям и обусловили быстрое победное распространение покрытосеменных по всей Земле и вытеснение ранее господствовавших групп высших архегониальных растений.
Цветковые, или покрытосеменные – сравнительно молодая группа высших растений, появившаяся в меловом периоде, стремительно размножившаяся в середине – конце мела и с тех пор занимающая господствующее положение на суше. Проблеме происхождения цветковых посвящено огромное количество публикаций, однако в этом вопросе по-прежнему много неясного. Общепризнано, что цветковые произошли от голосеменных.
Д. Д. Соколов и А. К. Тимонин подробно рассмотрели основные теории происхождения покрытосеменных. Авторы обращают внимание на парадоксальный факт: цветы покрытосеменных до такой степени разнообразны, что не существует ни одного признака или структурной особенности, которая была бы присуща всем цветкам без исключения. Поэтому и сформулировать, что такое цветок, весьма трудно. Тем не менее, ясно, что именно появление цветка было ключевым событием в происхождении покрытосеменных.
Впрочем, если не рассматривать мужские цветки, у остальных все-таки есть общая черта, отличающая цветок покрытосеменных от фруктификаций голосеменных.
У цветковых плодолистики свернуты и образуют, во-первых, рыльце, улавливающее пыльцу, во вторых – замкнутую полость (завязь), в которой развиваются семязачатки.
У некоторых голосеменных есть нечто похожее – замкнутые капсулы с семязачатками, однако у них нет завязи (пыльцу улавливают сами семязачатки при помощи длинных отростков, высовывающихся из узкого отверстия капсулы).
К тому же исходный листочек, несущий семязачатки (мегаспорофилл, у цветковых ему соответствует плодолистик) свернут у голосеменных нижней стороной внутрь, а у цветковых – наоборот, верхней стороной внутрь. Соответственно и семязачатки сидят на разных сторонах листочка.
Эти различия представляются настолько радикальными, что большинство ботаников не видят возможности постепенного перехода из одного состояния в другое и предполагают сальтационное превращение (одним скачком).
Это вполне возможно, если учесть, что развитие фруктификаций у растений управляется небольшим числом генов-регуляторов, мутации в которых действительно могут приводить к скачкообразным изменениям плана строения. Аналогичная ситуация наблюдается и у животных, план строения которых в основном определяется небольшим набором генов-регуляторов.
В начале XX века была популярна теория происхождения цветковых от беннеттитов – вымершей группы голосеменных, у которых имелись обоеполые фруктификации (стробилы) – в отличие от других голосеменных, имеющих однополые фруктификации.
Из стробилов беннеттитов, действительно напоминающих цветы, ботаники и пытались вывести типичный для покрытосеменных обоеполый цветок. Но тут возникла проблема: у беннеттитов не обнаружилось никаких структур, из которых можно было бы вывести плодолистики. У них вообще нет мегаспорофиллов, а семязачатки сидят прямо на оси стробила. Проблема казалась неразрешимой.

Рисунок 1. Лист беннеттитового Pterophyllum (поздний триас)
В 80-х годах XX века крупнейший российский палеоботаник С. В. Мейен выдвинул гипотезу, разрешающую это противоречие. Он предположил, что у каких-то беннеттитовых сальтационным путем произошла перестройка программы развития фруктификации. В результате ключевые элементы программы развития мужских элементов стробилы (микроспорофиллов, у цветковых это тычинки) стали реализовываться при развитии женских элементов.
Это привело к появлению химерных образований – микроспорофиллов, несущих семязачатки вместо пыльников. Именно из таких «мутантных» микроспорофиллов и образовались плодолистики. У беннеттитовых микроспорофиллы несут пыльники как раз на своей верхней стороне, в отличие от других голосеменных. Поэтому такая мутация сразу привела к появлению «правильных» прототипов для формирования плодолистиков, с семязачатками на нужной стороне.
Ничего невероятного в таком сценарии нет, исходя из того, что известно об общих принципах генетической регуляции развития. Гипотеза Мейена объясняет противоречие, казавшееся неразрешимым. Ее нельзя проверить классическими методами палеоботаники и морфологии (поскольку никаких переходных форм гипотеза Мейена не предполагает). Однако ее можно проверить методами сравнительно-генетического анализа.
Генетическая регуляция развития фруктификаций у семенных растений стала активно изучаться в начале 90-х годов XX века. Оказалось, что у всех цветковых главным регулятором развития цветка является ген LFY.
У голосеменных есть аналогичный ген (Leaf family), который экпрессируется (работает) на ранних стадиях развития мужских фруктификаций. Кроме того, у голосеменных есть еще один сходный ген (Needle family), отсутствующий у цветковых, который участвует в формировании женских фруктификаций. У большинства голосеменных, напомним, фруктификации раздельнополы. При появлении цветковых, таким образом, «женский ген» был утрачен, а «мужской» стал руководить развитием всего цветка, с его мужскими и женскими частями.
На этом основании Фролик и Паркер (Frolich, Parker, 2000) выдвинули «преимущественно мужскую теорию» происхождения цветка. Они предположили, что в результате утраты гена из семейства Needle у предков цветковых женские фруктификации перестали формироваться, а семязачатки стали развиваться на верхней стороне плоских микроспорофиллов. Казалось бы, это в общем то же самое, что предполагал С. В. Мейен.
Авторы «преимущественно мужской теории», однако, подчеркивают, что Мейен предполагал перенос признаков мужских структур на женские, а они говорят о переносе органов с места на место (семязачатков на микроспорофиллы). Но это, конечно, больше похоже на игру в слова, а не на существенное различие двух теорий. Различие на самом деле в другом, а именно в вопросе о предковой группе цветковых.
Эволюционные реконструкции, основанные на морфологии, подтверждали родство покрытосеменных и беннеттитовых (а также родственных беннеттитам гнетовых, которые дожили до наших дней). Однако реконструкции, основанные на молекулярных данных, показали иное. Они показали, что все дожившие до современности голосеменные представляют собой монофилетичную группу (т.е. группу, происходящую от одного общего предка и включающую всех без исключения потомков этого предка).
Это значит, что последний общий предок всех современных голосеменных, в том числе гнетовых (а стало быть и беннеттитов тоже) не был предком покрытосеменных.
Таким образом, предка цветковых нужно искать среди других вымерших голосеменных, более древних, точнее, обособившихся раньше, чем беннеттитовые. Фролик и Паркер предложили свою кандидатуру – користоспермовые семенные папоротники (Corystospermataceae). У них, правда, микроспорангии (пыльники) располагались на нижней, а не на верхней стороне микроспорофиллов.
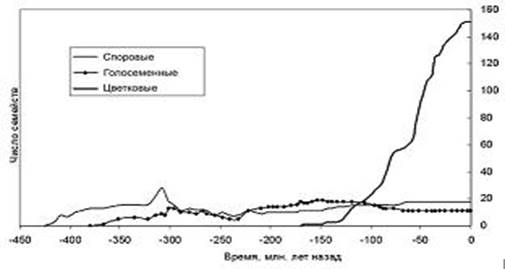
Рисунок 2. Динамика разнообразия основных групп наземных растений во времени
Д. Д.Соколов и А. К.Тимонин также приводят доводы в пользу того, что строение семязачатков користоспермовых не позволяет вывести из них семязачатки покрытосеменных. «Подходящие» семязачатки есть у другой группы голосеменных – у кейтониевых, но зато у них нет ничего похожего на плоские микроспорофиллы, из которых можно было бы вывести плодолистики.
Авторы предполагают, что причина всех этих противоречий, вероятно, кроется в недостаточной надежности эволюционных реконструкций, основанных на молекулярных данных. По их мнению, голосеменные растения плохо подходят для молекулярно-филогенетического анализа, поскольку большинство семейств голосеменных давно вымерло. Мы, таким образом, имеем недостаточно репрезентативную выборку таксонов, что, как хорошо известно, может приводить к ошибочным молекулярным филогениям. В конечном счете, по мнению авторов, на сегодняшний день нет оснований для безоговорочного отрицания идеи Мейена о происхождении цветковых от беннеттитовых. Эту идею вполне можно сохранить в качестве рабочей гипотезы.
По вопросу о месте первоначального возникновения покрытосеменных высказывались разные предположения. Одни считают, что они впервые появились на гипотетическом тропическом материке, расположенном между Америкой, Азией и Австралией и впоследствии погрузившемся в воды Тихого океана. Другие считают колыбелью их области современной арктической суши, третьи – горы субтропической и умеренно теплой зоны северного полушария. Большинство ботаников в настоящее время считают, что первичные покрытосеменные были древесными растениями, имевшими невысокие стволы, моноподиально разветвлявшиеся на немногочисленные толстые ветви.
Из них уже развились более крупные симподиально ветвящиеся деревья с многочисленными толстыми и тонкими ветвями. Из древесных же форм в разное время и разных филогенетических линиях развивались кустарники, полукустарники и травянистые формы, вначале многолетние, затем в различных родах в связи со специфическими условиями климата и местообитаний – двухлетники и однолетники.
Благодаря большой пластичности покрытосеменных, у них в процессе эволюции выработалось огромное разнообразие вегетативных органов, особенно в листьях, многочисленные метаморфозы, а также бесконечное разнообразие в цветках и плодах.
Сложность и разнообразие химического состава и физиологических реакций также очень характерно для них.
Эволюция цветка, на строении которого главным образом базируется систематика покрытосеменных, говоря в общем и схематическом виде шла у них от цветков с длинным цветоложем (типа шишки) от обоеполых, актиноморфных со спиральным расположением свободных (несросшихся) и не фиксированных в числе членов, с верхней завязью и многочисленными семяпочками – к цветкам циклическим, зигоморфным, раздельнополым, со строго фиксированным числом более или менее сросшихся членов на плоском цветоложе, с нижней одногнездною завязью и немногими или одной семяпочкой. Эта эволюция цветка покрытосеменных происходила в разных эволюционных рядах их независимо друг от друга.
1.2 Анатомо-морфологическая характеристика отдела покрытосеменных
Покрытосеменные – самый крупный тип растений, к которому относится более половины всех известных видов. Наиболее характерно для них наличие пестика, образованного одним или несколькими плодолистиками (макро- и мегаспорофиллами), сросшимися своими краями, так что в нижней части пестика образуется замкнутое полое вместилище – завязь, в которой развиваются семяпочки (макро- и мегаспорангии). После оплодотворения завязь разрастается в плод, внутри которого находятся развившиеся из семяпочек семена (или одно семя).
Кроме того для покрытосеменных характерны: восьмиядерный, или производный из него, зародышевый мешок, двойное оплодотворение, триплойдный эндосперм, образующийся только после оплодотворения, рыльце у пестика, улавливающее пыльцу, и для подавляющего большинства – более или менее типичный цветок с околоцветником. Из анатомических признаков для покрытосеменных характерно наличие настоящих сосудов (трахей), тогда как у голосеменных развиты только трахеиты, а сосуды встречаются крайне редко.
За небольшими исключениями, все покрытосеменные растения имеют корень, стебель и листья.
Корень (radix) развивается под землей и служит растению для укрепления в почве и всасывания из нее воды и растворенных в ней неорганических веществ, необходимых для его нормального существования. Нормальные корни каждого растения образуют его корневую систему.
Различают два основных типа корневой системы: стержневая корневая система (дуб, наперстянка, одуванчик и др.) и мочковатая или пучковатая корневая система (все луковичные, такие, как чеснок, лук и др., а также и большинство злаков, таких, как кукуруза, пшеница и др.).
У некоторых растений корни видоизменены: мясистые корни (морковь, свекла, редька и др.), корневые клубни (большая часть болгарских ятрышников), клубневые корни (георгина, таволга, калужница и др.). Во всех видоизмененных таким образом корнях откладываются запасные пищевые вещества.
Стебель (caulis и cauloma), когда он нормальный, развивается над поверхностью земли. По стеблю проводится всосавшаяся из почвы вода с растворенными в ней неорганическими веществами наверх к листьям и возвращается вниз в корни вода с растворенными в ней органическими обработанными веществами; кроме того, на стебле и на его разветвлениях расположены листья и цветки.
Помимо нормальных надземных стеблей, у некоторых растений имеются видоизмененные подземные стебли, какими являются, например, корневища (rhizoma – касатик, ландыш майский, чемерица и др.), луковица (bulbus – подснежник, лук, чеснок и др.) и клубень (tuber – картофель, земляная груша и др.). И в видоизмененных стеблях откладываются запасные питательные вещества.
В зависимости от стойкости нормальных надземных стеблей растения делятся на два основных типа: древесные и травянистые. К первому типу причисляют деревья (дуб, черешня, береза и др.), кустарники (шиповник, терн, лещина и др.) и полукустарники (ежевика, черника и брусника, лаванда и др.).
Ко второму типу относятся многолетние травянистые растения, из подземных многолетних частей которых каждый вегетационный период вырастают надземные стебли, а на них образуются цветки, плоды и семена; зимой, однако, вся надземная часть таких растений погибает (белладонна, большинство видов горечавки, морозник, зверобой, ландыш майский и др.).
Двухлетние травянистые растения, у которых в первый год образуется прикорневая розетка листьев, во время зимы сохраняют свою подземную часть и только на второй год у них развивается стебель с цветками, плодами и семенами, однако зимой второго года они погибают (некоторые виды коровяка, иногда наперстянка шерстистая, иногда пастушья сумка, а из культурных растений – капуста, морковь и др.).
У однолетних травянистых растений за один вегетационный период из семени развивается взрослое растение, которое после образования цветков, плодов и семян, зимой или даже раньше погибает (дурман, очанка, золототысячник, а из культурных растений – кукуруза, пшеница и др.). Некоторые из однолетних растений могут перезимовать, когда семена их прорастают осенью, но это обычно наблюдается мягкой зимой.
Стебли растений бывают разветвленными (деревья, кустарники и многие травянистые) и неразветвленными (подснежник, ландыш, пшеница и др.). Разветвление у разных растений различное.
Листная система (лист – folium) является фотосинтезирующим аппаратом растения, где главным образом образуются органические вещества. Существует очень большое разнообразие, в особенности в отношении внешнего строения (морфологии) листа, а именно: прикрепление листьев и расположение их по стеблю, наличие или отсутствие черешка, влагалища или прилистников, устройство пластинки листа — простая или сложная пластинка, форма пластинки листа у простых листьев, форма края пластинки, жилкование, консистенция листьев и др.
Лист покрытосеменных растений состоит из черешка (стебелька листа), листовой пластинки (лопасти) и прилистников (парных придатков, расположенных по обеим сторонам основания черешка). Место, где черешок примыкает к стеблю, называется влагалищем листа. Угол, образованный листом (черешком листа) и вышерасположенным междоузлием стебля, называется пазухой листа. В пазухе листа может образоваться почка (которая в этом случае называется пазушной почкой), цветок (называется пазушным цветком), соцветие (называется пазушным соцветием).
Цветок (flos). Цветок обеспечивает половое размножение или воспроизводство растения. Он представляет собой укороченный стебель с ограниченным ростом; на этом стебле видоизмененные листья располагаются в определенном порядке, в котором можно установить известные закономерности. Такие видоизмененные листья называют частями цветка.
Существенными частями являются тычинки и пестик, а несущественными – чашечка и венчик или простой околоцветник (перигон), когда нет обособленных чашечки и венчика (тюльпан, ландыш, подснежник и др.). Когда один и тот же цветок имеет и тычинки, и пестик, его называют обоеполым (гермафродитным); когда у цветка только тычинки – это однополый, мужской (тычиночный) цветок, а когда налицо только пестик – это однополый, женский (пестичный) цветок.
Когда мужские и женские цветки расположены на одном и том же индивиде, растение однодомное (грецкий орех, лещина, тыква и др.), когда они расположены на двух различных индивидах – двудомное (большинство видов ивы, конопля, крапива двудомная и др.).
Части цветка в обоеполых цветках располагаются в пять кругов по цветоложу, а именно, снаружи чашечка, затем, идя внутрь, венчик, тычинки (в два круга) и пестик. При простом околоцветнике лепестки расположены чаще всего в два круга. Каждая тычинка имеет пыльник, в гнездах которого образуется пыльца (цветочная пыльца), и тычиночную нить (не всегда).
Пестик состоит из длинной расширенной части – завязи, столбика (иногда отсутствует) и одного рыльца или нескольких рылец. Пестик образуется от срастания одного или большего числа плодолистиков. Если плодолистиков два или больше и каждый из них закроется сам по себе, получается сборный или сложный пестик (малина, лютик, морозник и др.). При срастании двух или большего числа плодолистиков (вместе) может образоваться одногнездная или многогнездная завязь.
В завязи образуется одна (черешня, слива, малина, подсолнечник и др.) или больше (томат, белладонна, груша, тыква и др.) семяпочек, которые после оплодотворения и развития в них зародыша превращаются в семена. В зависимости, главным образом, от способа опыления, то есть от способа перенесения пыльцы из пыльников тычинок на рыльце пестика (насекомыми, ветром, водой, птицами и др.) цветки бывают различно устроены – существует поразительное приспособление к переносчикам пыльцы.
В отношении симметрии цветки бывают полисимметричными или актиноморфными (черешня, подснежник, дурман и др.), оносимметричными или зигоморфными (фиалка, бобовые, наперстянка и др.) и ассимметричными или неправильными (валериана, хна и др.). Махровые цветки, которые почти всегда бывают бесплодными, получаются путем превращения тычинок в лепестки.
Соцветия (inflorescentiae). Очень редко цветки располагаются по одному на стебле (подснежник, мак, тюльпан, пион и др.)- Когда несколько или много цветков, между которыми нет нормальных листьев, я только прилистники, расположены в непосредственной близости друг от друга, они образуют соцветие. Соцветия покрытосеменных растений очень разнообразны, причем нередко они бывают сложными и комбинированными. Их можно разделить на две основные группы: 1) рацемозные (кисти, бокоцветные), с моноподиальным разветвлением; 2) цимозные (верхоцветные), с симподиальным или ложным, вильчатым разветвлением.
Семя (semen) и плод (fructus). Материальной предпосылкой оплодотворения является опыление, о котором уже упоминалось выше, а оплодотворение состоит в том, что обе оплодотворяющие клетки (спермии), образующиеся при прорастании пылинки на рыльце (проходя через пыльцевую трубочку), проникают в зародышевый мешок (эмбриосак) семяпочки и сливаются – одна с яйцеклеткой, а другая с вторичным ядром. После многократного деления оплодотворенной клетки образуется зародыш, а из оплодотворенного вторичного ядра образуется вторичная питательная ткань (эндосперм).
После окончательного развития и оформления зародыша семяпочка разрастается и превращается в семя с собственной оболочкой (testa), а завязь, в которой скрыта семяпочка (или семяпочки), так же разрастается, увеличивается и превращается в плод, у которого тоже есть своя оболочка (pericarpium). Строение и размеры семян очень разнообразные (орхидеи, ива, мак, тыква, каштан конский, кокосовая пальма и др.). По своему строению и размерам плоды еще более разнообразны, «следствие чего созданы различные классификации плодов. Одной из наиболее естественных и приемлемых является классификация, созданная на основании консистенции плодов и строения оболочки плода, получающейся при разрастании плодолистика или плодолистиков (когда завязь образована от сращения вместе двух или нескольких плодолистиков). Существует два основных типа плодов: сухие, у которых весь околоплодник деревянистый, кожистый или пленчатый, и сочные или мясистые, у которых весь околоплодник или часть его (пласт или пласты) сочные или мясистые.
Сухие плоды.
1. Сухие невскрывающиеся (и нераспадающиеся). Почти все имеют по одному семени: а) орех (пих) и орешек (nucula) орешник – лещина, липа, конопля, горец птичьи и др.; б) желудь (glans – но только для плода дуба) – у дуба, бука и др.;в) зерновка (caryopsis) – у злаковых растений;
г) плодосемянка (achena или achenium) – у подсолнечника и всех растений семейства сложно¬цветных; д) однокрылатка у березы, ясеня, береста и др.;
2. Сухие вскрывающиеся (все многосемянные):
а) пузырчатка (follicujus _ вскрывается по одному шву) живокость, барвинок (Vinca) и др.; б) боб (legumen) – у большинства бобовых;
в) стручок и стручочек (siliqua и silicula) – у крестоцветных (горчица, капуста, пастушья сумка и др.).
г) коробочка (capsula) (вскрывается самыми различными способами или семена выпадают через дырочки) - у тюльпана, фиалки, дурмана, наперстянки, табака, мака и др.
3. Сухие вскрывающиеся. У них весь плод распадается на 2, 3, 4 и более частей (плодиков), в каждом из них имеется по одному семени – у большинства зонтичных, подмаренника цепкого, клещевины, губоцветных, бурачниковых, алтея, просвирника и др.
Сочные или мясистые плоды.
1. Ягода (Bасса). Весь околоплодник сочный и охватывает семена, только самая наружная часть его кожистая — у томата, винограда, белладонны и др. Сюда относят также и плоды, отличающиеся тем, что наружная часть околоплодника (кожура) более или менее плотная – у лимона, тыквы, арбуза и др.
2. Костичковый плод или костянка (drupa) чаще всего бывает –односемянным. Околоплодник состой от трех слоев: наружный – тонкий, кожистый, средний – сочный (мясистый) и внутренний – твердый, деревянистый – у черешни, сливы, маслины, барбариса и др.
Существуют сборные или сложные плоды, образованные из многих мелких плодиков, которые каждый сам по себе могут быть сухими или сочными – лютик, пион, малина, ежевика и др. Ложные плоды имеются у шиповника, клубники и др., сочная часть этих плодов образована из разросшегося цветоложа. Ошибочно названы плодом некоторые соплодия, как, например, соплодия шелковицы, инжира, ананаса и др.
У покрытосеменных к генеративным органам относят цветок, семя и плод.
2 СИСТЕМАТИКА ОТДЕЛА ПОКРЫТОСЕМЕННЫХ
2.1 Краткая характеристика классов
Покрытосеменные разделяют на два класса – двудольные и однодольные. Для двудольных характерны: две семядоли в семени, открытые проводящие пучки (с камбием), сохранение в течение всей жизни главного корня (у особей, родившихся из семян), перистое и сетчатое жилкование листьев, 5-4-2-членный тип цветков.
Однодольные характеризуются противоположными признаками: одна семядоля в семени, закрытые проводящие пучки (без камбия), раннее отмирание главного корня и развитие придаточной корневой системы, параллельное или дуговое жилкование, трехчленный тип цветков. Отдельные признаки одной группы могут встречаться и у представителей другой группы, поэтому важна вся совокупность признаков.
Однодольные растения (лат. Liliopsida, лат. Monocotyledones, англ. monocots) – класс покрытосеменных, или цветковых, растений, самым многочисленным семейством которого являются Орхидные, отличающиеся чрезвычайно сложными, красивыми цветами. На втором месте по количеству видов стоит весьма важное в хозяйственном отношении семейство Злаки.
Традиционным латинским названием для этой группы растений является Monocotyledones, хотя в последнее время, например в системе Кронквиста (Cronquist) их официальное название – Liliopsida (лилиопсиды). Так как однодольные – группа рангом выше семейства, выбор названия ничем не ограничен. Статья 16 МКБН позволяет как описательное название, так и имя, образованное от типового рода группы.
Традиционное название однодольные, Monocotyledones или Monocotyledoneae, происходит от того факта, что зародыши большинства членов группы имеют только одну семядолю в противоположность двудольным, у которых их обычно две. С диагностической точки зрения определение количества семядолей не является ни легко доступным способом, ни надёжной отличительной характеристикой растения. Различение однодольных и двудольных впервые было использовано в систематике растений ещё в начале XVIII века английским натуралистом Дж. Рэем.
Тем не менее, у однодольных имеются более наглядные отличительные признаки. Зародышевый корешок обыкновенно скоро перестаёт расти и заменяется придаточными корнями. Стеблевые сосудистые пучки замкнутые, рассеянные по всему сечению стебля; камбия нет, поэтому утолщения стеблей по типу двудольных или голосеменных не наблюдается. Стебли редко ветвятся. Листья большей частью стеблеобъемлющие, всегда без прилистников, обыкновенно узкие и дугонервные.
Цветы обыкновенно построены по тройному типу: околоцветник из двух трехчленных кругов, тычинок также 3 + 3, плодолистиков 3, реже вместо числа 3 в цветке наблюдаются числа 2 или 4.
Однодольные представляют собой монофилетическую группу, возникшую на заре истории развития покрытосеменных растений. Древнейшие ископаемые растения, которых можно отнести к однодольным, имеют возраст начала мелового периода.
Система научной классификации APG II, разработанная группой APG (англ. Angiosperm Phylogeny Group), определяет однодольные как одну из двух крупнейших групп среди покрытосеменных растений. Вторая группа -- «эудикоты» (eudicots), по устоявшейся традиции иногда называется «палеодикоты» (palaeodicots). Среди однодольных выделяются десять порядков и два семейства, которые ещё окончательно не приписаны ни к одному из порядков. Эти порядки распределены следующим образом:
Основные однодольные
· Семейство Петросавиевые (Petrosaviaceae) / en:Petrosaviaceae
· Порядок Аироцветные (Acorales) / en:Acorales
· Порядок Частухоцветные (Alismatales) / en:Alismatales
· Порядок Спаржецветные (Asparagales) / en:Asparagales
· Порядок Диоскореецветные (Dioscoreales) / en:Dioscoreales
· Порядок Лилиецветные (Liliales) / en:Liliales
· Порядок Панданоцветные (Pandanales) / en:Pandanales Commelinids
· Семейство (Dasypogonaceae) / en:Dasypogonaceae
· Порядок Пальмоцветные (Arecales) / en:Arecales
· Порядок Коммелиноцветные (Commelinales) / en:Commelinales
· Порядок Злакоцветные (Poales) / en:Poales
· Порядок Имбирецветные (Zingiberales) / en:Zingiberales
Более традиционной классификацией является система Кронквиста (1981), согласно которой все однодольные разбивались на пять подклассов со следующими порядками:
Алисматиды (Alismatidae)
· Порядок Частуховые (Alismatales)
· Порядок Водокрасовые (Hydrocharitales)
· Порядок Наядовые (Najadales)
· Порядок Триурисовые (Triuridales)
Арециды (Arecidae)
· Порядок Пальмы (Arecales)
· Порядок Циклантовые (Cyclanthales)
· Порядок Панданоцветные (Pandanales)
· Порядок Аронниковые (Arales)
Коммелиноцветные (Commelinidae)
· Порядок Коммелиноцветные (Commelinales)
· Порядок Эриокаулоновые (Eriocaulales)
· Порядок Рестиевые (Restionales)
· Порядок Ситникоцветные (Juncales)
· Порядок Осокоцветные (Cyperales)
· Порядок Гидателловые (Hydatellales)
· Порядок Рогозовые (Typhales)
Имбирные (Zingiberidae)
· Порядок Бромелиевые (Bromeliales)
· Порядок Zingiberales
Лилииды (Liliidae)
· Порядок Лилиецветные (Liliales)
· Порядок Орхидоцветные (Orchidales)
Класс Двудольные относится к отделу Цветковых (Anthophyta), или Покрытосеменных (Magnoliophyta, или Angiospermae) растений. Этот класс значительно разнообразнее и больше по объёму второго класса из этого отдела -- Однодольных (Monocotiledonae или Liliopsida). Из общего числа цветковых растений на долю Двудольных приходится около 80%.
Класс Двудольные характеризуется наличием следующих признаков, отличающих его от Однодольных:
1. Зародыш с двумя семядолями.
2. Главный корень хорошо развит и сохраняется в течение всей жизни, поэтому преобладает стержневая (реже мочковатая) корневая система.
3. Стебель способен к вторичному утолщению благодяря наличию камбия; проводящие пучки открытые.
4. Листья разнообразны по форме и рассечению, имеют пальчатое или перистое жилкование, форма края листовой пластинки может быть разной.
5. Цветки ациклические, полуциклические и циклические. Число членов каждого круга кратно 5, редко 2, еще реже 3.
К классу Двудольных относятся около 200 000 видов, 10 000 родов, около 300 семейств (в зависимости от принятой классификации). Это травянистые и древесные растения.
Систематикой цветковых растений, начиная, с 18 века занималось множество учёных-ботаников, как отечественных, так и зарубежных. Все они внесли бесценный вклад в современное построение филогенетической (естественной) системы цветковых растений. Однако, общепринятой системы классификации покрытосеменных растений до сих пор нет.
Наиболее спорным является вопрос, какие группы покрытосеменных наиболее близки к древним предковым формам. В системах известных ботаников-филогенетиков А. Энглера и Р. Веттштейна за наиболее примитивные группы принимаются семейства с однопокровными и беспокровными, невзрачными, анемофильными цветками (ивовые, березовые и др.).
В более современных системах в качестве примитивной группы рассматривают семейства с хорошо развитыми многочленными, раздельнолистными, энтомофильными цветками, так называемые многоплодниковые (семейства магнолиевые, лютиковые и др.). Семейства с однопокровными цветками считаются вторичноупрощенными. Такими системами являются системы ботаников Н. А. Буша, А. А. Гроссгейма, А. Л. Taxтаджяна, Гетчинсона (Англия) и др. Одной из последних систем, в которой учитывается наибольшее число признаков, является система А. Л. Тахтаджяна (1970).
Согласно А. Л. Тахтаджяну, класс Двудольные включает 7 подклассов: Magnoliidae, Ranunculidae, Hamamelididae, Caryophyllidae, Dilleniidae, Rosidae и Asteridae. В пределах каждого подкласса, его семейства объединены в порядки. Весь класс Двудольные включает 71 порядок. Первые по счету охватывают наиболее примитивные семейства, последние – филогенетически более продвинутые.
Основные порядки класса Двудольных:
Подкласс раздельнолепестные (Choripetalae): порядок магнолиецветные (Magnoliales), порядок лютикоцветные (Ranunculales), порядок макоцветные (Papaverales), порядок каперсоцветные (Capparales), порядок розоцветные (Rоsales), порядок бобовоцветные (Fabales), порядок мальвоцветные (Malvales), порядок гераниецветные (Geraniales), порядок терпентинные (Terebinthales), порядок зонтикоцветные (Umbellales), порядок центросеменные (Centrospermae), порядок гречихоцветные (Polygonales), порядок букоцветные (Fagales).
Подкласс спайнолепестные (Sympetalae): порядок норичникоцветные (Scrophulariales), порядок тыквенноцветные (Cucurbitales), порядок астроцветные (Asterales).
2.2 Жизненный цикл покрытосеменных
Навашин Сергей Гаврилович (1857 – 1930), цитолог и эмбриолог растений, основатель научной школы, академик АН СССР и АН Украины, открыл двойное оплодотворение (1898) у покрытосеменных растений. Заложил основы морфологии хромосом и кариосистематики.
Навашин сделал открытие величайшего биологического значения. Он установил, что процесс оплодотворения у покрытосеменных растений принципиально отличается от процесса оплодотворения у голосеменных растений тем, что у покрытосеменных в оплодотворении участвуют два спермия, один из которых сливается с яйцеклеткой и даёт начало зародышу, второй сливается с одной из клеток зародышевого мешка и даёт начало эндосперму. Следовательно, в отличие от голосеменных, эндосперм покрытосеменных растений является продуктом полового процесса. Процесс, открытый академиком С. Г. Навашиным, получил в науке название двойного оплодотворения.
Эндосперм образуется, когда два спермия из пыльцевого зерна (мужского гаметофита) прорастают до зародышевого мешка (женского гаметофита). Один спермий оплодотворяет яйцеклетку, при этом образуется зигота, а второй сливается с двумя полярными тельцами в центре зародышевого мешка, и при этом образуется первичная клетка эндосперма (с тройным набором хромосом), которая развивается в эндосперм. Такой процесс называется двойным оплодотворением.
Около 70 % покрытосемянных имеют полиплоидные эндоспермы,http://ru.wikipedia.org/wiki/%D0%AD%D0%BD%D0%B4%D0%BE%D1%81%D0%BF%D0%B5%D1%80%D0%BC - cite_note-Olsen2007-3 обычно триплоидные, но встречаются и варианты от 2n до 15n (наборов хромосом).
Древнее цветковое растение Nuphar polysepala имеет диплоидный эндосперм, результат слияния спермия с одной полярной клеткой. Считается, что на ранней стадии развития цветковых произошло изменение в данной типе развития и стал образовываться не семиклеточный, а восьмиклеточный зародышевый мешок с диплоидным эндоспермом.
Рядом точных эмбриологических исследований на различных представителях покрытосеменных (лилейных, лютиковых, сложноцветных) С. Г. Навашин убедительно показал, что эндосперм, как и зародыш, является продуктом полового процесса.
Это необычное, свойственное лишь покрытосеменным растениям, явление он назвал двойным оплодотворением. О своем открытии он сообщил в августе 1898 г. на проходившем в Киеве Х съезде русских естествоиспытателей и врачей, а в ноябре того же года опубликовал на эту тему небольшую статью в "Известиях Петербургской Академии Наук".
Мысль о существовании двойного оплодотворения возникла у С. Г. Навашина еще в 1895 г. в процессе работы над изучением халазогамии у грецкого ореха. Окончательное же подтверждение и оформление в стройную теорию эта мысль получила при исследовании оплодотворения у лилии.
Позднее С. Г. Навашин описал двойное оплодотворение и у других цветковых растений, систематически далеко отстоящих друг от друга - у представителей лютиковых, сложноцветных, орехоцветных, доказав тем самым общность этого явления для всех покрытосеменных. Двойное оплодотворение – отличительный признак, отделяющий покрытосеменных от голосеменных.
Открытие С. Г. Навашиным двойного оплодотворения у покрытосеменных сыграло важную роль в науке. Явление двойного оплодотворения внесло ясность не только в вопрос происхождения эндосперма, но и разъяснило загадочность такого явления, как ксения у кукурузы. Работы Навашина по оплодотворению покрытосеменных растений были встречены ботаниками всего мира с огромным интересом.
3 РОЛЬ ПОКРЫТОСЕМЕННЫХ В ПРИРОДЕ И ЖИЗНИ ЧЕЛОВЕКА
Покрытосеменные (или цветковые) – это крупнейший отдел растительного мира. Он насчитывает 533 семейства, 13000 родов. Почти 250 тысяч видов покрытосеменных делятся на два класса: двудольные и однодольные, различающиеся, прежде всего, по количеству семядолей в зародышах, строению листа и цветка.
Победа покрытосеменных повлекла за собой изменения и в животном населении Земли; особенно она должна была сказаться в быстрой эволюции насекомых, млекопитающих и птиц, питающихся насекомыми, затем хищных и плодоядных. В свою очередь и у покрытосеменных постепенно возникали в процессе эволюции бесчисленные приспособительные изменения формы, химизма и функций в связи со сложными и разнообразными взаимоотношениями их с животным миром.
В настоящее время покрытосеменные распространены повсюду почти до крайних пределов растительности и определяют характер ландшафтов везде, кроме хвойных лесов, торфяных болот и некоторых типов тундр.
Цветковые растения распространены по всему миру, от Арктики до Антарктики. В основе их систематики лежит строение цветка и соцветия, пыльцевых зёрен, семени, анатомия ксилемы и флоэмы. Цветковые растения являются одним из ключевых компонентов биосферы: они производят органические вещества, связывают углекислоту и выделяют в атмосферу молекулярный кислород, с них начинаются большинство пастбищных цепей питания. Многие цветковые растения используются человеком для приготовления пищи, строительства жилища, изготовления различных хозяйственных материалов, в медицинских целях.
В жизни и хозяйственной деятельности человека роль покрытосеменных неизмеримо больше, чем остальных групп растений.
Пища, одежда, фураж для скота, ароматические, наркотические, лекарственные, дубильные вещества, каучук и гуттаперча, пробка и многое другое получается из покрытосеменных; материал для жилищ, топливо поделочные материалы, бумага тоже в значительной степени поставляются покрытосеменными.
Представители отдела покрытосеменных господствуют на большей части суши нашей планеты и играют решающую роль в формировании растительного покрова. Они создают основную часть наземной фитомассы и в конечном счете определяют возможность существования человека как биологического вида.
ЗАКЛЮЧЕНИЕ
Покрытосеменные – самый крупный тип растений, к которому относится более половины всех известных видов. Наиболее характерно для них наличие пестика, образованного одним или несколькими плодолистиками (макро- и мегаспорофиллами), сросшимися своими краями, так что в нижней части пестика образуется замкнутое полое вместилище – завязь, в которой развиваются семяпочки (макро- и мегаспорангии). После оплодотворения завязь разрастается в плод, внутри которого находятся развившиеся из семяпочек семена (или одно семя).
Кроме того для покрытосеменных характерны: восьмиядерный, или производный из него, зародышевый мешок, двойное оплодотворение, триплойдный эндосперм, образующийся только после оплодотворения, рыльце у пестика, улавливающее пыльцу, и для подавляющего большинства – более или менее типичный цветок с околоцветником. Из анатомических признаков для покрытосеменных характерно наличие настоящих сосудов (трахей), тогда как у голосеменных развиты только трахеиты, а сосуды встречаются крайне редко.
За небольшими исключениями, все покрытосеменные растения имеют корень, стебель и листья.
Наиболее ранние и очень отрывочные ископаемые остатки покрытосеменных (пыльца, древесина) известны из юрского геологического периода. Из нижнемеловых отложений известны тоже немногочисленные достоверные остатки покрытосеменных, а в отложениях середины мелового периода они встречаются сразу в больших количествах и в значительном разнообразии форм, которые все принадлежат ко многим различным ныне живущим семействам и даже родам.
В качестве предполагаемых предков покрытосеменных указывали различные группы нижестоящих в системе растений – кейтониевые, семенные папоротники, беннеттиты, гнетовые.
В начале XX века была популярна теория происхождения цветковых от беннеттитов – вымершей группы голосеменных, у которых имелись обоеполые фруктификации (стробилы) – в отличие от других голосеменных, имеющих однополые фруктификации.
По вопросу о месте первоначального возникновения покрытосеменных высказывались разные предположения. Одни считают, что они впервые появились на гипотетическом тропическом материке, расположенном между Америкой, Азией и Австралией и впоследствии погрузившемся в воды Тихого океана. Другие считают колыбелью их области современной арктической суши, третьи – горы субтропической и умеренно теплой зоны северного полушария. Большинство ботаников в настоящее время считают, что первичные покрытосеменные были древесными растениями, имевшими невысокие стволы, моноподиально разветвлявшиеся на немногочисленные толстые ветви.
Покрытосеменные разделяют на два класса – двудольные и однодольные. Для двудольных характерны: две семядоли в семени, открытые проводящие пучки (с камбием), сохранение в течение всей жизни главного корня (у особей, родившихся из семян), перистое и сетчатое жилкование листьев, 5-4-2-членный тип цветков.
Однодольные характеризуются противоположными признаками: одна семядоля в семени, закрытые проводящие пучки (без камбия), раннее отмирание главного корня и развитие придаточной корневой системы, параллельное или дуговое жилкование, трехчленный тип цветков. Отдельные признаки одной группы могут встречаться и у представителей другой группы, поэтому важна вся совокупность признаков.
Навашин сделал открытие величайшего биологического значения. Он установил, что процесс оплодотворения у покрытосеменных растений принципиально отличается от процесса оплодотворения у голосеменных растений тем, что у покрытосеменных в оплодотворении участвуют два спермия, один из которых сливается с яйцеклеткой и даёт начало зародышу, второй сливается с одной из клеток зародышевого мешка и даёт начало эндосперму. Следовательно, в отличие от голосеменных, эндосперм покрытосеменных растений является продуктом полового процесса. Процесс, открытый академиком С. Г. Навашиным, получил в науке название двойного оплодотворения.
Дискуссии о сущности двойного оплодотворения продолжались и в начале ХХ в. В ходе этих дискуссий, а главное, благодаря новым исследованиям, выводы Навашина получили полное подтверждение и дальнейшее развитие.
В жизни и хозяйственной деятельности человека роль покрытосеменных неизмеримо больше, чем остальных групп растений.
Пища, одежда, фураж для скота, ароматические, наркотические, лекарственные, дубильные вещества, каучук и гуттаперча, пробка и многое другое получается из покрытосеменных; материал для жилищ, топливо поделочные материалы, бумага тоже в значительной степени поставляются покрытосеменными.
Представители отдела покрытосеменных господствуют на большей части суши нашей планеты и играют решающую роль в формировании растительного покрова. Они создают основную часть наземной фитомассы и в конечном счете определяют возможность существования человека как биологического вида.
ЛИТЕРАТУРА
1. Анучин Н.А., Атрохин В.Г., Виноградов В.Н. и др. - М.: Сов. Энциклопедия, 1986.-631 с., ил.
2. Банникова В.П., Хведынич О.А. Основы эмбриологии растений. М., 2002. 164с.
3. Баранов П. А. История эмбриологии растений. М.-Л.: Изд. АН СССР. 1955. 439с.
4. Герасимова-Навашина Е.Н. Двойное оплодотворение покрытосеменных и некоторые его теоретические аспекты. В сб.: Проблемы эмбриологии. Киев: Наукова думка. 1971. С. 113-152.
5. Гиляров М. С. Биология. Большой энциклопедический словарь. - 3 изд. – Москва: Большая Российская энциклопедия, 1999. - 864с.
6. Жизнь растений. В 6-ти т. Т. 6. Цветковые растения. / Под ред. А.Л. Тахтаджяна. – М.: Просвещение, 1982. – 543 с., ил, 34 л. ил.
7. Кордюм Е. Л. Эволюционная цитоэмбриология покрытосеменных растений. М., 2008. 213 с.
8. Лесная энциклопедия: В 2-х т., т.2/Гл. ред. Воробьев Г.И.; Ред. кол:
9. Марков А. В. Происхождение цветковых растений остается загадочным. // Журнал Общей Биологии Том 69, 2008.
10. Поддубная-Арнольди В.А. Цитоэмбриология покрытосеменных растений. М.: Наука. 2006. 486с.
11. Романов И.Д. Типы развития зародышевого мешка покрытосеменных растений. М., 2006. С. 72-113.
12. Савченко М. И. Морфология семяпочки покрытосеменных растений. Л.: Наука. 1983. 110с.
13. Соколов Д. Д., Тимонин А. К. Морфологические и молекулярно-генетические данные о происхождении цветка: на пути к синтезу // Журнал Общей Биологии Том 68, 2007. № 2, Март-Апрель. Стр. 83-97.
14. Сладков А. Н., Гревцова Н. А. Введение в эмбриологию покрытосеменных (учебно-методическое пособие). М.: Изд-во МГУ. 1991. 79 с.
15. Челомбитько А.С. Биология: учебник для Вузов.
16. Эмбриология растений / Под ред. И.П. Ермакова (пер. с англ. Embryology of Angiosperms, ed. by B.M. Johri, 1984). М: Агропромиздат. 1990. Т. 1. 496 с., т. 2. 427 с.
Приложение 1

Приложение 2
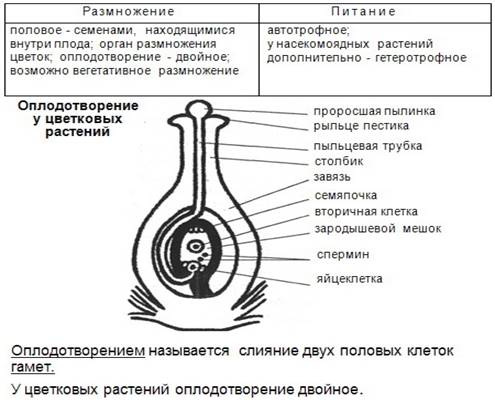
Рисунок. Отделпокрытосеменные(цветковые)