| Скачать .docx | Скачать .pdf |
Реферат: Термодинамика в существовании биологических систем
Термодинамика в существовании биологических систем.
Содержание
ВВЕДЕНИЕ
ТЕРМОДИНАМИКА БИОЛОГИЧЕСКИХ ПРОЦЕССОВ
АНАЛИЗ ТЕРМОДИНАМИЧЕСКИХ ПРОЦЕССОВ, ПРОИСХОДЯЩИХ В ЖИВЫХ ОРГАНИЗМАХ.
ТЕРМОДИНАМИКА НЕОБРАТИМЫХ ПРОЦЕССОВ В БИОЛОГИЧЕСКИХ СИСТЕМАХ ВБЛИЗИ РАВНОВЕСИЯ
ИЗМЕНЕНИЕ ЭНТРОПИИ В ОТКРЫТЫХ СИСТЕМАХ
СООТНОШЕНИЕ ОНЗАГЕРА
ТЕОРЕМА ПРИГОЖИНА
ТЕРМОДИНАМИКА АКТИВНОГО ТРАНСПОРТА
ТЕРМОДИНАМИКА СИСТЕМ ВДАЛИ ОТ РАВНОВЕСИЯ
ЭНТРОПИЯ И БИОЛОГИЧЕСКАЯ ИНФОРМАЦИЯ
СПИСОК ЛИТЕРАТУРЫ
Итак, жизнь есть вихрь, то более быстрый, то более медленный, более сложный или менее сложный, увлекающий в одном и том же направлении одинаковые молекулы. Но каждая отдельная молекула вступает в него и покидает его, и это длится непрерывно, так что форма живого существа более существенна, чем материал .
Жорж Кювье
ВВЕДЕНИЕ
Биология как наука является одной из старейших в мире.
Уже в Ветхом завете, насчитывающем более 3000 лет, имеются конкретные биологические сведения о растительном и животном мире, даны рекомендации о режиме питания, применении в пищу тех или иных видов растительности. До нашего времени дошли труды многих великих ученых древности: Аристотеля, Авиценны и других, внесших огромный вклад в понимание биологических процессов. Тем не менее, за всю многовековую историю биологии из-за исключительной сложности изучаемого предмета ученым так и не удалось полностью раскрыть секреты механизмов и движущих сил, обеспечивающих жизнеспособность живых организмов.
Достаточно сказать, что даже само определение жизни до настоящего времени не однозначно. В связи с этим за все время своего становления, вплоть до начала 20-го века, биология как наука опиралась на различные гипотезы и теории, которые периодически опровергали друг друга по мере ее развития и получения новых научных данных.
Принципиально новым шагом в биологии стала работа Эрвина Симоновича Бауэра «Теоретическая биология», опубликованная в 1935 году.
В этой работе им впервые было установлено, что живые организмы всегда находятся в состоянии термодинамического неравновесия, что противоречило популярной в то время теории термодинамического равновесия (первому и второму началам термодинамики).
Э. Бауэром был сформулирован “Всеобщий закон биологии” в следующей редакции:
“Все и только живые системы никогда не бывают в равновесии и исполняют за счет своей свободной энергии постоянную работу против равновесия, требуемого законами физики и химии при существующих внешних условиях”.
Э. Бауэром также был сформулирован “Принцип устойчивого неравновесия живых систем”:
“Для живых систем характерно именно то, что они за счет своей свободной энергии производят работу против ожидаемого равновесия”.
Практически одновременно с Э. Бауэром, в 1931 г. Ларс Онсагер открыл первые общие соотношения (соотношения взаимности) неравновесной термодинамики в линейной, слабо неравновесной области. Соотношения взаимности Онсагера были первым значительным результатом в неравновесной термодинамике – термодинамике необратимых процессов.
В дальнейшем основные работы в области теории неравновесной термодинамики были выполнены бельгийским физиком и физикохимиком, лауреатом Нобелевской премии 1977г. Ильей Романовичем Пригожиным.
Как утверждает И. Пригожин: “…и биосфера в целом, и ее различные компоненты, живые или неживые, существуют в сильно неравновесных условиях. В этом смысле жизнь, заведомо укладывающаяся в рамки естественного порядка, предстает перед нами как высшее проявление происходящих в природе процессов самоорганизации ”.
Тем не менее, до настоящего времени, несмотря на огромный накопленный материал, биология является в основном наукой описательной. Получение той или иной биологической информации часто достигается статистически, путем огромного количества экспериментов и наблюдений. В результате этого целый ряд важнейших научных данных до сих пор является недоступным. В качестве примера можно привести проблему количественной оценки сочетанного воздействия на живые организмы нескольких различных физических и химических нагрузок.
Поэтому одними из важнейших задач современной биологии являются определения законов, описывающих физически и математически принципы и процессы функционирования живых организмов.
ТЕРМОДИНАМИКА БИОЛОГИЧЕСКИХ ПРОЦЕССОВ
Термодинамика рассматривает общие закономерности превращения энергии в форме тепла и работы между телами. В открытых биологических системах постоянно происходит процесс обмена энергией с внешней средой. Внутренние метаболические процессы также сопровождаются превращениями одних форм энергии в другие. Достаточно напомнить о механических процессах, трансформации энергии кванта света в энергию электронного возбуждения молекул пигментов, а затем в энергию химических связей восстановленных соединений в фотосинтезе. Другой пример - преобразование энергии электрохимического трансмембранного потенциала в энергию АТФ в биологических мембранах.
Механизмы трансформации энергии в биоструктурах связаны с конформационными превращениями особых макромолекулярных комплексов, таких, как реакционные центры фотосинтеза, Н-АТФаза хлоропластов и митохондрий, бактериородопсин. Однако помимо выяснения детального характера происходящих здесь процессов особый интерес представляют общие характеристики эффективности преобразования энергии в таких макромолекулярных машинах.
На эти вопросы призвана ответить термодинамика биологических процессов - один из разделов теоретической биофизики.
АНАЛИЗ ТЕРМОДИНАМИЧЕСКИХ ПРОЦЕССОВ, ПРОИСХОДЯЩИХ В ЖИВЫХ ОРГАНИЗМАХ.
Как известно из биологии, получение свободной энергии из продуктов питания и ее потребление для обеспечения жизнедеятельности живыми организмами осуществляется с помощью метаболизма – циклов обмена веществ, непрерывно происходящих в их клетках, и представляющих собой комплексы разнообразных биохимических реакций расщепления и синтеза веществ различными метаболическими путями.
Поскольку обмен веществ происходит циклами, то в клетках, в соответствии с этими циклами, происходят непрерывные периодические изменения концентраций веществ, участвующих в многочисленных биохимических реакциях. На рис.1. в качестве примера представлен график внутриклеточных колебаний концентрации кальция.
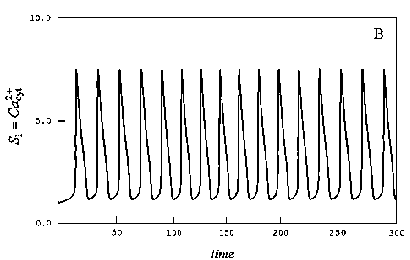
Рис.1. График внутриклеточных колебаний кальция..
Как видно из рис.1, внутриклеточные колебания кальция представляют собой непрерывный периодический процесс.
Среди всех биохимических реакций особую роль играют реакции синтеза из молекул углеводов и жиров, содержащихся в питательных веществах, аденозинтрифосфата (АТФ), и его последующего расщепления, в результате чего выделяется энергия. Структура АТФ показана на рис.2.
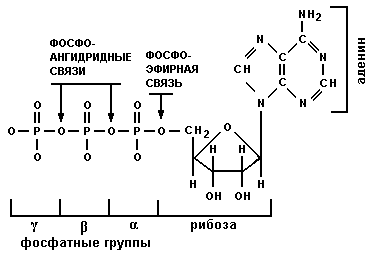
Рис.2. Структура АТФ
Как видно из рис.2, АТФ состоит из трех фосфатных групп, остатков азотистого основания аденина) и остатка сахара (рибозы). При разрыве фосфоангидридных и фосфоэфирной связей выделяется энергия.
Фосфатные группы могут быть постадийно отщеплены путем растворения в воде гидролиза) и образования ортофосфата или неорганического фосфата и аденозиндифосфата АДФ, а затем, после расщепления АДФ, и аденозинмонофосфата с выделением энергии на каждой стадии:
АТФ + Н2 О→ АДФ + Фн + H+ + ΔG = -30 кДж/моль
АДФ + Н2 О→АМФ + Фн + H+ + ΔG = -30 кДж/моль
АМФ + Н2 О→аденозин + Фн + H+ + ΔG = -13 кДж/моль
Здесь: Фн - неорганический фосфат;
H+ - положительный ион водорода;
ΔG - изменение свободной энергии, выделяемой при отрыве концевой фосфатной группы.
Обычно клетки извлекают энергию из АТФ во время клеточного дыхания, отщепляя от его молекулы только одну фосфатную группу.
На рис.3 приведен пример упрощенного графика циклов синтеза-расщепления АТФ.
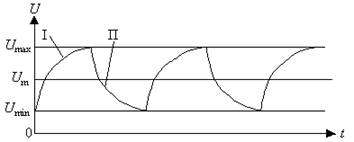
Рис.3. Упрощенный график синтеза-расщепления АТФ.
U - количество АТФ в клетке; I – процесс синтеза; П – процесс расщепления.
Как видно из графика на рис.3, изображенные на нем процессы синтеза АТФ (кривая I), и расщепления АТФ (кривая II) по своей форме близки к экспонентам.
Верхние и нижние границы концентрации биохимических веществ U max и U min определяются соответствующими положительной и отрицательной обратными связями, имеющимися в живых системах.
Скорость протекания биохимических реакций регулируется соответствующими биохимическими катализаторами, ускоряющими эти реакции, и ингибиторами, замедляющими их, как это видно из рис.4, где представлен пример схемы работы механизма регуляции ферментативных реакций.
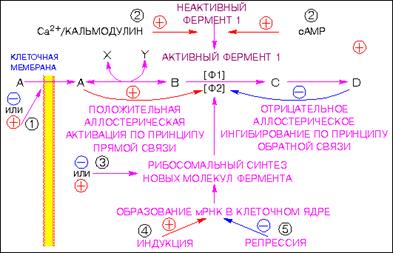
Рис.4. Пример схемы регуляции ферментативных реакций
Цифры, заключенные в кружки, указывают вероятные участки действия гормонов.
1 - изменение проницаемости мембраны;
2 - переход фермента из неактивной формы в активную;
3 - изменение скорости трансляции мРНК на рибосомальном уровне;
4 - индукция образования новой мРНК;
5 - репрессия образования мРНК.
Аналогично происходят групповые когерентные биохимические реакции на уровне органов, систем и организмов в целом.
На рис.6 представлены графики чередования синтеза и расщепления АТФ и чередования затрат и выделения энергии.
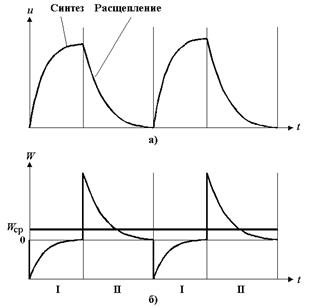
Рис.6.
а ) График чередования фаз синтеза и расщепления веществ;
б ) График чередования фаз потребления и выделения энергии;
I – фаза потребления энергии;
II - фаза выделения энергии.
W п – среднедействующее значение потребляемой энергии при синтезе АТФ;
W в – среднедействующее значение выделяемой энергии при расщеплении АТФ;
W ср – среднедействующее результирующее значение выделяемой энергии;
Как видно из рис.6, в результате последовательных циклов биохимических реакций синтеза и расщепления АТФ, при которых происходят термодинамические колебания, выделяется энергия W ср >0.
Значение энергии W ср определяется реакцией организма на внутренние воздействия и воздействия внешней среды путем ферментативной регуляции процессов синтеза и расщепления АТФ.
Таким образом, эти термодинамические колебания потребления и выделения энергии и являются механизмом обеспечения устойчивого неравновесного термодинамического состояния.
Величина устойчивости неравновесного термодинамического состояния определяется параметрами механизмов регуляции ферментативных реакций и биологическими свойствами организма.
Датчиками, определяющими скорость и характер метаболических процессов, в живых организмах являются аллостерические модуляторы и гормоны, непрерывно контролирующие термодинамическое состояние организма.
Устойчивость неравновесного термодинамического состояния биологических систем обеспечивается следующим образом:
- при минимальном значении неравновесного термодинамического состояния клеток, органов и целостного организма, например температуры, датчики включают режим расщепления АТФ, в результате чего энергетика организма (его температура) начинает возрастать, достигая некоторого максимального значения;
- при максимальном значении неравновесного термодинамического состояния клеток, органов и целостного организма (температуре) датчики включают режим синтеза АТФ, при котором энергетика организма (температура) начинает уменьшаться.
Отсюда видно, что неравновесное термодинамическое состояния организма всегда непрерывно колеблется в определенных пределах, обеспечивая тем самым устойчивость этого состояния.
В этом также нетрудно убедиться на примере частично управляемой физиологической функции организма – дыхании, при котором в одной фазе – вдохе - обеспечивается снабжение организма кислородом, в при другой фазе – выдохе – удаляется углекислый газ. Процессы вдоха и выдоха непрерывно регулируются. В спокойном состоянии глубина дыхания незначительная, а его частота – низкая. Однако при интенсивной работе значительно возрастает как глубина дыхания, так и его частота. Отсюда видно, что в зависимости от нагрузки эта физиологическая функция, совместно с другими, оперативно изменяет свои параметры для сохранения устойчивого неравновесного термодинамического состояния организма при изменившихся условиях.
Из этого следует, что кроме “Всеобщего закона биологии” Э. Бауэра биологические системы подчиняются также закону Доброборского, излагаемому в следующей редакции:
Закон Доброборского
Устойчивость неравновесного термодинамического состояния биологических систем обеспечивается непрерывным чередованием фаз потребления и выделения энергии посредством управляемых реакций синтеза и расщепления АТФ
Из этого закона вытекают следующие следствия:
1. В живых организмах ни один процесс не может происходить непрерывно, а должен чередоваться с противоположно направленным: вдох с выдохом, работа с отдыхом, бодрствование со сном, синтез с расщеплением и т.д.
2. Состояние живого организма никогда не бывает статическим, а все его физиологические и энергетические параметры всегда находятся в состоянии непрерывных колебаний относительно средних значений как по частоте, так и по амплитуде.
Эти колебания являются биоритмами.
Таким образом, живые организмы с помощью биоритмов обеспечивают устойчивость своего неравновесного термодинамического состояния. Отсюда можно считать, что биоритмы являются способом существования всех живых организмов.
ТЕРМОДИНАМИКА НЕОБРАТИМЫХ ПРОЦЕССОВ В БИОЛОГИЧЕСКИХ СИСТЕМАХ ВБЛИЗИ РАВНОВЕСИЯ
В классической термодинамике рассматриваются главным образом равновесные состояния системы, в которых параметры не изменяются во времени. Однако в открытых системах реакции и соответствующие энергетические превращения происходят постоянно, и поэтому здесь необходимо знать скорости трансформации энергии в каждый момент времени. Это значит, что в энергетических расчетах надо учитывать и фактор времени. Для этого необходимо каким-то образом сочетать термодинамический и кинетический подходы в описании свойств открытой системы.
Напомним вначале содержание основных законов классической термодинамики и результаты их применения в биологии. Согласно первому закону, количество теплоты δQ , поглощенное системой из внешней среды, идет на увеличение ее внутренней энергии dU и совершение общей работы δA , которая включает работу против сил внешнего давления P по изменению объема dV системы и максимальную полезную работу δAmax , сопровождающую химические превращения:
δQ = dU + δA,
где работа
δА = pdV + δAmax
или
δQ = dU + pdV + δAmax . (1)
Опытная проверка первого закона проводилась в специальных калориметрах, где измерялась теплота, выделенная организмом в процессах метаболизма, при испарениях, а также вместе с продуктами выделения. Оказалось, что выделенная организмом теплота полностью соответствует энергии, поглощенной вместе с питательными веществами. Справедливость первого закона означает, что сам по себе организм не является независимым источником какой-либо новой энергии.
Второй закон термодинамики дает критерий направленности самопроизвольных необратимых процессов. Всякое изменение состояния системы описывается соответствующим изменением особой функции состояния - энтропии S , которая определяется суммарной величиной поглощенных системой приведенных теплот Q / T :
![]()
Знак неравенства относится к неравновесным процессам. В изолированных системах dQ = 0 и, следовательно,
dS ≥ 0. (3)
В этом и состоит эволюционный критерий направленности необратимых изменений в изолированных системах, которые всегда идут с увеличением энтропии до ее максимальных значений при окончании процесса и установлении термодинамического равновесия. Увеличение энтропии означает падение степени упорядоченности и организованности в системе, ее хаотизацию.
ИЗМЕНЕНИЕ ЭНТРОПИИ В ОТКРЫТЫХ СИСТЕМАХ
Применение второго закона к биологическим системам в его классической формулировке приводит, как кажется на первый взгляд, к парадоксальному выводу, что процессы жизнедеятельности идут с нарушением принципов термодинамики.
В самом деле, усложнение и увеличение упорядоченности организмов в период их роста сопровождаются кажущимся уменьшением, а не увеличением энтропии, как должно было бы следовать из второго закона.
Однако увеличение энтропии в необратимых самопроизвольных процессах происходит в изолированных системах, а биологические системы являются открытыми. Проблема поэтому заключается в том, чтобы, во-первых, понять, как связано изменение энтропии с параметрами процессов в открытой системе, а во-вторых, выяснить, можно ли предсказать общее направление необратимых процессов в открытой системе по изменению ее энтропии. Главная трудность в решении этой проблемы состоит в том, что мы должны учитывать изменение всех термодинамических величин во времени непосредственно в ходе процессов в открытой системе. Постулат И.Р. Пригожина состоит в том, что общее изменение энтропии dS открытой системы может происходить независимо либо за счет процессов обмена с внешней средой (de S ), либо вследствие внутренних необратимых процессов (di S ):
dS = de S + di S. (4)
Во всех реальных случаях di S > 0 , и только если внутренние процессы идут обратимо и равновесно, то diS = 0 . Для изолированных систем de S = 0 , и мы приходим к классической формулировке второго закона:
dS = di S = 0.
В клеточном метаболизме всегда можно выделить такие две группы процессов. Например, поступление извне глюкозы, выделение наружу продуктов ее окисления (de S ) и окисление глюкозы в процессах дыхания (di S ).
В фотосинтезе приток свободной энергии света приводит к уменьшению энтропии клетки de S < 0 , а процессы дыхания, диссимиляции в клетке увеличивают ее энтропию di S > 0 . В зависимости от соотношения скоростей изменения de S и di S общая энтропия dS открытой системы может либо увеличиваться, либо уменьшаться со временем.
Если единственной причиной необратимости и увеличения энтропии системы являются ее внутренние процессы, то они ведут к уменьшению ее термодинамического потенциала. В этом случае
![]()
где G - полный термодинамический потенциал (или энергия Гиббса G = U + PV ̶̶ TS ).
Можно показать, что скорость возникновения положительной энтропии внутри открытой химической системы зависит от химического сродства А и скорости реакции ν:
![]()
Химическое сродство А определяется разностью химических потенциалов реагентов реакции, то есть ее движущей силой.
Выражение (6) имеет простой смысл. Оно показывает, что скорость образования в системе положительной энтропии в ходе необратимого химического процесса прямо пропорциональна его движущей силе A и скорости ν . Очевидно, что величина di S / dt является, вообще говоря, переменной, поскольку в ходе химической реакции все время изменяются переменные концентрации реагирующих веществ, а следовательно, и зависящие от них величины А и ν .
СООТНОШЕНИЕ ОНЗАГЕРА
Между движущими силами и скоростями (потоками) должна, очевидно, существовать взаимосвязь, при которой увеличение (уменьшение) движущей силы вызывает соответствующее увеличение (уменьшение) скорости процесса. Это относится не только к химическим реакциям, но и к другим необратимым процессам.
Например, процессы переноса тепла и диффузии вещества через мембрану из одной фазы в другую включают движущие силы - градиенты температуры и концентраций, а потоки соответствуют переносу тепла или вещества между двумя фазами. Во всех этих случаях возрастание энтропии имеет вид
![]()
где Х - движущая сила, J - величина потока.
Если система находится вблизи равновесия, где величины движущих сил и потоков очень малы, то между ними имеется прямая пропорциональная зависимость:
J = LX, (8)
где L - постоянный линейный коэффициент.
Если в открытой системе вблизи равновесия протекают одновременно несколько процессов, то между ними существуют термодинамические соотношения, отражающие их взаимное влияние. Для двух процессов (J1 , X1 ) и (J2 , X2 ) эти соотношения имеют вид
J1 = L11 X1 + L12 X2 , (9)
J2 = L21 X1 + L22 X2 ,
где постоянные коэффициенты L11 , L22 отражают зависимость потока от своей силы, а коэффициенты L12 , L21 соответствуют взаимному влиянию силы одного процесса на поток другого процесса. Они носят название коэффициентов взаимности Онзагера. Вблизи равновесия
L12 = L21 . (10)
Теперь можно установить количественную связь между одновременно протекающими в клетке процессами, не зная их молекулярных механизмов.
Рассмотрим процесс активного переноса вещества через биологическую мембрану, который происходит за счет энергии сопрягающего метаболического процесса и поэтому может идти против градиента концентрации переносимого вещества. Тогда
J1 = L11 X1 + L12 X2 ,
J2 = L21 X1 + L22 X2 , L12 = L21 ,
где процесс (J1 , X1 ) сопряженного переноса идет против градиента силы X1 (J1 , X1 < 0 ) за счет энергии сопрягающего процесса (J2 , X2 > 0 ). Если сопряжение отсутствует, то L12 = L21 = 0 и процессы идут независимо друг от друга под действием только своих движущих сил
J1 = L11 X1 , J2 = L22 X2 .
В начальные моменты запуска системы большая скорость сопрягающего процесса J2
снижается до минимальных значений, одновременно растет величина X1
. В результате этих изменений устанавливается стационарное состояние, когда результирующий сопряженный поток обращается в нуль: J1
= 0
. Если система полностью сопряжена, то и для сопрягающего потока устанавливается стационарное состояние J2
= 0
. В этом случае в системе нет видимых изменений и вся энергия сопрягающего потока тратится на поддержание силы X1
. Можно мысленно представить себе колесо турбины в воде (X1
), скорость ее движения (J1
) и потока воды (J2
). Эти соображения справедливы не только для активного переноса, но и для других случаев. Так, в митохондриях скорость окисления субстрата, то есть скорость движения (J2
), связана с отношением АДФ / АТФ, то есть движущей силой X1
. В состоянии митохондрий, когда концентрация АДФ равна нулю и видимого образования АТФ не происходит (J1
= 0
), вся энергия тратится на поддержание максимального уровня ![]() . Добавление разобщителей уменьшает величину X1
, но тогда уже J1
≠ 0
, что приводит к ускорению сопрягающего потока.
. Добавление разобщителей уменьшает величину X1
, но тогда уже J1
≠ 0
, что приводит к ускорению сопрягающего потока.
Коэффициент трансформации энергии в сопрягающих процессах равен | J1 X1 | / J2 X2 и в митохондриях может достигать значений 80-90%. Применение уравнений Онзагера позволяет получить характеристики макромолекулярных комплексов - биологических трансформаторов энергии, не прибегая к детальному анализу механизмов их функционирования.
ТЕОРЕМА ПРИГОЖИНА
Мы уже видели, что в стационарном состоянии в открытой системе
![]()
причем каждый из членов de S / dt и di S / dt отличен от нуля. Возникает вопрос, можно ли по характеру изменений величины di S / dt во времени предсказать установление в открытой системе стационарного состояния. Ответ на этот вопрос дает теорема Пригожина, согласно которой в стационарном состоянии положительная функция di S / dt принимает минимальное положительное значение. Следовательно, по мере приближения к стационарному состоянию скорость образования энтропии внутри открытой системы монотонно уменьшается, постепенно приближаясь к своему минимальному положительному значению. В этом состоит критерий направленности необратимых процессов в открытых системах, находящихся вблизи равновесия, где справедливы соотношения Онзагера.
Из монотонного характера изменения Tdi S / dt следует, что вблизи равновесия стационарное состояние не может представлять собой автоколебательный режим. Действительно, в этом случае переменные концентрации в системе (а следовательно, величины J и X ) изменяются периодически, что несовместимо с однонаправленным монотонным изменением Tdi S / dt и ее постоянством в стационарной точке. Экспериментальные измерения скорости образования энтропии внутри системы можно проводить в калориметрах, изучая тепловые потоки, сопровождающие образование энтропии при необратимых изменениях в системе. В опытах на биологических объектах было показано, например, что скорость теплопродукции и интенсивность дыхания в процессе развития зародышей непрерывно уменьшаются начиная с первых стадий развития организма и достигают постоянных значений в стационарной фазе роста. Следует, однако, иметь в виду, что уровень термогенеза может меняться в ходе развития организма не только вследствие изменения величин движущих сил и потоков. Теплопродукция организма зависит и от состояния мембранных структур и степени сопряжения процессов окислительного фосфорилирования. Наконец, принципиальным является то, что биологические системы, вообще говоря, находятся вдали от равновесия, где пропорциональность между J и X (9) или соотношения взаимности (10) нарушаются. Это особенно важно для биохимических процессов, где наиболее характерны переходы с изменениями ∆G = = 1-2 ккал/мол, а линейные соотношения Онзагера справедливы при G ≤ 0,2 ккал/моль. В таких условиях в стационарных состояниях, далеких от равновесия, теорема Пригожина несправедлива (автоколебательный режим).
ТЕРМОДИНАМИКА АКТИВНОГО ТРАНСПОРТА
Мы рассмотрим систему активного транспорта одного иона (натрия), не сопряженного с переносом других веществ. Для простоты рассуждения будем считать, что можно выделить один метаболический процесс, приводящий в движение активный транспорт натрия.
Обозначим скорость активного транспорта катиона через ![]() , скорость метаболизма через Jr
, тогда
, скорость метаболизма через Jr
, тогда
![]()
![]()
где Х+ - отрицательная разность электрохимических потенциалов катиона (идущего против градиента "своей" движущей силы), А - сродство метаболической реакции, обеспечивающей транспорт. В случае одной метаболической реакции (гидролиз АТФ) скорости потребления и производства всех метаболитов связаны стехиометрически. Поэтому для оценки скорости метаболизма можно взять, например, скорость потребления О2 в дыхании. Тогда сродство А может быть выражено как отрицательное изменение полного термодинамического потенциала метаболической реакции (гидролиз АТФ) на моль потребления О2 .
Помещая с обеих сторон мембраны одинаковые растворы ![]() и изменяя трансмембранную разность потенциалов ∆φ можно найти из (11) феноменологические коэффициенты
и изменяя трансмембранную разность потенциалов ∆φ можно найти из (11) феноменологические коэффициенты  ,
, ![]() , которые определяются из наклона соответствующих прямых ∆
, которые определяются из наклона соответствующих прямых ∆![]() и
и ![]()
Очевидно, в экспериментальных условиях для нахождения коэффициентов важно сохранить параметры системы и уметь направленно варьировать Na при постоянстве А . Опыты проводили на коже лягушки, где варьировали значения XNa путем изменения ∆φ. При этом сохранялся состав смывающей жидкости и концентрация натрия поддерживалась неизменной.
В прямых экспериментах была подтверждена линейная зависимость скорости активного транспорта ![]() от ∆φ на коже лягушки, где ∆φ изменяли симметрично в области 0
от ∆φ на коже лягушки, где ∆φ изменяли симметрично в области 0 ![]() 80 мВ. Величину Jr
можно определять по поглощению О2
с применением кислородных электродов. Оказалось, что при симметричных возмущениях потенциала соотношение между Jr
и ∆φ было линейным в интервале 0
80 мВ. Величину Jr
можно определять по поглощению О2
с применением кислородных электродов. Оказалось, что при симметричных возмущениях потенциала соотношение между Jr
и ∆φ было линейным в интервале 0 ![]() 70 мВ. Была изучена также зависимость и J2
от наружной концентрации натрия в условиях постоянства его внутренней концентрации при постоянной нулевой разности электрических потенциалов (∆φ=0). В этих условиях также наблюдалась линейная зависимость скорости активного транспорта JNa
и поглощения кислорода Jr
от разности химических потенциалов ∆μNa
на мембране. Однако если ХNa
изменять путем варьирования внутренней концентрации натрия, то линейность уже не соблюдается. Это обусловлено уже изменениями в микроструктуре и составе самой мембраны.
70 мВ. Была изучена также зависимость и J2
от наружной концентрации натрия в условиях постоянства его внутренней концентрации при постоянной нулевой разности электрических потенциалов (∆φ=0). В этих условиях также наблюдалась линейная зависимость скорости активного транспорта JNa
и поглощения кислорода Jr
от разности химических потенциалов ∆μNa
на мембране. Однако если ХNa
изменять путем варьирования внутренней концентрации натрия, то линейность уже не соблюдается. Это обусловлено уже изменениями в микроструктуре и составе самой мембраны.
Подобные исследования были успешно проведены и в отношении активного транспорта протонов с применением уравнений неравновесной термодинамики для двух потоков. Во всех случаях варьирование Х+ позволяет оценить феноменологические коэффициенты и сродство А движущей метаболической реакции. В последнее время делаются успешные попытки применить подобный формализм для описания процессов фосфорилирования в митохондриях и хлоропластах. Считается общепринятым, что в этих объектах имеется тесное сопряжение между тремя главными процессами, лежащими в основе биоэнергетики клеточных мембран: электронный транспорт с окислением субстрата (J0 , А0 ), фосфорилирование АДФ с образованием АТФ (JР , АР ), транслокация протонов через сопрягающую мембрану (JH , ∆μH ). Ключевую роль играет трансмембранная циркуляция протонов, которая индуцируется переносом электронов и, в свою очередь, запускает синтез АТФ. Как оказалось, здесь также имеет место линейная зависимость между силами и потоками, что должно позволить найти опытным путем коэффициенты L .
На основе такого подхода обсуждаются различные гипотезы энергетического сопряжения. В частности, хемиосмотическая гипотеза предполагает непосредственную связь с образованием АТФ только транслокации протонов, но не переноса электронов. В этом предельном случае коэффициент LP0 должен быть равен нулю. Работы в этом направлении продолжаются. Надо, конечно, ясно понимать, что термодинамический анализ может помочь оценить энергетическую эффективность и степень сопряжения процессов, но ничего не говорит об их молекулярных механизмах.
ТЕРМОДИНАМИКА СИСТЕМ ВДАЛИ ОТ РАВНОВЕСИЯ
Определить возможность самопроизвольного перехода изолированной системы между двумя состояниями можно методами классической термодинамики, сравнивая значение энтропии этих состояний. В открытой системе возникают стационарные состояния, которые могут находиться далеко от термодинамического равновесия. Вопрос о возможности перехода открытой системы из некоторого начального в конечное стационарное состояние можно решить на основе теоремы Пригожина, если оба состояния лежат вблизи термодинамического равновесия. Однако вдали от равновесия уже нельзя сделать однозначных выводов о том, как меняется скорость образования энтропии при приближении к стационарному состоянию. Эволюция таких неравновесных динамических систем определяется прежде всего кинетикой взаимодействия составных элементов, а не статистической упорядоченностью начального и конечного состояния системы согласно классической термодинамике. Такие системы имеют ограниченное число конечных состояний и ведут себя наподобие химических машин. Поэтому распространение идей термодинамики на неравновесные системы может дать лишь дополнительную характеристику далеких от равновесия стационарных состояний, положение и пути достижения которых определяются кинетическими уравнениями.
По мере удаления от равновесия будут расти величины X и J и система может удалиться от равновесия и покинуть область линейной термодинамики, не теряя общей устойчивости. Возможно, однако, что при удалении от равновесия в системе наступает бифуркационное изменение и возникает неустойчивость. Возникает, как говорят, термодинамическая флуктуация, уводящая систему от неустойчивой точки, что может стать причиной распада системы. Однако при определенных значениях параметров эта флуктуация как бы дает толчок, переводящий систему к новому состоянию, которому и передается устойчивость. Например, появлению предельного цикла, возникновению диссипативных структур в распределенных системах также предшествует нарушение термодинамической устойчивости вдали от равновесия. Наконец, переходы между устойчивыми стационарными состояниями происходят на границе устойчивости, когда система совершает скачкообразный переход между ними.
Таким образом, термодинамические признаки устойчивости стационарных состояний совпадают с соответствующими математическими признаками и могут служить их дополнительной характеристикой. Однако вдали от равновесия не существует общих термодинамических критериев направления движения открытой системы, поскольку ее поведение определяется динамическими свойствами и механизмами регуляции, а не общими статистическими закономерностями, как во втором законе классической термодинамики. Эта особенность обусловливает также и сложность применения понятий энтропии и информации при описании общих свойств биологических систем.
ЭНТРОПИЯ И БИОЛОГИЧЕСКАЯ ИНФОРМАЦИЯ
Согласно формуле Больцмана, энтропия определяется как логарифм числа микросостояний, возможных в данной макроскопической системе:
S = kБ ln W, (12)
где kБ = 1,38 · 10-16 эрг/град, или 3,31·10- 24 энтропийных единиц (1 э.е.= 1 кал/град = 4,1 Дж/К), или 1,38 · 10- 23 Дж/К - постоянная Больцмана, W - число микросостояний (например, число способов, которыми можно разместить молекулы газа в сосуде). Именно в этом смысле энтропия есть мера неупорядоченности и хаотизации системы. В реальных системах существуют устойчивые и неустойчивые степени свободы. Им соответствуют, например, твердые стенки сосуда и молекулы заключенного в нем газа.
Понятие энтропии связано именно с неустойчивыми степенями, по которым возможна хаотизация системы, а число возможных микросостояний намного больше единицы. В полностью устойчивых системах реализуется только одно-единственное решение, то есть число способов, которыми осуществляется это единственное макросостояние системы, равно единице (W = 1), а следовательно, энтропия равна нулю. В биологии использовать понятие "энтропия", а следовательно, и термодинамические представления можно только по отношению к конкретным метаболическим процессам, а не для описания в целом поведения и общебиологических свойств организмов. Связь энтропии и информации в теории информации была установлена для статистических степеней свободы. Допустим, что мы получили информацию о том, каким конкретно способом из всех возможных способов осуществлено данное макросостояние системы. Очевидно, количество информации, которое мы при этом получали, будет тем больше, чем больше была исходная неопределенность, или энтропия, системы.
Напомним, что, согласно теории информации, в этом случае количество информации о единственном реальном состоянии системы
I = log2 W. (13)
За единицу количества информации (бит) принимается информация, содержащаяся в достоверном сообщении, когда число исходных возможных состояний было равно W = 2:
I = log2 W = 1 бит. (14)
Например, сообщение о том, на какую сторону упала монета при бросании в воздух, содержит количество информации в 1 бит. Сопоставляя формулы (12) и (13) можно найти связь между энтропией (в энтропийных единицах) и информацией (в битах)
S (э.е.) = 2,3 · 10- 24 . (15)
Теперь попытаемся формально оценить количество информации, содержащейся в теле человека, где имеется 1013 клеток. С помощью формулы (12) получим величину
I = log2 (1013 !) ̶ 1013 log2 1013 ̶ 4 · 1014 бит.
Такое количество информации необходимо было бы исходно получить, чтобы осуществить единственно правильное расположение клеток в организме. Этому эквивалентно весьма незначительное снижение энтропии системы на
∆S = 2,3 · 10- 24 ·4 · 1014 ̶ 10- 9 э.е. ̶ 4 · 10- 9 Дж/К.
Если считать, что в организме осуществляется также уникальный характер расположения аминокислотных остатков в белках и нуклеиновых остатков в ДНК, то общее количество информации, содержащейся в теле человека, составит
I = 1,3 · 1026 бит,
что эквивалентно небольшому понижению энтропии на ∆S
![]() 300 э.е. = 1200 Дж/К. В процессах метаболизма это снижение энтропии легко компенсируется увеличением энтропии при окислении 900 молекул глюкозы. Таким образом, формально сопоставление формул (12) и (13) показывает, что биологические системы не обладают какой-либо повышенной информационной емкостью по сравнению с другими неживыми системами, состоящими из того же числа структурных элементов. Этот вывод на первый взгляд противоречит роли и значению информационных процессов в биологии.
300 э.е. = 1200 Дж/К. В процессах метаболизма это снижение энтропии легко компенсируется увеличением энтропии при окислении 900 молекул глюкозы. Таким образом, формально сопоставление формул (12) и (13) показывает, что биологические системы не обладают какой-либо повышенной информационной емкостью по сравнению с другими неживыми системами, состоящими из того же числа структурных элементов. Этот вывод на первый взгляд противоречит роли и значению информационных процессов в биологии.
Однако связь между I и S в (15) справедлива лишь по отношению к информации о том, какое из всех W микросостояний реализовано в данный момент. Эта микроинформация, связанная с расположением всех атомов в системе, на самом деле не может быть запомнена и сохранена, поскольку любое из таких микросостояний быстро перейдет в другое из-за тепловых флуктуаций. А ценность биологической информации определяется не количеством, а прежде всего возможностью ее запоминания, хранения, переработки и дальнейшей передачи для использования в жизнедеятельности организма.
Основное условие восприятия и запоминания информации - способность рецепторной системы переходить вследствие полученной информации в одно из устойчивых состояний, заранее заданных в силу ее организации. Поэтому информационные процессы в организованных системах связаны только с определенными степенями свободы. Сам процесс запоминания информации должен сопровождаться некоторой потерей энергии в рецепторной системе для того, чтобы она могла в ней сохраниться достаточное время и не теряться вследствие тепловых флуктуаций. Именно здесь и осуществляется превращение микроинформации, которую система не могла запомнить, в макроинформацию, которую система запоминает, хранит и затем может передать другим акцепторным системам. Как говорят, энтропия есть мера множества незапоминаемых системой микросостояний, а макроинформация - мера множества их состояний, о пребывании в которых система должна помнить.
Информационная емкость в ДНК, например, определяется не только количеством определенных нуклеотидов, а общим числом микросостояний, включающих колебания всех атомов цепочки ДНК. Процесс запоминания информации в ДНК - это фиксация определенного расположения нуклеотидов, которое устойчиво вследствие образующихся химических связей в цепочке. Дальнейшая передача генетической информации осуществляется в результате биохимических процессов, в которых диссипация энергии и образование соответствующих химических устойчивых структур обеспечивают эффективность биологической переработки информации. В целом информационные процессы широко распространены в биологии. На молекулярном уровне они протекают не только при запоминании и переработке генетической информации, но и при взаимном узнавании макромолекул, обеспечивают специфичность и направленный характер ферментативных реакций, имеют важное значение при взаимодействии клеточных мембран и поверхностей. Физиологические рецепторные процессы, играющие самостоятельную информационную роль в жизнедеятельности организма, также основаны на взаимодействиях макромолекул. Во всех случаях макроинформация возникает исходно в виде конформационных изменений при диссипации части энергии по определенным степеням свободы во взаимодействующих макромолекулах. В результате макроинформация оказывается записанной в виде набора достаточно энергетически глубоких конформационных подсостояний, которые позволяют сохранять эту информацию в течение времени, необходимого для ее дальнейшей переработки. Биологический смысл этой макроинформации реализуется уже в соответствии с особенностями организации биологической системы и конкретными клеточными структурами, на которых разыгрываются дальнейшие процессы, приводящие в итоге к соответствующим физиолого-биохимическим эффектам.
ЗАКЛЮЧЕНИЕ
Приведенные материалы показали, что примененный подход к решению ряда проблем теоретической биологии с позиций неравновесной термодинамики позволил в определенной степени объяснить сущность и закономерность функционирования живых организмов и их способностей к адаптации. Изложенные в работе новые законы и положения позволяют с достаточной уверенностью утверждать, что движущей силой в процессах жизнедеятельности живых организмов является максимальное обеспечение устойчивости неравновесного термодинамического состояния, осуществляемое путем непрерывного чередования циклов, содержащих фазы синтеза АТФ из питательных веществ и его последующего расщепления.
На эту основную цель направлена вся совокупность деятельности клеток, органов, систем и целостного организма.
Поэтому при исследованиях их функционирования необходимо исходить из того, что все реакции и процессы в живых организмах направлены именно на эту цель.
Тогда, возможно, станут более понятны те сложнейшие процессы, которые происходят в живом организме, а также определена их закономерность.
СПИСОК ЛИТЕРАТУРЫ
1. Рубин А.Б. Термодинамика биологических процессов. М., 1984.
2. Николис Г., Пригожин И. Самоорганизация в неравновесных системах. М., 1979.
3. Бауэр Э.С. Теоретическая биология. Росток. 2002.
4. Доброборский Б.С. Термодинамика биологических систем. Учебное пособие для студентов высших медицинских учебных заведений. Под редакцией профессора Е.С. Мандрыко. Санкт-Петербург 2006.