| Похожие рефераты | Скачать .docx |
Реферат: Зоотехника
КОНТРОЛЬНАЯ РАБОТА №2
Гр3-75 «Зоотехника»
ВОПРОС №1 ВЛИЯНИЕ КОРМЛЕНИЯ НА РОСТ. РАЗВИТИЕ, ВОСПРОИЗВОДИТЕЛЬНУЮ СПОСОБНОСТЬ, ПРОДУКТИВНОСТЬ И ПЛЕМЕННЫЕ КАЧЕСТВА ЖИВОТНЫХ
Для сохранения здоровья, нормального роста и развития, репродукции, получения высокой продуктивности организм животного требует беспрерывного, в течение всей жизни, поступления из внешней среды кислорода воздуха, воды и пищи (корма). При отсутствии всех или любого из этих факторов организм существовать не может. Эти обязательные, природные, незаменимые факторы являются потребностями животного.
Поглощение пищи (корма) с физиологической точки зрения представляет сложный безусловный рефлекс, связанный с деятельностью головного мозга.
Характерной особенностью животного организма является непрерывный обмен веществ, составляющий основу жизни. Обмен веществ между животным организмом и окружающей средой регулируется нервной системой; он охватывает все проявления жизни организма, определяет функциональное состояние и характер деятельности органов, тканей, являясь реакцией организма на внутренние и внешние воздействия.
Для управления развитием и продуктивностью животных необходимо знать способы регуляции обмена веществ в организме, знать, каким по величине и характеру должен быть обмен веществ, для того чтобы в результате получалась желательная продуктивность или работоспособность животного.
Среди внешних условий, влияющих на количественную и качественную стороны обмена, первенствующая роль принадлежит кормлению, обеспечивающему жизнь животного. Основным условием установления зависимости между притоком пищи, состояния организма и его жизненных функций является знание вопроса о потребности животного в корме. Отсюда, количество и качество питательных и биологически активных веществ, необходимых животному при том или другом состоянии и использовании, характеризует его потребность в пище при данных условиях.
Недостаток или чрезмерный избыток в корме необходимых питательных веществ или неспособность организма использовать их изменяют течение биохимических процессов, нарушают нормальные жизненные функции, снижают продуктивность животных и нередко вызывают заболевания.
Потребность животного в конкретном питательном веществе определяют, пользуясь классической методикой синтетического рациона. Синтетическим называется рацион, составленный из отдельных химически чистых или высокоочищенных веществ. В таком рационе можно произвольно менять пропорции питательных веществ или целиком удалять из него те или иные вещества.
Если животное кормить синтетическим рационом, в котором изъято какое-то вещество, и животное в ответ на это кормление реагирует потерей аппетита, снижением продуктивности, ухудшением здоровья, то есть основание полагать, что изъятое из рациона вещество является необходимым для животного. После того, как в результате кормления рационом с изъятым веществом появляются признаки расстройства обмена, изъятое вещество вводится в рацион. На полном рационе животное поправляется. Это и будет новым доказательством, что животное нуждается в данном веществе. Таким путем была показана потребность животных в белке, макроэлементах, витаминах, а также в клетчатке, отдельных углеводах, жире, аминокислотах, микроэлементах и других веществах.
Методом синтетического рациона определяют не только потребность и незаменимость того или иного питательного вещества, но и количество, в котором оно должно поступать с кормом. Этим методом ветеринарные врачи пользуются для выяснения значения и потребности того или иного вещества у лабораторных животных.
Для установления потребности сельскохозяйственных животных в энергии, питательных и биологически активных веществах применяют и другие методы: анализ массовой практики кормления животных в хозяйствах, начно-хозяйственные опыты и опыты с определением баланса веществ и энергии в организме, а также изучение состояния животных по клиническим показателям.
Признаками обеспечения потребностей животного организма являются нормальное физиологическое состояние, обусловленное действием того или иного питательного вещества, и оптимальное содержание в теле или в продукции вещества, которое тесно связано с соответствующим фактором питания. Так: например, содержание витамина А в молоке животного зависит от поступления в организм самого витамина или его предшественника — каротина.
Показателями потребности животных являются: 1) определенное количество сухого вещества, которое характеризует объем кормовой дачи и сытость животного. Достаточный, но не обременяющий объем необходим для нормального наполнения пищеварительного тракта, при котором его деятельность протекает более успешно; 2) определенное количество общего органического вещества в доступной для усвоения форме, которое выражается величиной обменной энергии или в кормовых единицах, Количество органического вещества определяет общий уровень кормления животных; 3) определенное количество азотсодержащих веществ, также в доступной для усвоения форме, которое выражается в сыром и переваримом протеине или белке, в валовом содержании незаменимых аминокислот; 4) оптимальное количество сырой клетчатки, обеспечивающее работу пищеварительного тракта и возможное переваривание микроорганизмами, а также легкоусвояемых углеводов (сахара и крахмала); 5) оптимальное количество липидов (жиров), имеющих самую высокую калорийность и обеспечивающих животных жизненно необходимыми жирными кислотами (линолевой, линоленовой, арахидоновой и др.) и жирорастворимыми витаминами; 6) определенное количество минеральных веществ (поваренной соли, макро- и микроэлементов) в правильном соотношении, хотя они и не имеют энергетической ценности; 7) определенное количество витаминов как с индуктивным действием на организм животного (A, D, Е, С, В4), так и с биокаталитическим действием (комплекс витаминов В и витамин К).
Имеется определенная связь между потребностями животных в отдельных питательных веществах. Например, потребность в протеине зависит от суммарного содержания сухого вещества, в витамине Bi — от наличия в корме углеводов, в никотиновой кислоте — от содержания в корме протеина и т. д.
Потребности животных в питательных веществах в количественном и качественном отношении непостоянны. Они зависят от вида, возраста, физиологического состояния и использования животных. Для практического животноводства наибольшее значение представляет знание потребностей животных при поддерживающем кормлении, в связи с ростом, репродукцией, лактацией, откормом и работой.
Потребность животных в питательных и биологически активных веществах для поддержания жизни. В хозяйствах приходится содержать в течение некоторого времени взрослых животных, не дающих продукции и не используемых для работы, как, например, гулевых лошадей при продолжительном периоде покоя, холостых взрослых маток, взрослых производителей в неслучной период и др. В таком состоянии животные ничем не окупают расходов по кормлению и уходу, и если они здоровы и нормально упитанны, то кормление организуют так, чтобы поддержать их в этих кондициях с наименьшими расходами корма (передержка). Передержку животных применяют и в условиях недостатка кормов. В этом случае потребность в энергии и питательных веществах у животных минимальная и кормление называется поддерживающим.
Потребность в энергии и питательных веществах определяется затратами на работу органов кровообращения, дыхания, пищеварения, выделения и т.д. на необходимую мышечную деятельность и на поддержание температуры тела.
Потребность для поддержания жизни у животных разных видов зависит, прежде всего, от их живой массы. Чем больше масса тела животного, тем выше его потребность. Например, взрослой корове, имеющей среднюю упитанность, при живой массе 300 кг на поддержание жизни требуется в сутки 3,2 корм, ед., или 36 МДж обменной энергии, при массе 400 кг — 4,0 корм, ед., или 45 МДж энергии, при массе 500 кг — 4,7 корм, ед., или 54 МДж, при массе 600 кг — 5,5 корм, ед., или 63 МДж обменной энергии. Эти затраты энергии на поддержание жизни приблизительно пропорциональны живой массе в степени 0,73-0,75.
Животные разных видов и пород соответственно своей природе (наследственности) требуют для роста и развития различное количество питательных веществ. Зная природные требования молодых животных в разные периоды жизни к условиям кормления и содержания и реакцию организма на изменение этих условий, можно управлять формированием организма в период роста. Кормление молодых животных с учетом их потребности в питательных веществах ускоряет рост, способствует увеличению их конечной живой массы и улучшает телосложение. В противном случае, помимо задержки в осте, у животных появляются тяжелые патологические изменения в организме, иногда непоправимые (недоразвитие, незаразные болезни, впоследствии — низкая продуктивность).
Для определения потребности растущих животных в питательных запасах, необходимо знать, как изменяется в процессе роста химический состав организма в целом и состав прироста в разном возрасте. Для этого доводят химический анализ тканей и органов животных, убитых в разном возрасте, или в балансовых опытах определяют отложение в организме бел-i, жира и минеральных веществ у растущих животных.
Характерным показателем роста молодых животных является увеличение в теле главным образом количества белка и минеральных веществ. Отклонение белка происходит вследствие увеличения числа и размера клеток и тканей, повышения содержания минеральных веществ — в результате роста тканей и минерализации костяка. Содержание белка и минеральных веществ быстро нарастает в первые 5-10 месяцев жизни животного.
С возрастом изменяется также содержание в организме воды и жира. Содержание воды в организме с возрастом понижается (с 74% при рождении до 53% у взрослых). Уменьшение процента воды во всем организме происходит вследствие постепенного, с возрастом, обеднения водой органов и тканей и увеличением в теле жира. Особенно значительны возрастные изменения в содержании воды в мышцах и костной ткани. Обеднение тканей водой происходит в связи с изменением коллоидного состояния белков протоплазмы, они становятся более грубодисперсными и менее гидрофильными, В связи с этик создаются менее благоприятные условия для интенсивного обмена веществ. С понижением обмена веществ понижается жизнедеятельность клеток, снижаются процессы синтеза, а следовательно, и уменьшается скорость роста организма.
Чтобы обеспечить высокий прирост, растущие животные требуют большого количества легкопереваримых полноценных питательных веществ. В первую очередь растущему животному требуется определенное количество полноценного протеина. Белок является важнейшим веществом в составе тканей и органов растущего организма, рост связан с увеличением белка в организме. Наиболее интенсивно отложение белка идет в первые месяцы жизни, затем постепенно уменьшается и с окончанием роста, почти прекращается.
На отложение белка и скорость роста наряду с возрастом животных оказывает влияние уровень протеинового питания, причем он влияет не только на скорость роста и отложение белка, но и на соотношение жира к белку в приросте растущих животных; с повышением содержания протеина в корме понижается процент жира в приросте. Однако избыток протеина в рационе не оказывает положительного влияния на отложение белка в теле и не ускоряет роста, но понижает использование кормового протеина растущими животными.
Для нормального роста всем животным требуются витамины. Недостаток витаминов в кормах замедляет рост молодняка и вызывает многие заболевания.
Потребность в витаминах зависит от вида и возраста животных, интенсивности роста (суточного прироста), состава кормового рациона, состояния здоровья, условий содержания и др. В раннем возрасте молодые животные нуждаются в большем количестве витаминов на единицу массы тела по сравнению с более взрослыми. При кормлении рационами, хорошо сбалансированными по органическим и минеральным веществам, потребность в витаминах ниже, чем при скармливании рационов менее полноценных.
Рост животных сопровождается при полноценном кормлении увеличением в теле содержания жира. Большая часть жира откладывается в «жировом депо» организма — в подкожной клетчатке, в межмускульной соединительной ткани и в брюшной полости. Этот жир называется резервным, состоит преимущественно из глицеридов пальмитиновой, стеариновой и олеиновой кислот, он используется организмом как источник энергии.
Наряду с этим жиры входят в состав протоплазмы и принимают участие в клеточном обмене. Этот жир называется структурным (тканевым). Наиболее известными представителями структурного жира являются фосфолипиды.
Установлено, что животные могут синтезировать жир из углеводов, однако известно, что некоторые жирные кислоты, которые не синтезируются в организме растущих животных, должны поступать в корме. К ним относятся линолевая, линоленовая и арахидоновая жирные кислоты, которые для молодняка являются незаменимыми. При недостатке этих кислот у растущих животных появляются различные заболевания, главным образом кожного покрова (шелушение кожи, язвы, воспалительные явления на голове, конечностях, хвосте и др.). Тем не менее, нормы потребности их не установлены.
Потребность в сыром жире и его нормы установлены лишь для молодняка крупного рогатого скота и поросят до 20 кг живой массы, а также для плотоядных животных. Например, телятам сырого жира требуется в возрасте 1 месяца — около 4,5 г, 3 месяцев — 2,5 г, 6 месяцев — 1,5 г, 9 месяцев — 1,2 г на 1 кг живой массы. Потребность в сыром жире у поросят с живой массой 6 кг составляет 6 г, с массой 10 кг — 3,8 г и с массой 18 кг — 2,4 г на 1 кг живой массы.
Потребность животных в питательных и биологически активных веществах в связи с репродукцией. В любой отрасли животноводства необходимо иметь маток с нормальной плодовитостью, высокой оплодотворяемостью и жизнеспособным приплодом. Б числе факторов, влияющих на воспроизводство, большое значение принадлежит кормлению.
Длительное недокармливание вызывает прекращение овуляции у самок, дегенерацию половых клеток и атрофию семенников у самцов; у беременных животных приводит к абортам или резобции плода, удлинению периода беременности, к рождению слабого, недоразвитого приплода, весьма восприимчивого к заболеваниям пищеварительного канала и дыхательных путей.
Вредно действует на плодовитость и систематическое перекармливание, в этом случае на почве гипофункции гипофиза наблюдается жировое перерождение и инфильтрация яичников.
На плодовитость животных большое влияние оказывает состав кормового рациона по содержанию полноценного протеина, жира, углеводов, минеральных веществ и витаминов.
Рационы с низким содержанием протеина или с неполноценным протеидом, не удовлетворяющие потребность самок, приводят к бесплодию. Значительный избыток протеина в рационах по сравнению с нормой вызывает патологические изменения в яичниках и может привести к стерильности.
Недостаток в рационах жира нарушает половую функцию как у самок, так и у самцов. Животные не обладают способностью синтезировать в организме непредельные жирные кислоты (линолевую, линоленовую, арахидоновую и др.), которые необходимы для синтеза мужских и женских половых гормонов
Из безазотистых питательных веществ имеют значение для воспроизводства животных углеводы (крахмал, сахар, клетчатка). Недостаточная энергетическая ценность рациона задерживает половое созревание, наблюдаются перелы. Избыточное энергетическое питание ведет, как правило, к яловости потребность в энергии в период беременности самок увеличивается с возрастом - ода. Главной причиной пониженной плодовитости у животных является достаточная калорийность корма в последнюю четверть периода беременности. Репродукция животных во многом зависит от наличия минеральных веществ в рационе. Часто причиной яловости и абортов является недостаток в кормах кальция и фосфора. У животных иногда наблюдается полная стерильность при одновременном недостатке в рационе кальция и протеина. При этом у самцов наблюдаются патологические изменения в семенниках и дегенерация половых клеток. Ярким показателем недостатка кальция в рационе является увеличение числа мертворожденных поросят, причем с каждым последующим пометом количество мертворожденных возрастает. При этом у поросят, рожденных живыми, наблюдаются патологические изменения в костной ткани. Результатом недостаточного снабжения кальцием обычно является плохая молочность маток после опороса, выраженная анемия и лимфоцитоз.
Недостаток кальция в корме оказывает отрицательное влияние и на репродукцию крупного рогатого скота. Коровы при этом плохо оплодотворяются и дают слабых, плохо развитых телят.
Нарушения, а репродукции животных отмечаются из-за недостатка в рационах йода. Приплод в таких случаях рождается слабым (часто мертвым), с голой кожей, наличием зоба. Установлена зависимость репродукции и от содержания в кормах цинка, натрия, калия, железа, кобальта, марганца и других элементов. В частности, при недостатке марганца в рационах бычки дают мало семени, концентрация сперматозоидов низкая, они малоподвижны.
Большое значение для нормальной репродукции животных имеют витамины. Недостаток в рационах витаминов, отрицательно влияет и на репродукцию. Например, недостаток в кормах витамина А вызывает у самцов нарушение образования семени и дегенеративные изменения эпителия семенных канальцев, в результате ухудшается качество семени и понижается половая активность. У самок на А-авитаминозных рационах происходят изменения в слизистой оболочке вагины, матки, плаценты (кератинизация. некроз), что затрудняет оплодотворение, препятствует имплантации оплодотворенного яйца, нормальному питанию эмбриона и приводит к смерти и рассасыванию плода, к абортам или к рождению слабого, иногда уродливого потомства. На ранних стадиях авитаминоза яичники не поражаются, и овуляция происходит регулярно, затем наступает атрофия яичников, приводящая к дегенерации яйцевых клеток.
Недостаток витамина D в рационе беременных животных вызывает расстройство в кальциевом и фосфорном обмене у матери и плода. Репродукция зависит и от витамина Е. Недостаток этого витамина вызывает у самцов патологические изменения в генеративном эпителии и в образовании семени, у самок — раннее прекращение беременности. Е-витаминная терапия оказывается полезной для устранения бесплодия у крупного рогатого скота. свиней и других животных.
Способность жвачных животных синтезировать витамины группы В рубце делает их независимыми от содержания этих витаминов в рационе. У свиней, лошадей и других животных при недостатке в корме витаминов группы В оплодотворение не происходит или беременность протекает патологически.
Для нормальной репродукции необходимо наличие в кормах и в организме достаточного количества витамина С (аскорбиновой кислоты). Содержание аскорбиновой кислоты в крови у нормальных в половом отношении животных обычно выше, чем у животных с нарушенной продукцией. Максимальная потребность в аскорбиновой кислоте у самок наблюдается в ранний период беременности, у самцов — в период случки. Животные обладают способностью синтезировать в организме витамин С, но скорость синтеза зависит от многих условий и часто его синтезируется меньше, чем требуется. Терапия аскорбиновой кислотой является успешной при устранении импотенции у производителей и плохой оплодотворяемости у маток.
Таким образом, для обеспечения нормальной репродукции необходимо нормировать потребность животных в питательных веществах. Недостаточное питание несбалансированными рационами в отношении энергии, протеина, жира, углеводов, минеральных веществ и витаминов отрицательно влияет на репродукцию животных.
Особенности кормления беременных животных. Для получения хорошо развитого жизнеспособного потомства необходимо правильно кормить беременных животных. У млекопитающих оплодотворенное яйцо развивается внутри материнского организма, который снабжает эмбрион питательными веществами. Такой способ питания обусловливает огромное влияние материнского организма на эмбриональное развитие. Он же предохраняет эмбрион от резких колебаний в поступлении питательных веществ, так как мать при необходимости обеспечивает, в известной мере, плод за счет своего тела.
Кормление беременных животных должно быть организовано так, чтобы они в хорошо сбалансированных рационах получали достаточное количество питательных веществ на развитие жизнеспособного приплода и сами в течение всей беременности имели хорошее здоровье.
Беременность вызывает весьма значительные изменения во всем организме матери. Внешними показателями этих изменений является увеличение живой массы на 10-25%. Обычно в первую половину беременности эти изменения невелики, а затем масса тела быстро увеличивается, особенно к концу беременности.
Увеличение массы животных является результатом роста плода, увеличения матки, молочных желез и при хорошем кормлении — отложения некоторых резервов в организме самой матери.
Количество питательных веществ, необходимое для отложения резервов, зависит от упитанности маток перед покрытием, их возраста и ожидаемой молочности. Эти резервы имеют большое значение после родов в обеспечении нормальной лактации, когда питательные вещества рациона не покрывают полностью потребности лактирующего животного, особенно в первые дни после родов.
Беременность существенно изменяет обмен веществ у самок. Эти изменения вызываются усиленной деятельностью желез внутренней секреции, в первую очередь щитовидной и гипофиза, и постепенно увеличивающимися затратами матери на питание плода. Значительно увеличивается газообмен и теплопродукция. В среднем обмен у сельскохозяйственных животных увеличивается за весь период беременности на 11-14%. Заметное усиление обмена наблюдается уже в конце первой половины беременности и весьма значительное (до 30-40%) — в последней стадии.
Беременность оказывает специфическое влияние на белковый и минеральный обмен в организме. Рост плода, увеличение матки и других органов связано с отложением в теле значительных количеств белка, кальция, фосфора, железа и других зольных элементов. На интенсивности обмена сильно сказывается многоплодность.
При рождении телята имеют живую массу от 20 до 50 кг, жеребята — от 40 до 60 кг, помет поросят — от 12 до 15 кг, ягнят — от 4 до 5 кг. Новорожденные животные содержат в своем теле 75-80% воды, 12-18% белка, 1,5-4,0% жира и 3,0-4,5% минеральных веществ. Таким образом, для роста и развития плода за весь период беременности требуется большое количество органических и минеральных веществ. Например, в теле новорожденного теленка содержится до 9 кг белка. При средней биологической ценности протеинов кормов в обычных рационах около 65% стельным коровам только на формирование плода требуется до 15 кг протеина. Кроме того, корова должна получать определенное количество протеина на отложение белка в виде резерва в организме.
Потребность беременных животных в питательных веществах отражена в нормах кормления животных разных видов.
Потребность беременных коров до 8-месячной стельности учитывается в нормах лактирующих животных, потребность в последние два месяца стельности предусмотрена в нормах стельных сухостойных коров, которые зависят от живой массы и предполагаемого удоя в следующую лактацию.
Потребность супоросных свиноматок в питательных веществах определяется возрастом, живой массой и периодом супоросности. Наиболее низкая потребность у свиноматок в первые 84 дня супоросности, поскольку в это время у них относительно невысокий обмен веществ при очень малом отложении питательных веществ в плоде и теле. В последний месяц супоросности у маток обмен веществ возрастает, отложение энергии и белка увеличивается в 8-10 раз.
Потребность беременных овец и коз в питательных веществах зависит от живой массы, направления продуктивности и периода суягности. Нормами для них предусмотрена потребность в первые 12-13 недель и последние 7-8 недель суягности.
Потребность беременных кобыл в питательных веществах зависит от породы, живой массы и физиологического состояния. Потребность жеребых кобыл до 9-месячной жеребости предусмотрена в нормах лактирующих кобыл, поэтому нормирование потребности жеребых кобыл начинают с 9-месячной жеребости.
Особенности кормления племенных производителей. Особенностью потребности производителей в питательных веществах является влияние кормления на количество, и качество семени, которым определяется оплодотворяющая способность производителей. При каждой садке бык выделяет в среднем 4-5 мл спермы, жеребец — 50-60, баран — 1-1,5, хряк — до 400-500 мл и более. Сперма содержит от 90 до 97% воды, от 1,2 до 8,7% органических веществ, от 0,6 до 0,9% минеральных элементов. Из органических веществ 1,2-2,0% составляет белок и около 0,2% липоиды (сложные жиры). Белок представлен альбуминами, глобулинами, нуклеопротеинами, муцином и альбумозами. В составе минеральных веществ преобладают кальций и фосфорная кислота. Нормы потребности производителей разработаны для быков и хряков с учетом живой массы и половой нагрузки; для 5аранов — с учетом направления овцеводства, живой массы и половой нагрузки; для жеребцов — с учетом породы, живой массы и режима полового использования.
Если случка производителей носит сезонный характер, то нормирование потребности в питательных веществах и подготовку племенных производителей начинают за месяц-полтора до начала случного периода, так как процесс образования половых клеток и прохождение их по каналу придатка семенников продолжается от 2 до 4 недель и, следовательно, влияние кормления на качество семени скажется не ранее этого срока.
Потребность животных в питательных и биологически активных веществах в связи с лактацией. Молоко у сельскохозяйственных животных вырабатывается за счет питательных веществ корма. Однако составные вещества молока— казеин, молочный альбумин, молочный сахар и молочный жир — не встречаются в обычных кормах и по своей химической природе сличаются от составных частей тела и крови. Молоко содержит примерно в 10 раз больше липидов и углеводов и вдвое меньше белка, чем кровяная сыворотка; молочного сахара и казеина в крови совсем нет; молочный альбумин не тождественен с альбумином крови; молоко богаче кальцием в 14 раз, фосфором — в 9 раз. Все питательные вещества корма, поступающие в кровь, должны подвергаться коренной переработке, прежде чем войти в состав молока. Эта переработка происходит в альвеолах молочной железы. Работа молочной железы связана с материнством и регулируется сложными нервно-моральными реакциями. Молочная железа работает очень интенсивно. коровы, например, молочная железа составляет лишь 2-3% массы животного, но выделяет за год в молоке сухого вещества в 3-4 раза больше, чем содержится во всем теле. В период лактации для образования 1 л молока через молочную железу протекает до 500-600 л крови.
Молочный сахар образуется из глюкозы, содержащейся в плазме крови. Белки молока синтезируются из аминокислот, белков и полипептидов крови. Основным материалом для образования молочного жира служат липиды крови (нейтральный жир, фосфатиды); у жвачных животных в образовании жира молока участвуют летучие жирные кислоты (преимущественно уксусная), образующиеся в рубце при брожении углеводов.
Огромное влияние на молокообразование и молокоотделение оказывает кормление. Зависимость лактации от кормления имеет много общих черт для всех видов животных. Недостаточное и неполноценное питание изменяют нормальный ход лактации и состав молока. Эти нарушения в лактации происходят из-за недостатка в организме органических и минеральных веществ, используемых для образования молока, а также вследствие изменений в функциях органов и систем, которые прямо или косвенно влияют на молочную секрецию.
Установлено, что на лактацию оказывает большое влияние уровень биологически полноценного протеина. Протеиновое питание лактирующих животных в первую очередь обеспечивает получение не только максимального количества молока, но и молока с высоким содержанием жира и белка. Потребность лактирующих животных в протеине, как и в других питательных веществах, определяется в первую очередь функциональным состоянием молочной железы, которое зависит от периода лактации, общего состояния организма, полноценности протеина и др. Неполноценность протеина в рационе ограничивает молочную продуктивность и вызывает распад тканевого белка тела для снабжения молочной железы недостающими в корме аминокислотами. Как минимум, на образование 1 кг молока требуется следующее количество незаменимых аминокислот (г): лизина — 2,3, метионина — 1,2, триптофана — 0,8, аргинина — 1,3, гистидина — 0,6, фенилаланина — 1,2, лейцина — 3,6 и валина — 2,5.
Лактирующим коровам, овцам и свиньям на образование молока требуется примерно в 1,5 раза больше переваримого протеина, чем содержится белка в молоке. В этом случае на 1 кг молока (без протеина на поддержание жизни) требуется переваримого протеина (г): коровам — 60, овцам — 100, лошадям — 40-45 и свиньям — 90. По ныне существующим нормам кормления лактирующим животным в расчете на 1 корм. ед. рациона требуется следующее количество переваримого протеина (г): коровам — 95-110 (в зависимости от суточного удоя), овцам — 90-110 (в зависимости от живой массы, периода лактации и направления овцеводства), свиньям — 110-112 (в зависимости от возраста, живой массы и количества поросят в приплоде), лошадям — 108-110 (в зависимости от живой массы и направления коневодства). У лактирующих животных белковый обмен идет очень интенсивно в первую половину лактации, поэтому максимальное количество переваримого протеина требуется в этот период. Скармливание кормов в рационе с содержанием протеина выше нормы приводит к белковому перекорму, что может быть причиной патологического состояния организма, появляются почечные патологии, в крови повышается концентрация различных фракций небелкового азота.
Лактирующим животным требуется определенное количество жира. Для образования молочного жира животные используют не только жир корма, но и углеводы и протеин. Установлено, что животные с молоком выделяют значительно больше жира, чем получают в корме. Имеется тесная зависимость между липидами крови и содержанием жира в рационе, с понижением количества жира в кормах падает концентрация жирных кислот, фосфатидов и холестерина в плазме крови.
Количество жира в рационе и его физико-химические свойства оказывают определенное влияние на свойства молочного жира. На рационах с недостатком жира животные продуцируют молоко с пониженным содержанием жира, который при этом имеет низкое йодное число и крайне мало непредельных жирных кислот.
У жвачных животных в вымени синтезируется значительная часть молочного жира из летучих жирных кислот, образующихся при брожении корма в рубце, около 50% молочного жира дают непосредственно липиды плазмы крови.
Количество необходимого корове жира составляет около 65% от общего количества жира, выделяемого в молоке, свиноматкам — около 30%. В существующих ныне нормах кормления потребность в сыром жире установлена только для лактирующих коров, она составляет от 28 до 40 г на 1 корм. ед. рациона.
Лактирующим животным требуется определенное количество углеводов. Жвачным углеводы необходимы для нормального обмена в рубце. Для этого нужны клетчатка и легкоусвояемые углеводы (сахар, крахмал). Продукты ферментации углеводов — летучие жирные кислоты — являются предшественниками молочного жира. Нормами кормления предусмотрена, потребность коров в сахаре и крахмале, для других животных — только в клетчатке. Например, дойной корове в зависимости от суточного удоя требуется :ахара от 75 до 120 г, крахмала — от 110 до 180 г на 1 корм. ед. рациона. Потребность в клетчатке нормируют в расчете на 1 кг сухого вещества рациона: дойным коровам требуется от 16 до 28% (в зависимости от суточного удоя: чем выше удой, тем меньше норма клетчатки), свиноматкам — не более 290-370 г в сутки (в зависимости от живой массы и количества поросят в помете).
Большое значение в кормлении лактирующих животных имеют минеральные вещества. Они нужны для поддержания жизни в здоровом состоянии, для нормального усвоения питательных веществ корма и для образования молока. 3 1 кг молока выделяется значительное количество минеральных веществ (г): коровы — 7,5, у овцы — 9,5, у кобылы — 5,2 и у свиньи — 9,8. Например, 1 кг коровьего молока содержится (г): кальция — 1,19, фосфора — 0,84, калия — 1,48, натрия — 0,72, хлора — 1,37, магния — 0,14 и т. д. В золе молока всех животных преобладают калий, кальций, фосфор и хлор.
Минеральный состав молока считается более или менее постоянным, однако содержание некоторых элементов варьирует в связи с изменением состава органических веществ в молоке, а также с ходом лактации. Существует
положительная корреляция между процентом жира в молоке и содержанием в нем кальция, магния и фосфора. К концу лактации повышается содержание в молоке хлора, натрия и кальция. При неблагоприятных условиях кормления в молоке снижается содержание кальция.
На минеральный обмен у лактирующих животных оказывает большое влияние уровень молочной продуктивности и состав, размер и состав кормовой дачи, и другие факторы, например, количество выпиваемой животными воды. Кроме того, животные обладают способностью накапливать в своем теле значительные резервы минеральных веществ, которые также оказывают влияние на минеральный обмен.
В начальной стадии лактации у большинства млекопитающих животных часто наблюдается отрицательный баланс минеральных веществ, особенно кальция и фосфора, даже при обильном по содержанию органических и минеральных веществ кормлении. Животные в начале лактации из обычных рационов не всегда могут усваивать столько кальция и фосфора, сколько требуется для возмещения затрат на образование молока, и вследствие этого заимствуют минеральные вещества из организма. В этот период важную профилактическую роль играют резервы минеральных веществ. Эти резервы сосредоточены в губчатом веществе костей, главным образом трубчатых, отсюда при необходимости заимствуется кальций и фосфор. Если животное вынуждено продолжительное время жить за счет резервов, не восстанавливаемых кормлением, тогда затрачивается не только губчатый, но и плотный основной слой костной ткани, костяк обедняется минеральными веществами, делается пористым, непрочным, у лактирующих животных появляется заболевание — остеопороз.
Важная профилактическая роль минеральных резервов организма в начале лактации указывает на необходимость создавать эти резервы у животных во вторую половину лактации и вне лактации, а у молодых животных — в период роста и подготовки к первой лактации.
В современных нормах кормления потребность лактирующих животных в минеральных веществах определена по их содержанию в молоке и данных об усвоении элементов животными из кормовых рационов по результатам балансовых опытов, с учетом содержания в крови. Считается, что в среднем лактирующие животные усваивают для молокообразования из обычных кормовых рационов около 30-50% содержащихся в них минеральных веществ. Поэтому в корме их должно быть в 2-3 раза больше, чем выделяется с молоком.
Кроме протеина, жира, углеводов и минеральных веществ, в кормлении лактирующих животных большое значение имеют витамины. Они нужны не только для поддержания в норме всех жизненно важных физиологических процессов и здоровья животных, но и для получения богатого витаминами молока, необходимого для питания человека и выращивания новорожденных животных.
Содержание витаминов в молоке сильно варьирует в зависимости от условий кормления и содержания. Установлено, что содержание в молоке, например, витамина А и каротина зависит только от кормления. Животные разных видов различаются по способности превращать каротин в витамин А. Свиньи, овцы, козы более совершенно переводят каротин в витамин, чем крупный рогатый скот, а плотоядные такой способностью не обладают. Вследствие этого лактирующие животные разных видов, при одинаковой обеспеченности каротином в корме, дают молоко с разным соотношением в нем желтоокрашенного каротина и бесцветного витамина А.
Наблюдаются различия в способности превращать каротин в витамин и между породами скота. Например, коровы джерсейской породы дают молоко с более интенсивно окрашенным молочным жиром, чем коровы голландской и швицкой пород. Витаминная ценность молока в первом случае обязана преимущественно каротину, тогда как во втором случае — витамину А.
Потребность лактирующих животных в каротине установлена для всех видов: для коров — 40-50 мг, овец — 9-10 мг, свиней — 8-9 мг и лошадей — 18-20 мг на 1 корм. ед. рациона.
При резкой недостаточности витамина D у лактирующих животных наблюдается снижение кальция и фосфора в крови, ограничение движений и опухание суставов. Кормление и содержание лактирующих животных оказывает прямое влияние на концентрацию витамина D в молоке. Богатое витамином D молоко (до 25-40 ME в 1 л) дают животные летом и очень бедное (до 3-4 ME в 1 л) — зимой, при стойловом содержании. Главным фактором, обусловливающим содержание в молоке витамина D, является действие солнечного света на животное и скармливание облученных дрожжей и препаратов витамина. Потребность лактирующих животных в витамине D в среднем составляет у коров — около 1 тыс. ME, у овец — 500 ME, у свиней — 450 ME и у лошадей — 600 ME на 1 корм. ед. рациона.
В период лактации животные нуждаются в повышенных дозах витаминов комплекса В и др. Потребность животных в тиамине (Bi) связана с количеством получаемых в корме углеводов. Для лактации необходимы рибофлавин (Bg), никотиновая кислота (Вз) и др., которые влияют на витаминный состав молока, в то время как содержание аскорбиновой кислоты (С) в молоке мало или совсем не зависит от кормления. Считается, что сельскохозяйственные животные обладают способностью синтезировать аскорбиновую кислоту в своем организме и выделяют молоко с более или менее постоянным содержанием витамина С (у коров — около 20-30 мг в 1 л).
В витаминах группы В в большей степени нуждаются лактирующие свиноматки, кобылы и собаки, потребность в которых установлена и предусмотрена нормами кормления.
Потребность лактирующих коров в сухом веществе, кормовых единицах и обменной энергии, характеризующая общий уровень кормления, зависит от вида, возраста, живой массы, продуктивности, направления животноводства и других факторов. Например, лактирующим коровам в среднем требуется 2,8-3,2 кг сухого вещества в расчете на 100 кг живой массы, высокопродуктивным — 3,5-3,8 кг и в отдельных случаях — до 4-4,7 кг. Чем выше удои коровы, тем больше требуется энергии в 1 кг сухого вещества рациона. При снижении концентрации энергии в рационе корова не может поедать достаточное количество кормов для удовлетворения потребности в энергии.
Недопустимо снижение энергии ниже 0,65 корм, ед., или 8 МДж энергии в 1 кг сухого вещества. У лактирующих коров с удоем 28 кг и более в сутки концентрация энергии в сухом веществе должна быть не менее 1,05 корм, ед., или 11,4 МДж обменной энергии.
Лактирующим овцематкам в среднем требуется 3,6-4,2 кг сухого вещества, 3-4 корм, ед., или 35-40 МДж обменной энергии на 100 кг живой массы. Потребность овцематок в общем уровне кормления повышается в первые 6-8 недель лактации и снижается во вторую половину лактации.
Потребность лактирующих свиноматок в общем уровне кормления нормируют в зависимости от живой массы, числа поросят в приплоде и продолжительности подсосного периода. В среднем лактирующей свиноматке с 10 поросятами на 100 кг живой массы требуется около 2,8 кг сухого вещества с содержанием 1,3 корм, ед., или 14,4 МДж обменной энергии в 1 кг. Потребность лактирующих кобыл в сухом веществе в среднем составляет 3 кг на 100 кг живой массы при содержании в 1 кг 0,8 корм, ед., или 8,37 МДж обменной энергии.
Потребность животных в питательных и биологически активных веществах в связи с откормом. Откормом называют избыточное кормление животных, предназначенных на убой, направленное на отложение в теле наибольшего количества структурных и резервных питательных веществ (белка, жира, минеральных элементов, витаминов и др.) в виде мяса и сала.
Сало состоит из переполненных жиром клеток соединительной ткани с редуцированной протоплазмой. Главной составной частью его является жир (от 76 до 93%), на долю белка, образующего оболочки и протоплазму клеток соединительной ткани, приходится от 2 до 4%.
Остающееся после отделения сала мясо состоит преимущественно из мускульной ткани. Как мускульная ткань, так и проходящие в ней кровеносные и лимфатические сосуды, нервы и связки являются по составу белковыми образованиями, в мясе в небольшом количестве (от 1 до 6%) всегда находится и жир. Соотношение между мясом и салом в тушах сильно меняется в зависимости от вида, возраста, степени упитанности, кормления животных и других факторов.
Для производства мяса используют два процесса — рост и откармливание животных. Рост свойственен только молодым животным, не закончившим своего развития, у растущих животных накопление мяса является результатом образования новых мускульных волокон.
Откармливание возможно во всяком возрасте при обильном кормлении. Хорошо откармливаются молодые, растущие животные, а также и взрослые, получающие значительный избыток питательных веществ сверх поддерживающего кормления.
По мере откорма, с улучшением упитанности в теле животных происходит ряд существенных изменений: изменяется химический состав тела, понижается содержание воды, белка и возрастает процент жира; изменяется распределение жира в теле. Жир откладывается, главным образом, в подкожной соединительной ткани и в брюшной полости (сальнике). В различных частях тела жир откладывается неравномерно, вследствие чего калорийность мяса из
разных частей тела изменяется при откорме неодинаково. Отложенный между волокнами и в межмускульной соединительной ткани жир придает мясу «мраморность», улучшает его вкус, сочность и повышает питательность. Жир мышечной ткани оказывает также влияние на цвет и плотность мяса и его сохранность при низкой температуре; повышается убойный выход, измеряемый отношением массы туши к живой массе животного.
У молодых откармливаемых животных потребность в общем уровне кормления определяется затратами питательных веществ на рост и на отложение в теле жира (собственно откорм).
У взрослых животных, поступающих на откорм в состоянии упитанности, близкой к средней, процесс откорма сводится к отложению в теле резервного жира.
В практике продуктивность откармливаемых животных измеряют среднесуточным приростом живой массы, в связи с этим и потребность в корме как у молодых, так и взрослых животных определяется величиной и составом прироста.
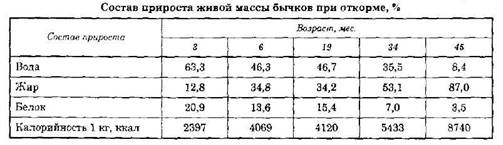
При откармливании молодых, растущих животных состав прироста изменяется в зависимости от возраста и степени упитанности (табл. 99). Например, у телят моложе трех месяцев прирост состоит преимущественно из белка (15-20%) и воды (до 60-70%), затем с возрастом в приросте животных постепенно увеличивается содержание жира, и у хорошо откормленных животных, закончивших рост, жир составляет около 90%.
Вследствие изменений в составе прироста увеличивается его калорийность. Аналогичные изменения наблюдаются и в составе прироста у откармливаемого молодняка других видов сельскохозяйственных животных.
При откорме взрослых животных прирост живой массы на 75-80% состоит также из жира, а в составе органического вещества он занимает до 90%. Многочисленными опытами доказано, что жир у откармливаемых животных образуется из жиров, протеинов и углеводов корма. Из этих основных групп питательных веществ, несомненно, самую важную роль в жирообразовании играют углеводы. Практикой кормления давно установлено, что для успешного откармливания животных лучшими кормами, богатыми углеводами, являются картофель, кукуруза, ячмень и др.
Жир корма оказывает влияние не только на отложение его в организме, но и на качество животного жира (сала), особенно при откорме свиней и птицы.
Сельскохозяйственные животные обычно получают в корме, можно сказать, чужеродный жир, отличающийся по химическому составу и физическим свойствам от жира, содержащегося в их теле. При небольшом содержании жира в корме животные полностью трансформируют его в жир, характерный для данного вида животных. При усиленном питании животных кормами, богатыми жирами, кормовой жир откладывается в теле, сохраняя свои характерные особенности и частично изменяя свойства резервного жира.
Откармливаемым животным требуется определенное количество протеина. Эта потребность тесно связана с их способностью к отложению в организме белка, что зависит в первую очередь от возраста животных, их природных особенностей и индивидуальных свойств. Нормально упитанные взрослые животные по своим биологическим особенностям не могут откладывать в своем теле значительного количества белка.
Величина отложения белка зависит не только от количества протеина в корме, но и от протеинового соотношения в рационе. При одном и том же содержании протеина в рационе отложение возрастает с увеличением безазотистых веществ (углеводов и др.). Поэтому в рационах откармливаемых животных регулируют не только потребность в протеине, но и в протеиновом соотношении. Откорм идет успешнее при широком протеиновом соотношении в рационе (1 : 10). При более широком соотношении (шире чем 1 : 10-12) наступает ухудшение переваримости корма.
Для откармливания молодых животных на единицу живой массы протеина требуется значительно больше, а протеиновое соотношение ближе к среднему (1 : 6-8). Откормочный молодняк крупного рогатого скота и подсвинки при мясном откорме откладывают в теле до 100-150 г белка в сутки. При недостатке протеина в корме рост молодых животных задерживается, мышцы не достигают полного развития, выход мяса в туше уменьшается.
Нормирование потребности откармливаемых животных в энергии производят в расчете на 1 кг прироста живой массы. Например, потребность в энергии у молодняка крупного рогатого скота при откорме в зависимости от живой массы и периода откорма составляет: в начале откорма — 6,1-7,0 корм, ед., или 51-61 МДж обменной энергии, в середине откорма — соответственно 7,9-8,2, или 69-74, в конце откорма — 9,2-9,7 корм, ед., или 85-107 МДж обменной энергии на 1 кг суточного прироста. Потребность в переваримом протеине для этих животных составляет в среднем 90-110 г на 1 корм. ед. рациона.
Откармливаемому молодняку крупного рогатого скота на 100 кг живой массы требуется (г): поваренной соли — 8-12, кальция — 9-21, фосфора — 6-9, магния — 4-6, калия — 17-20, серы — 7-10. На 1 кг сухого вещества рациона этим животным требуется (мг): железа — 50-70, меди — 7-10, цинка — 40-50, кобальта — 0,5-0,6, марганца — 30-50, йода — 0,2-0,4; каротина — 9-10, витамина D — 1,0-1,2 тыс. ME, витамина Е — 25 мг.
Потребность животных в питательных и биологически активных веществах в связи с работой. В сельском хозяйстве и транспорте используют в качестве рабочих животных лошадей, буйволов, волов, верблюдов, ослов, мулов, оленей, собак.
ВОПРОС №2 МЕТОДЫ ОПРЕДЕЛЕНИЯ ПЕРЕВАРИМОСТИ И СПОСОБЫ ЕГО ВЫЧИСЛЕНИЯ
Определение переваримости питательных веществ и энергии. Валовое содержание в корме питательных веществ и энергии не может служить показателем его истинной ценности, поскольку значительная часть питательных веществ корма не всасывается в желудочно-кишечном тракте, а выделяется с калом. унося при этом часть валовой энергии корма.
Более объективное представление о питательности корм дает наличие в нем переваримых питательных веществ. Для определения переваримости в опыте группе животных дают строго взвешенное количество корма и учитывают количество выделенного кала. Из скормленных кормов и выделенного кала за период опыта отбирают пропорциональные пробы, в которых определяют содержание питательных веществ (химическим путем) и энергии (путем/ сжигания в калориметрической бомбе).
В связи с тем, что в процессе переваривания, всасывания и межуточного обмена поступающие с кормом питательные вещества претерпевают существенные изменения, их «судьба» в процессе обмена может быть изучена лишь по косвенным показателям и прежде всего по балансу азота и углерода элементов, составляющих основу органических веществ. По балансу азота (азот корма = азот, отложенный в теле животного + азот кала + азот мочи) можно определить уровень отложения белка в теле животного, а также количество углерода, отложенного в белке.
По балансу углерода (углерод корма = углерод продуктов дыхания + углерод кишечных газов + углерод кала-f углерод мочи +углерод белка и жира), зная отложение азота, можно судить об отложении углерода в теле в виде жира (углерод отложения — углерод белка = углерод жира).
Поскольку значительное количество углерода покидает организм через органы дыхания, то определение его баланса требует постановки опытов с учетом газообмена. Для этого применяют респирационные установки, представляющие собой герметизированную и термостатированную камеру, оборудованную для кормления, поения, доения животного, сбора мочи и кала, а также имеющую устройство для точного учета количества вдыхаемого воздуха и автоматического отбора пропорциональных проб.
Крахмальные эквиваленты Кельнера. Чтобы установить продуктивное действие корма. Кельнер предварительно проведя серии респирационных опытов определил баланс азота и углерода при скармливании взрослому волу чистых питательных веществ: белка, жира, крахмала, тростникового сахара. Затем но балансу азота и углерода вычислил отложение белка и жира в организме. Пересчитав по энергетической ценности белок в жир (I г жира содержит 9,5 ккал, 1 г белка — 5,7 ккал).
Корма, для которых жирообразующая способность, рассчитанная по переваримым питательным веществам, не отличалась от таковой, рассчитанной по балансу азота и углерода, О. Кельнер отнес к группе полноценных кормов. Разницу между расчетным и фактическим (по балансу С и N) жироотложением он характеризовал как затрату энергии переваренных питательных веществ на переваривание клетчатки.
Экспериментальным путем Кельнер установил, что 100 г поступившей сырой клетчатки уменьшали образование жира на 14,3 г.
Величину фактического жирообразования, рассчитанную по балансу азота и углерода, отнесенную к величине жирообразования, рассчитанной по переваримым питательным веществам, О. Кельнер назвал коэффициентом полноценности. Так, в примере по определению продуктивного действия лугового сена расчетная жирообразующая способность 100 г его равна 12,75 г, фактическая — 8,1. Коэффициент полноценности лугового сена равен 0,63 (8,1 : 12,75). Для удобства пользования предлагаемой системой оценки кормов в практике О. Кельнер в качестве мерила продуктивного действия избрал жирообразующую способность крахмала.
Приняв ее за единицу, он вычислил, что для переваримого белка она равна 0,94 (23,5:24,8), для переваримого жира — 1,95—2,45 (47,4:24,8; 59,8:24,8), для переваримых безазотистых экстрактивных веществ и клетчатки— 1. Таким образом, зная содержание переваримых питательных веществ в любом корме и используя вычисленные эквиваленты, легко рассчитать питательную ценность корма в крахмальных эквивалентах. Например, в 100 кг сена содержится следующее количество переваримых питательных веществ: белка — 2,8 кг, жира — 1,6, безазотистых экстрактивных веществ — 23,9, переваримой клетчатки —3,5 и сырой клетчатки — 25,6 кг.
Если бы клетчатка переваривалась с такими же затратами энергии, что и безазотистые экстрактивные вещества, то 42,09 можно было бы считать крахмальным эквивалентом сена. Но в 100 кг сена содержится 25,6 кг сырой клетчатки, на переработку которой животное должно затратить энергию и тем самым снизить отложение жира. Как уже говорилось, 100 г поступившей сырой клетчатки снижает жироотложение на 14,3 г, то есть на ,58 крахмального эквивалента (14,3 :24,8). Следовательно, в данном случае продуктивная ценность сена снизится на 14,79 крахмального эквивалента (0,58-25,5) и составит 28,3 крахмального эквивалента (42,09 — 14,79).
Крахмальный эквивалент корма — это количество крахмала (и кг), эквивалентное по отложению жира 100 кг оцениваемого >корма.
Термы Армсби. Примерно в одно время с Кельнером профессор из Пенсильвании Армсби провел исследования, в результате которых также была предложена система оценки энергетической питательности корма по их продуктивному действию. В отличие от Кельнера он предложил оценивать энергетическую питательность кормов по энергии, отложенной в продукции, то ость по чистой энергии. В качестве единицы измерения чистой энергии он использовал I терм (1000 ккал, или 4,187 МДж). Под чистой энергией Армсби имел в виду ту часть энергии, которая остается в организме после вычитания из валовой энергии корма энергии кала, мочи и кишечных газов, а также потерь тепловой энергии, связанных с потреблением и перевариванием корма.
Таким образом, для определения чистой энергии, откладывающейся в организме при скармливании различных кормов. Армсби в опытах на животных должен был узнать: 1) энергию съеденного корма, 2) энергию выделенных кала и мочи, 3) энергию, выделенную с пищеварительными газами, 4) энергию, затраченную животным на поддержание температуры тела, мышечную работу и переработку корма, выделяющуюся в виде тепла.
Валовую энергию корма, кала и мочи определяли прямым калориметрированием (сжиганием в калориметрической бомбе), энергию пищеварительных газов (метана) — по их количеству (в опытах Кельнера и Армсби было установлено, что на каждые 100 г переваримых углеводов образуется 4,5 г метана), потери энергии в виде тепла определяли с помощью респирационного калориметра.
Оценка по сумме переваримых питательных веществ (СППВ). Эта система оценки питательности кормов широко распространена в США. СППВ рассчитывают по следующей формуле:
Переваримый сырой протеин+Переваримая сырая клетчатка
Потребленный корм
Переваримые БЭВ+(переваримый жир-2,25) Потребленный корм
Положительной стороной системы оценки по СППВ является простота в применении. Однако ей присущи определенные недостатки, из-за которых она постепенно заменяется в США калифорнийской системой оценки по чистой энергии. Недостаток системы оценки по СППВ заключается в том, что учитываются потери энергии только в процессе переваривания и не берутся во внимание потери энергии с мочой, газами и теплопродукцией. Кроме того, не отражается полная картина действительного обмена энергии в организме, поскольку расчеты производятся только на основе анализов химического состава кормов; завышается питательность грубых кормов по сравнению с концентратами в рационах с высоким содержанием клетчатки, что связано с отсутствием учета повышенных потерь энергии на теплопродукцию.
Оценка питательности в овсяных кормовых единицах. Питательность кормов и нормы кормления животных должны быть выражены в одинаковых единицах, В нашей стране общую питательность кормов в соответствии с ОСТ-ВКС-6333 от 24 декабря 1933 г. оценивают в кормовых единицах. Одна кормовая единица по продуктивному действию (жироотложение у взрослого вола) эквивалентна продуктивному действию I кг овса, скормленного сверх сбалансированного рациона, достаточного для поддержания жизни. При этом считается, что в теле вола должно отложиться 150 г жира. Эта величина является результирующей от использования в организме всех органических веществ потребленного корма. В связи с этим питательность корма, выраженную в кормовых единицах, принято называть общей питательностью.
Общую питательность корма определяют расчетным путем на основании данных о химическом составе {содержание сырого протеина, сырой клетчатки, сырого жира и безазотистых экстрактивных веществ); переваримости этих веществ; показателей продуктивного действия переваримых питательных веществ; величины снижении питательности корма в зависимости от содержания в нем сырой клетчатки в грубых и зеленых кормах или коэффициентов полноценности и кормах, содержащих незначительное количество клетчатки.
После определения содержания к корме переваримых питательных веществ на основе эксперимента или табличных данных рассчитывают продуктивное действие непереваримых питательных веществ, содержащихся и 1 кг корма, умножай количество последних на показатели продуктивного действия отдельных переваримых питательных веществ (табл. 1).
Как уже отмечалось, еще Кельнером было установлено, что на переваривание сырой клетчатки животное затрачивает значительное количество энергии, высвобождающейся при использовании других переваримых питательных веществ, и в связи г этим при увеличении содержания в корме сырой клетчатки продуктивное действие снижается. Это положение учтено и методике определения общей питательности кормов в кормовых единицах.
Таблица 1. Продуктивное действие 1 кг переваримых питательных веществ, корм. ед.


Общая питательность сена, соломы, силоса, сенажа и зеленых кормов, содержащих 16% и более сырой клетчатки, а также зеленых кормов с содержанием 14, 12, 10, 8 и 6% сырой клетчатки, снижается соответственно на 0,97; 0,88; 0,80; 0,72; 0,63 и 0,57 кормовой единицы в расчете на 1 кг сырой клетчатки.
Общую питательность концентрированных кормов, корнеклубнеплодов и побочных продуктов перерабатывающей промышленности определяют аналогичным образом, то есть по сумме продуктивного действия содержащихся в них переваримых питательных веществ, умножением суммарного показателя питательности на коэффициент полноценности корма (табл, II).
Расчет питательности кормов в обменной энергии. В соответствии с принятыми Нормами кормления (1985 г.) в нашей стране в качестве основного показателя энергетической питательности кормов и рационов используют содержание обменной энергии, выраженной в мегаджоулях (МДж), в единице натурального корма и сухого вещества корма или рациона. Содержание обменной энергии устанавливают в опытах по определению переваримости питательных веществ по разности содержания энергии в принятом корме и выделенной в кале и моче (для жвачных и лошадей). В респирациопных опытах учитывают энергию газов, образующихся в желудочно-кишечном тракте, или определяют расчетным путем на основании данных о химическом составе и переваримости питательных веществ е помощью соответствующих уравнений регрессии:
для крупного рогатого скота
Оценка питательности кормов по чистой энергии. Эта система разработана Лофгрином и Гарреттом (1968) для растущего и откармливаемого крупного рогатого скота и Реттреем (1973) для растущих овец. Система предусматривает деление чистой энергии кормов на чистую энергию поддержания жизни и чистую энергию продукции.
Потребность в чистой энергии на поддержание жизни (НЭпж) приравнивается к теплопродукции животного в голодном состоянии. Для растущего крупного рогатого скота величина НЭпж, определенная экспериментальным путем, составляет 77 ккал (322 кДж), для овец — 63 ккал (264 кДж) на 1 юг обменной массы тела (МТ0'75), Чтобы определить общую потребность в чистой энергии на поддержание жизни, необходимо умножить эти величины на обменную массу тела, то есть живую массу тела взять в степени 0,75.
При получении экспериментальных данных о величинах теплопродукции и отложения энергии в приросте тела методикой предусмотрен анализ тела животного в начале и конце эксперимента методом сравнительного убоя и определение энергии в приросте. Недостатками этой (калифорнийской) системы оценки кормов по чистой энергии являются ее применимость только для растущего крупного рогатого скота и овец и довольно условное разделение энергетической ценности корма на чистую энергию для поддержания жизни и чистую энергию на продукцию.
Система оценки кормов по обменной энергии (британская Система Блэкстера, 1965). Эта система разработана Блэкстером Для жвачных животных и принята Сельскохозяйственным научно-исследовательским советом Великобритании в 1965 г. Согласно этой системе потребность животных в энергии выражается в форме обменной энергии, а эффективность ее использования зависит от живой массы, продуктивности животного и концентрации обменной энергии в 1 кг сухого вещества рациона. Концентрация обменной энергии в сухом веществе кормов — характерный показатель системы Блэкстера, влияющей на эффективность использования обменной энергии для поддержания жизни и образования продукции (табл. 12).
Чем выше концентрация обменной энергии, тем выше эффективность ее использования,
Зоотехническая наука и практика располагают огромными Материалами, свидетельствующими о неодинаковой питательной ценности протеина разных кормов. На протяжении последних 50—60 Лет предложено много биологических и химических методов определения качества протеина. Понятие о биологической ценности протеина было впервые введено Томасом (1909) и позднее -развито и широко использовалось Митчеллом (1924, 1944). Метод Томаса — Митчелла (процент всосавшегося азота, который может быть использован на поддержание жизни и рост) основан на знании баланса азота в организме животных, который определяют по формуле
Азот корма — (Пищевой азот кала +Пищевой азот мочи) Азот корма Пищевой азот кала
Этим методом была определена биологическая ценность протеина многих кормов. Данные (И. С. Попов, 1957), полученные на свиньях, показывают, что биологическая ценность протеина молока равна 84—95, казеина молока — 78—82, рыбной муки — 74, ячменя —71, соевого шрота — 67, льняного шрота — 61, люпина — 55, картофеля — 73, кукурузы — 54.
Недостаток метода Томаса — Митчелла — его сложность и то, что, он предполагает существование двух независимых друг от друга форм азотистого обмена в организме — экзогенного (распад кормового протеина) и эндогенного (распад тканевых белков). При этом принимают, что эндогенный обмен не изменяется при разных условиях кормления. И.С. Попов (1960) подверг критике эти положения и убедительно показал неверность представления о двух независимых формах протеинового катаболизма, постоянство же эндогенного азотистого катаболизма не подтвердилось в последующих экспериментах.
Для характеристики питательной ценности протеина Всесоюзным научно-исследовательским институтом животноводства предложены коэффициент использования переваримого протеина (КИПП), или коэффициент физиологически полезного протеина (КФПП).
ВОПРОС №3 КОЭФФИЦИЭНТ ПЕРЕВАРИМОСТИ И СПОСОБЫ ЕГО ВЫЧИСЛЕНИЯ
В зоотехнической литературе широко описан метод, основанный на определении коэффициента использования протеина (КИП), предложенный Осборном и др. (1919). Согласно этому методу, прирост живой массы в граммах делят на потребленный протеин в граммах при сохранении стандартных условий кормления и содержания животных. На величину КИП оказывают влияние уровень протеина, жира, качество жира в рационе, возраст, порода и пол животных. Недостаток метода Осборна — ошибочное положение о том, что прирост имеет постоянный состав, а потребленный протеин используется только для роста.
Довольно широко применяется оценка качества протеина по методике коэффициента нетто-протеина (КНП —NP), разработанной Бендером и Доеллом (1957). Значение КНП рассчитывают путем добавления потерь массы тела животных отрицательной контрольной группы к приросту животных опытной группы и деления на протеин, потребленный опытной группой. По данным Моррисона (1964), значение КНП менее изменчиво, чем кип.
Бендером и Миллером (1953, 1955) предложен метод определения качества протеина по данным анализа тут. Отношение отложенного азота к потребленному в процентах было названо использованием нетто (чистого) протеина (ИИП-РИ). Предложены и другие биологические методы определения качества протеина: по восстановлению белков печени (Генри и др., 1961), по активности ферментов печени (Аллисон, 1955).
Один из методов определения качества протеина для моно-гастричных животных — определение коэффициента свободных аминокислот плазмы кропи (КАП-РАА). Этот коэффициент рассчитывают делением содержания свободных аминокислот плазмы крови на относительную потребность в них животных. Аминокислоты с меньшим значением коэффициентов определяются как лимитирующие, то есть дефицитные, например лизин в глютене, триптофан в желатине, аргинин а казеине.
Для оценки питательности протеина по его аминокислотному составу предложены и химические методы, основанные на хроматографических и микробиологических анализах. Блок и Митчелл (1946) предложили определять питательную ценность протеина на основании сравнения его аминокислотного состава с аминокислотным составом белков яйца. Во многих случаях была подтверждена высокая степень корреляции между показателями питательной ценности протеина, установленная химическим методом, и в опытах на животных. Мак-Лауглаиом и др. (1963) был разработан упрощенный химический метод, основанный на отношении содержания лизина, метионина и цистина в протеине кормов к содержанию их в полноценном белке яйца.
По методу «индекс незаменимых аминокислот» Озера и др. (1960) питательную ценность протеина определяют как отношение геометрического среднего значения незаменимых аминокислот в оцениваемом протеине к соответствующему значению незаменимых аминокислот белков яйца.
Потребление азота
Анализ всех приведенных выше методов оценки питательной ценности протеина показывает, что главным их недостатком является то, что практически нее они не учитывают степень доступности аминокислот для организма.
Многие из них разрабатывались применительно к оценке пищевых продуктов, проведены на лабораторных животных (крысах) и поэтому их затруднительно использовать в практике кормления сельскохозяйственных животных. Требуется дальнейшее совершенствование методов оценки питательности протеина аминокислотном уровне с учетом доступности аминокислот и применимости этих методов в практике.
Система оценки кормов для крупного рогатого скота по сырому и переваримому протеину не учитывает природы содержащихся в корме азотистых веществ и особенностей превращения их в преджелудках, что в значительной мере влияет на количество протеина, поступающего в кишечник, и эффективность его использовании.
В преджелудках происходят значительные количественные и качественные преобразования азотсодержащих веществ корма. Протеин в преджелудках не только разрушается, но и синтезируется микрофлорой. Видимая переваримость протеина и поступление его, и тонкий кишечник зависят от того, какой m утих процессов превалирует. С увеличением доли растворимых фракций протеина в рационе видимая переваримость его возрастает вследствие быстрого расщеплении и всасывания значительных количеств аммиака в рубце. При этом поступление протеина в тонкий кишечник уменьшается и снижается истинная обеспеченность им животного.
Чем выше растворимость и следовательно, расщепляемость протеина в рубцовой жидкости, тем больше разница между видимо переваримым протеином, рассчитываемым по разнице между потребленным с кормом и выделенным с калом, и протеином, доступным для усвоения животным (всасывающемся в тонком кишечнике).
Новые системы оценки протеинового питания жвачных животных, принятые в ряде стран (США, Англия, Франция), основаны на учете, всасывающегося в тонком кишечнике протеина корма, избежавшего разрушения в преджелудках, и микробного протеина, синтезированного в рубце.
По этим системам для оценки количественных характеристик поступлении аминокислот во внутреннюю среду организма из указанных двух источников необходимо иметь сведения о количестве сброженного в преджелудках органического вещества, содержании в корме обменной энергии, количестве потребления протеина, его растворимости и расщепляемости.
В нашей стране в связи с выходом детализированных норм кормления сельскохозяйственных животных. Предусматривающих балансирование рационов для разных видов животных по 28—32 показателям, оценка состава и питательности кормов проводится по содержанию в них: кормовых единиц, обменной энергии, сухого вещества, сырого протеина, в том числе переваримого, сырого жира, сырой клетчатки, безазотистых экстрактивных веществ, в том числе крахмала и сахара, аминокислот (лизин, метионин + цистин), макроэлементов (кальций, фосфор, магний, калий, натрий, хлор, сера), микроэлементов (железо, медь, цинк, марганец, кобальт, йод), витаминов (каротин, витамины A, D, Е, Вь В2, Вз, В4, Be, В.8)
ВОПРОС №4 СИЛОСОВАННЫЙ КОРМ, ЕГО ВЛИЯНИЕ НА ПРОДУКТИВНОСТЬ. ОСНОВНЫЕ ТЕХНОЛОГИЧЕСКИЕ УСЛОВИЯ ПРИГОТОВЛЕНИЯ ВЫСОКОКАЧЕСТВЕННОГО СИЛОСА
Силос — это сочный корм, полученный в результате консервирования зеленых растений молочной кислотой. Силос хорошего качества охотно поедается всеми видами сельскохозяйственных животных.
Силосование зеленых кормов сопровождается меньшими потерями питательных веществ, в частности протеина (белка), чем при сушке на сено. Если при обычных условиях уборки на сено из зеленой травы теряется до 30% и более питательных веществ, то при правильно проведенном силосовании в хороших силосных сооружениях потери в общей питательности редко достигают 10%, а в белке близки к нулю. Белки в процессе силосования распадаются частично на пептиды и аминокислоты, но это не существенно снижает их питательность.
Силосование дает возможность заготавливать сравнительно дешевый сочный корм на зимний период, а в засушливых районах — и на летние месяцы при недостатке пастбищного корма; позволяет возделывать такие кормовые культуры, которые дают наивысший урожай, и убирать их независимо от погоды в наиболее удобное для хозяйства время; дает возможность широко пользоваться пожнивными и промежуточными культурами, а также хорошо использовать осенью отаву, которую не удается высушить на сено; позволяет использовать на корм сорняки и грубое разнотравье, из которых при сушке получается плохое сено, а при силосовании — вполне удовлетворительный сочный корм.
В настоящее время трудно представить зимние рационы животных без силоса. Силос повышает аппетит животных, улучшает пищеварение, обеспечивает потребность животных в витаминах и минеральных веществах. В значительной мере этим качествам способствует специфический вкус и запах силоса, образующийся в процессе сложных биохимических превращений белка и углеводов силосуемой массы и напоминающий запах квашеной капусты и других овощей, хлебного кваса и свежевыпеченного хлеба.
Основное преимущество силосования состоит в том, что доброкачественный силос по своей питательности и биологической ценности почти не отличается от зеленой травы. В силосованном корме количество протеина, жира, клетчатки, минеральных веществ и каротина почти не изменяется. Уменьшается лишь содержание сахара на 60-90%, который расходуется на образование органических кислот, главным образом, молочной кислоты. Органические кислоты по своим энергетическим свойствам незначительно уступают простым сахарам и легко усваиваются организмом животного. Например, уксусная кислота, накапливающаяся в процессе силосования, необходима для образования молочного жира. В целом силос высокого качества оказывает положительное влияние на молочную продуктивность коров. Переваримость основных питательных веществ силоса по сравнению со свежескошенной травой изменяется незначительно.
Научные основы силосования кормов. Сущность силосования заключается в том, что в свежей растительной массе, плотно уложенной в непроницаемые для воздуха силосные сооружения, в результате биохимических процессов постепенно накапливаются органические кислоты, преимущественно молочная, которые служат консервирующим средством, предохраняя, при известной концентрации, растительную массу от дальнейшего разложения и порчи. Поэтому основная задача правильного силосования сводится, главным образом, к выработке в силосуемой массе необходимого минимума молочной кислоты.
Биохимические процессы вызываются, с одной стороны, действием ферментов растительных клеток, а с другой — разнообразными микроорганизмами, попадающими в силос с зеленой травой.
Первым показателем изменения, происходящего в зеленой массе, сложенной в силосные сооружения, является повышение температуры, которое обусловливается дыханием растительных клеток и процессами брожения, идущими в силосуемой массе. В свежескошенной траве, уложенной в силосохранилище, растительные клетки некоторое время остаются живыми и продолжают дышать. Кислород, необходимый для нормального дыхания, они заимствуют из воздуха, остающегося в силосе. Чем больше воздуха остается в силосе, тем энергичнее идут окислительные процессы, тем сильнее разогревается силосуемая масса и тем выше в ней потери питательных веществ. На дыхание клеток расходуются, главным образом, углеводы, при этом конечным продуктом окислительных процессов являются углекислота и вода.
Наряду с изменениями, происходящими в результате дыхания клеток, в силосуемой массе быстро развиваются и бактериальные процессы. Силосуемая зеленая масса обычно очень богата разнообразными видами бактерий, вызывающими брожение. Из многочисленных видов брожения — молочнокислого, уксуснокислого, маслянокислого — для успешного силосования желательно молочнокислое, в результате которого накапливается молочная кислота. Накопление уксусной и масляной кислот, продуктов гниения белка и тем более плесени ухудшает качество силосованного корма.
Для своего развития молочнокислые бактерии требуют влажной среды и достаточного количества питательных веществ в форме Сахаров; в кислороде они не нуждаются, предпочитая анаэробные условия (без кислорода воздуха). При сбраживании Сахаров в качестве основного продукта они образуют молочную кислоту.
Очень важной особенностью молочнокислых бактерий является их способность развиваться в кислой среде, в которой невозможна жизнедеятельность маслянокислых и гнилостных бактерий.
Плесени и бактерии уксуснокислого брожения являются строго аэробными организмами, развивающимися лишь при свободном доступе кислорода воздуха. Поэтому для успеха силосования необходимо создать условия, благоприятные для развития в силосуемой массе молочнокислого брожения.
Богатство силосуемой массы растворимыми углеводами (сахаром) при содержании воды около 70%, анаэробные (без кислорода воздуха) условия и слабое нагревание благоприятствуют росту молочнокислых бактерий. В такой среде они сразу же получают преобладание над другими бактериями и усиливают его по мере выработки молочной кислоты, пока, наконец, и сами не погибают от нее. Процесс силосования практически заканчивается, когда кислотность (рН) достигает 4,0-4,2.
Для того чтобы в короткий срок в силосуемой массе накопилась молочная кислота, а кислотность повысилась до 4,2, необходимо содержание определенного количества сахара, которое получило название сахарного минимума, под которым подразумевается содержание сахара, необходимое для накопления в силосуемой массе молочной кислоты в количестве, достаточном для смещения рН силоса до 4,2. От соотношения сахарного минимума к фактическому содержанию сахара в траве зависит силосуемость растительной массы.
В зависимости от сахарного минимума и фактического содержания сахара в растениях они делятся на легкосилосующиеся, трудносилосующиеся и несилосующиеся.
Хорошо силосуются все злаковые растения, кукуруза, подсолнечник, овес, сорго, суданская трава, райграс, вико-овсяная и горохо-овсяная и другие злаково-бобовые смеси, капуста, ботва корнеплодов и др. У этих растений сахарный минимум полностью обеспечивается фактическим содержанием сахара.
Трудно силосуются бобовые растения в чистом виде: вика, горох, клевер, люцерна, донник, могар и другие, у которых содержание сахара не обеспечивает полностью сахарный минимум. Поэтому эти растения смешивают с хорошо силосуемой зеленой массой.
Не силосуются в чистом виде молодая люцерна в период бутонизации, крапива, ботва картофеля, арбузы, тыквы, многие сорняки. Зеленая масса этих растений содержит явно недостаточное количество сахара для образования необходимой концентрации молочной кислоты для сохранения корма.
Следует отметить, что содержание сахара и сахарный минимум растений могут значительно изменяться в зависимости от фазы вегетации, дозы, вносимых в почву удобрений, времени уборки и др. Поэтому силосуемость трав необходимо определять в каждом отдельном случае. В среднем содержание сахара в силосуемой зеленой массе при натуральной влажности должно быть не ниже 1,5%.
Факторы, влияющие на ход силосования и качество силоса. Для нормального хода молочнокислого брожения в силосуемой массе должна быть определенная концентрация сахара и других веществ в соке. Если влажность силосуемой массы очень высокая, создается опасность сильного разбавления сахара, если масса закладывается сухой — попадает много воздуха. Оптимальной влажностью является 65-75%. При такой влажности происходят меньшие потери питательных веществ и получается более качественный силос. Более влажная масса дает много сока, в котором оказывается протеин и другие вещества, нейтрализующие молочную кислоту, что приводит к неправильному брожению. Если зеленая масса имеет повышенную влажность (более 75%), то ее нужно силосовать в смеси с более сухими кормами или провяливать траву. Крупностебельные растения не провяливают, а понижают влажность путем смешивания с сухой измельченной соломой. Если приходиться силосовать более сухую массу с влажностью ниже 65%, то необходимо ее смешивать с более влажной травой или, в крайнем случае, добавлять воду. При неурегулированной чрезмерно высокой влажности исходного силосуемого сырья в результате брожения накапливается много уксусной кислоты, аммиака, появляется масляная кислота, такой силос приобретает резкий кислый вкус и запах, что значительно снижает его качество и поедаемость животными.
ВОПРОС №5 ИЗМЕНЕНИЕ ПИТАТЕЛЬНОЙ ЦЕННОСТИ КОРМА ПОД ВОЗДЕЙСТВИЕМ РАЗЛИЧНЫХ МЕТОДОВ ПОДГОТОВКИ ЕГО К СКАРМЛИВАНИЮ
Знание питательных свойств кормов позволяет фермеру предвидеть эффективность их использования для поддержания нормального состояния здоровья животных и получения высокой продуктивности, с одной стороны, и с другой — обоснованно заменять один корм другим в рационах, не изменяя обеспечения потребностей животных в тех или иных питательных веществах. Кроме того, в фермерских хозяйствах ежегодно приходится определять объем заготавливаемых кормов на одну голову и на стадо животных в целом. В этом случае питательную ценность кормов, которая характеризуется химическим составом, переваримостью и усвоением веществ животными должна быть определена каким-либо одним показателем. Это дает возможность спланировать в хозяйстве потребность животных в отдельных кормах и определить площади пахотной земли для производства кормов.
ФАКТОРЫ, ВЛИЯЮЩИЕ НА СОСТАВ И ПИТАТЕЛЬНОСТЬ КОРМОВ
Корма растительного и животного происхождения по своему составу и питательной ценности очень разнообразны. Так, в зеленых кормах изменчивость в содержании протеина достигает 15—20%, в корнеклубнеплодах— 13—25 и в зерновых — 8—16%. Значительно большей вариабельностью отличаются показатели содержания сырого жира и сырой клетчатки.
Изменения химического состава и питательной ценности кормов в зависимости от вида, сорта, фазы вегетации и условии выращивания растений. В процессе вегетации независимо от вида растения в нем отмечается накопление сухого вещества, увеличение количества БЭВ и клетчатки (при этом их соотношение изменяется в сторону клетчатки), снижение уровня сырого протеина и повышение в нем доли белка.
При этом переваримость отдельных питательных веществ в них снижается. В частности, содержание переваримого протеина (г в 1 кг сухого вещества) в зеленой кукурузе в период от кущения до восковой спелости снижается вдвое (с 88,8 до 43); в горохе этот показатель возрастает с 172,2 до 230,8 в период до бутонизации, а затем резко снижается до 146,6 во время цветения, образования бобов (115) и в период молочной спелости зерна (108,5).
Химический состав и питательная ценность кормовых растений в определенной степени обусловлены их сортовой принадлежностью, однако эти различия в большей или меньшей степени проявляются в зависимости от конкретных условий выращивания растений, поскольку различные сорта характеризуются разной отзывчивостью на изменение уровня питания. Установлено, что азотистые удобрения повышают содержание протеина в суданской траве, овсе, кукурузе и почти никакого влияния не оказывают на уровень жира и клетчатки (табл. 34).
Оптимизация водного режима растений путем орошения также оказывает определенное воздействие на их химический состав, причем доля влияния условий питания растений на химический состав наиболее сильно проявляется на ранних фазах вегетации, в период интенсивного роста растений.
На химическом составе растений отражаются природно-климатические условия, в частности световой и температурный режимы. Так, растения, выращенные в разных географических зонах, различаются по содержанию протеина. Отмечена общая закономерность—повышение содержания протеина в растениях при продвижении их с севера на юг и с запада на восток. Одновременно наблюдается уменьшение доли водорастворимого белка в результате повышения солерастворимого у бобовых и щелочерастворимого у злаковых. Аналогичные изменения отмечаются в засушливые годы.
Установлена (Г.В. Пикулевский, К.П. Карташов и С.Л. Иванов) зависимость химического состава масел от климатических условий произрастания растений. На основании этих данных была сформулирована климатическая теория, согласно которой тропический климат с равномерной, без резких колебаний температурой благоприятствует образованию в семенах глицеридов насыщенных кислот и олеиновой кислоты. Континентальный климат с резкими колебаниями температуры способствует образованию глицеридов полиненасыщенных кислот и задерживает синтез глицеридов олеиновой кислоты. В начале созревания в наиболее жаркие месяцы года (июнь, июль) в семенах льна образуются главным образом глицериды олеиновой и линолевой кислот; йодное число масла, полученного из таких семян, равно 120—150. В августе, к концу созревания семян, когда температура ночью ниже, чем днем, в семенах иде1 интенсивное накопление глицеридов линолевой кислоты, йодное число масла повышается до 170.
Кроме климатических условий, на запальные особенности в химическом составе кормовых растений существенное влияние оказывают характер почв и способ их обработки. Особо следует остановиться на изменчивости минерального состава зеленых растений, что наглядно демонстрируется данными, приведенными в табл. 35.
Значительные колебания минерального состава растений, как и других составных компонентов, обусловливаются многими факторами, среди которых наиболее существенны видовые различия и изменение минерального состава по фазам вегетации. Колебания в содержании минеральных веществ в растениях разных видов даже внутри одного семейства, в 2—3 раза превышающие минимальное значение, следует считать обычным явлением. В связи с этим любые табличные данные о минеральном составе кормов следует считать лишь ориентировочными, требующими уточнения применительно к конкретным условиям.
Климат и метеорологические условия года оказывают влияние на формирование химического состава растений. В годы с оптимальным количеством и равномерным распределением осадков в период вегетации в растениях накапливается больше минеральных веществ, чем в засушливые годы. Безусловно, на минеральном составе растений сказывается и внесение удобрений. Однако предугадать, как изменится содержание тех или иных элементов в растении при внесении удобрений, пока не представляется возможным. При этом обычно проявляются и видовая специфичность растений, и агрофона, и почва, и метеорологические условия года, и ряд пока неизвестных и неучитываемых факторов.
Что касается микроэлементов, то на их содержание в растениях меньшее влияние оказывают агротехнические приемы, погодные условия, видовая и сортовая принадлежность. Естественное накопление микроэлементов в растениях разных семейств, а также в разных частях растения (корень, стебель, листья, плоды, семена) значительно различается, но основным фактором, определяющим изменчивость микроминерального состава золы растений, является место их произрастания. Работами В. И. Вернадского и его последователей доказано, что микроэлементы, как и другие минеральные вещества, распространены на земле неравномерно. Это зависит от 'неодинакового состава почвообразующих пород и от особенностей почвообразовательного процесса. Недостаток или избыток микроэлементов в почве в основном и обусловливает содержание их в растениях.
Развивая идеи В.И. Вернадского, академик А.П. Виноградов разработал учение о биогеохимических провинциях. Биогеохимические провинции — это ограниченные области земного пространства, имеющие в среде определенную, только им свойственную концентрацию одного или нескольких минеральных элементов. Эти концентрации могут быть избыточными или недостаточными. Такие провинции принято называть эндемическими. На территории СССР изучены и изучаются многие биогеохимические провинции, богатые .или бедные кобальтом, медью, йодом, бором, молибденом и другими элементами. Обобщив материал этих исследований, В. В. Ковальский (19G0) составил картограмму биогеохимических зон и провинций СССР, дающую представление о выявленных районах с избыточным и недостаточным содержанием в почвах, а следовательно, в кормах различных минеральных элементов.
Изменения химического состава и питательной ценности кормов в зависимости от технологии их приготовления и условий хранения. Эти изменения связаны, прежде всего, с темп биохимическими преобразованиями, которые происходят в скошенном растении в процессе сто консервации. При заготовке различных видов кормов отмечаются биохимические потери питательных веществ в процессе дыхания в тканях скошенных растений. В результате часть моно- и дисахаридов окисляются до диоксида углерода и воды, то есть практически теряются. Чем продолжительнее время между скашиванием и отмиранием (консервацией растений), тем большее количество углеводов теряется и ниже питательная ценность готового корма.
ВОПРОС №6 ОПИСАТЬ ТЕХНОЛОГИЮ ПРИГОТОВЛЕНИЯ СИЛОСА И СЕНАЖА В ХОЗЯИСТВЕ
Силосованные корма по своему химическому составу существенно отличаются от состава исходной массы. В этом случае легкосбраживаемые углеводы растений используются для синтеза молочной и уксусной кислот, служащих консервирующим началом в силосе. Изменение влажности исходного сырья (путем провяливания скошенных растений) снижает интенсивность бродильных процессов и способствует лучшей сохранности углеводов в процессе силосования корма. Наряду с углеводами большие потери при заготовке кормов наблюдаются в отношении жирорастворимых витаминов, в частности каротина, уровень которого может снижаться в несколько раз.
Во время скашивания и приготовления различных видов, консервированных кормов происходят существенные изменения в протеиновой питательности. Так, по данным В.Ф. Лемеша (1971), в процессе приготовления силоса заметно снижается содержание большинства аминокислот, кроме метионина, аланина и цистина, количество которых возрастает. Предварительное провяливание растений перед силосованием до содержания 32% сухого вещества улучшает сохранность всех незаменимых аминокислот.
Измельченность силосуемой массы. Степень измельчения силосуемых растений зависит от их влажности в момент укладки. При влажности 65% и ниже величина резки должна быть 2-3 см, при влажности 70-75% — 4-5 см, при влажности 80% — 8-10 см. Чем крупнее резка, тем меньше выделяется сока, меньше потери питательных веществ. В то же время подсушенная масса обеспечивает необходимое количество сока только при мелкой резке. Мелкотравянистую растительность с высокой влажностью лучше силосовать в неизмельченном виде. Соблюдая требования к длине резки, получают силос лучшего качества, с меньшими потерями. Огрубевшие, трудноуплотняемые растения необходимо измельчать до размера не более 2 см.
Сроки закладки силосуемой массы. При силосовании зеленых кормов важным технологическим приемом является быстрая загрузка массы в хранилище и тщательное его укрытие. Закладка силоса в крупнотоннажные силосные сооружения должна продолжаться не более 3-4 дней, а укрытие массы при хорошей трамбовке должно быть выполнено в первые сутки после окончания загрузки. Срок закладки массы в малообъемные хранилища (до 500 т) не должен превышать 2 дней.
При несоблюдении сроков закладки в процессе дыхания клеток растений и развития микроорганизмов уже через несколько часов зеленая масса разогревается до 50-60'С и выше. Если закладку проводят медленно и слой уплотненной травы составляет не более 30 см в день, то масса разогревается до высоких температур. В это время теряются до 30-40% самых ценных питательных веществ (сахара, крахмала, белков, витаминов), содержание переваримого протеина в силосе снижается в 1,5-2 раза, белок становится труднопереваримым, а каротин остается в малом количестве.
В условиях высоких температур в силосе происходит взаимодействие аминокислот с сахаром, образуются стойкие непереваримые соединения, придающие перегретому силосу бурый или темно-коричневый цвет. Несмотря на то что бурый силос скот поедает охотно, питательность и полноценность его очень низкая, а следовательно, и продуктивность скота будет невысокой.
Силосные сооружения. Хороший силос с минимальными потерями можно получить лишь в силосохранилищах, если они правильно устроены.
Силосохранилища должны удовлетворять следующим требованиям:
1) должны быть непроницаемыми для воздуха: чем больше воздуха проникает в силос, тем значительнее потери питательных веществ в процессе брожения и тем труднее получить молочнокислое брожение;
2) должны быть достаточно глубокими, чтобы силосуемая масса плотно укладывалась под давлением собственного веса и меньше задерживала внутри себя воздух;
3) должны иметь строго отвесные и гладкие стены, с закругленными углами, чтобы оседание силосуемой массы шло правильно и равномерно, без образования пустот;
4) наземные силосохранилища (траншеи) должны быть достаточно прочными, чтобы выдержать боковое давление силосуемой массы и, кроме того, кислотоупорными и хорошо предохраняющими силос от промерзания.
Выбор силосных сооружений зависит прежде всего от количества одновременно закладываемой массы в течение не более 4 дней.
Хорошими силосными сооружениями являются траншеи. Их делают заглубленными в землю и наземными. Первые устраивают в плотном глинистом грунте глубиной 2,5-3,5 м, стенки имеют уклон ко дну, ширина по верху — не менее 9 м, а длина — в зависимости от потребности, но не более 30 м. Такие траншеи строят там, где грунтовые воды стоят низко, и в том случае, если от их высшего уровня до дна траншеи не менее 0,5 м.
Наземные траншеи имеют некоторые преимущества. Их можно устраивать в поле независимо от уровня грунтовых вод, затраты на них минимальны. В этом случае траншеи строят в виде двух параллельных стен из досок, брусьев, железобетона, сборных панелей. Ширина между стенами — 6-15 м, высота стен — 2,3-3 м, длина — не более 30-40 м. Основание должно быть замощено или бетонировано.
При одновременной закладке небольших количеств силосуемой массы нужны силосохранилища для получения силоса хорошего качества. В этом случае наилучшими являются башни диаметром 7-9 м, высотой 22 м.
Размер силосохранилищ в каждом конкретном хозяйстве обычно определяется потребностью в силосованном корме в зависимости от количества скота, суточных норм скармливания и продолжительности кормления силосом.
Для небольших объемов заготовки силоса простейшим силосохранилищем являются ямы. Их следует устраивать только в плотном грунте, на сухом возвышенном месте, с низким стоянием грунтовых вод. Лучшим грунтом считается глинистый. При ежегодном силосовании ямы следует облицовывать вязкой глиной, кирпичом, бетоном.
При вычислении вместимости силосных сооружений принимают, что масса 1 м3 готового силоса из кукурузы, подсолнечника, сорго и других злаковых трав составляет около 700 кг, из разнотравья, вико-овсяной смеси и других бобово-злаковых смесей — около 600 кг (табл. 65).
надо следить за равномерностью натяжения пленки, тщательно заправляя ее вдоль боковых стен и по краям.
Выпускаемая промышленностью пленка даже толщиной 200 мкм слабо противостоит воздействию солнечных лучей, перепадам температуры и ветру, и если ее не укрывать слоем земли (5-10 см), песка, опилок, торфа, то через 2-3 месяца она полностью разрушается и происходит разгерметизация корма. Укрытие пленки соломой без слоя извести нежелательно, так как мыши приводят пленку в полную негодность.
Во многих хозяйствах при отсутствии пленки силос хорошего уплотнения укрывают землей, глиной слоем 15-30 см. Этот способ имеет ряд неудобств, связанных с большими затратами труда, особенно при снятии укрытия зимой, к тому же силос сильно загрязняется землей. Поэтому с наступлением заморозков часть грунта снимают, а силос укрывают соломой.
Особенности технологии силосования отдельных видов растений. Состав и питательность силоса. Для приготовления силоса используют различные виды растений, среди которых наиболее широкое распространение получили кукуруза, подсолнечник, сорго, суданская трава, озимая рожь, злаково-бобовые смеси, зеленая масса естественных и сеяных кормовых угодий.
Кукуруза. Относится к легкосилосующимся растениям. Является широко распространенной силосной культурой и возделывается во многих районах нашей страны. При оптимальных условиях агротехники урожай зеленой массы составляет 350-600 ц/га, а на орошаемых землях — 1000-1200 ц/га. Кукуруза практически до конца вегетации накапливает питательные вещества. Например, если принять за единицу сбор кормовых единиц с гектара посева кукурузы в фазе цветения, то в период молочно-восковой спелости он составит 1,5, а при восковой спелости зерна — 1,8 раза больше.
По мере созревания кукурузы существенно меняется соотношение морфологических частей растения, увеличивается масса зерна в початках, что повышает энергетическую ценность корма. Например, в фазе выбрасывания метелки листья занимают 58%, стебли — 42%; в начале образования зерна: листья — 45%, стебли — 46% и початки — 9%; в молочную спелость зерна — соответственно 33, 28 и 39%; в молочно-восковой спелости — 31, 25 и 44% и в восковой спелости — 25, 29 и 46% . В фазе выбрасывания метелки в 1 кг зеленой массы кукурузы содержится 0,15-0,18 корм, ед., в фазу молочно-восковой спелости зерна — 0,31 корм. ед. С увеличением возраста кукурузы содержание каротина уменьшается. Если в период выбрасывания метелки в зеленой массе кукурузы содержится 40-50 мг/кг каротина, то при молочно-восковой спелости зерна — только 15-20 мг/кг.
Поэтому на силос кукурузу целесообразно убирать в фазе молочно-восковой спелости зерна. В этот период влажность зеленой массы колеблется от 65 до 78%.
Кукуруза является теплолюбивой культурой, не выдерживает заморозков. Поврежденную заморозками кукурузу следует немедленно убирать на силос, так как подмерзшие листья быстро подсыхают, в результате чего теряется значительная часть протеина и каротина. По этой причине уборку кукурузы в северных районах страны начинают в более ранние фазы вегетации — молочной и молочно-восковой спелости зерна.
При силосовании зеленой кукурузы высокой влажности (80-85% и выше) корм, как правило, получается перекисленным (рН 3,5-3,8), часто с наличием масляной кислоты. Потери питательных веществ от угара и утечки сока нередко достигают 30% и более. В этом случае для повышения качества силоса целесообразно добавлять 10-15% измельченной соломы.
Технология силосования заключается в следующем. На дно траншеи ямы) укладывают соломенную резку слоем 80-100 см, затем зеленую массу кукурузы слоем 30-40 см, потом солому слоем 10—20 см, на которую укладывают кукурузу слоем 30-40 см, и т. д. Верхний слой укладывают только из кукурузы. Массу хорошо уплотняют и изолируют от воздуха. Солома для силосования должна быть без признаков порчи.
В южных районах страны после уборки кукурузы на зерно оставшиеся листья и стебли измельчают и силосуют. В этом случае влажность массы составляет не выше 50-60%. При силосовании в чистом виде добавляют воды или силосуют совместно с высоковлажными кормовыми культурами — бахчевыми, отходами овощеводства, ботвой свеклы в количестве 15-20% по массе.
Надежным способом консервирования початков кукурузы является силосование их в фазе восковой спелости зерна. Силосовать початки необходимо сразу же после уборки, измельчив их до состояния мезги, и хранить в облицованных сооружениях с хорошей герметизацией.
Початки в фазе восковой и полной спелости хорошо силосовать с неогрубевшими бобовыми травами и с измельченными плодами кормовых бахчевых культур. Эти добавки улучшают поедаемость свиньями и птицей силосованных початков и повышают их биологическую ценность. Состав и питательность кукурузного силоса в среднем представлены в таблице 66.
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
1. Аликаев В.А. Справочник по контролю кормления и содержания животных. – М.: Колос, 1982.
2. Боярский Л.Г. Производство и использование кормов. – М.: Росагопромиздат, 1988.
3. Девяткин А.И. Рациональное использование кормов. – М.: Агропромиздат, 1990.
4. Попов И.С. Рациональное кормление свиней. – М.: Колос, 1975.
5. Хохрин С.Н. Корма и кормление животных: Учебное пособие. СПб.: Издательство «Лань», 2002.
Похожие рефераты:
Технология молока и молочных продуктов
Особенности оценки продуктивности сельскохозяйственных животных
Муха (diptera muscidae) как продуцент кормового белка для птиц на Востоке Казахстана
Механизмы снижения себестоимости производства молока
Кормление коров на сельскохозяйственном предприятии
Совершенствование организации производства молока в СПК
Премиксы и их использование в кормлении сельскохозяйственных животных и птицы
Анализ факторов, влияющих на молочную продуктивность коров
Технология кормления и содержания стельных и сухостойных коров
Повышение эффективности продукции скотоводства
Анализ организационно-экономических условий племзавода ОАО "Восточный"
Кормление поросят-сосунов и отъемышей
Особенности выращивания крупного рогатого скота, свиней и птицы