| Скачать .docx | Скачать .pdf |
Реферат: Химия нуклеиновых кислот
Реферат на тему:
Химия нуклеиновых кислот
2009
Начнем с ДНК. Ее физические характеристики и «конструктивные» особенности были достаточно подробно описаны во введении. Настало время подробнее познакомиться с нуклеотидами, уточнить понятие «комплементарное», конкретно познакомиться со слабыми («водородными») связями, удерживающими две цепи ДНК друг подле друга, и строением самих цепей.
Путем ферментативного гидролиза гигантские молекулы ДНК удается полностью разбить на составляющие звенья и разделить их с помощью уже упомянутого метода хроматографии. Физико-химический анализ показал, что эти звенья («нуклеотиды») имеют в своем составе (подобно аминокислотам) одинаковые линейные элементы, которые собственно говоря, и выстраивают цепь, а также боковые ветви («нуклеиновые основания»), которыми они отличаются. Таковых оказалось, действительно всего четыре: два относительно меньших размеров —цитозин (Ц) и тимин (Т) и два более крупных — аденин (А) и гуанин (Г), и основание урацил (У), которое в РНК заменяет тимин.
Менее крупные молекулы (Ц, Т, У) являются производными молекулы пиримидина и потому называются пиримидиновыми основаниями. Две более крупных молекулы, — производные пурина, — именуются пуриновыми основаниями,
Все эти нуклеиновые основания содержат гетероциклы с сопряженными связями. Такие молекулы обладают ярко выраженной гидрофобностью и, кроме того, сильно поглощают ультрафиолетовый свет вблизи максимума поглощения при длине волны 260 тц. (По интенсивности УФ-поглощения можно количественно определять содержание ДНК или РНК в водном растворе.) Все эти молекулы плоские и содержат активные атомы и группы (N, NH, NH3, О), способные образовывать водородные связи. Ковалентные связи всех нуклеиновых оснований с линейными частями звеньев ДНК и РНК осуществляются по NH-группам, окруженным пунктирами.
Теперь мы уже можем их назвать — это пары: А—Т и Г—Ц.
Заметим, что в паре Г—Ц три связи, а в А—Т только две. Мы вправе заключить, что первая пара связана прочнее. Анализ наружных («линейных») участков нуклеотидов ДНК подтвердил, что они содержат плоские молекулы сахара дезоксирибозы (у РНК — рибозы)
В пятичленных кольцах этих Сахаров находится по четыре атома углерода, пятый атом углерода расположен вне кольца. Следует обратить внимание на нумерацию углеродов от Г до 5' (штрихи при номерах роли не играют — они «пришли» из общепринятой химической номенклатуры). Обведенные пунктиром гидроксилы в обеих молекулах участвуют в образовании ковалентных связей с нуклеиновыми основаниями.
В линейную часть звена ДНК (и РНК) входит еще остаток ортофосфорной кислоты. На рис. 25 изображена в полном составе пара кодирующих звеньев ДНК. В изображении используются общепринятые упрощения: во всех свободных углах колец надо представить себе стоящие там атомы углерода, а на концах выходящих из этих углов черточек — атомы водорода. Еще раз обратите внимание на нумерацию углеродов в дезоксирибозах. Не случайно на рис. 25 правый нуклеотид (Т) изображен как бы «вниз головой» относительно левого нуклеотида (А). Как видно из рис. 26, такое относительное расположение имеет место во всех парах цепи ДНК. Оно диктуется необходимостью соответственного положения оснований (при котором сближены группы, образующие водородные связи) и сахаро-фосфатных связей между нуклеотидами каждой нити. Комплементарные нити ДНК оказываются противоположно направленными. Если, к примеру, мы примем за направление нити переход от 5'-углерода одной концевой молекулы дезоксирибозы, несущей фосфатную группу, к 3'-углероду другой концевой дезоксирибозы, несущей ОН-группу, то тем самым мы определим направление всей нити. Такое направление принято называть направлением от 5'-конца нити к ее 3'-концу или сокращению
|
|
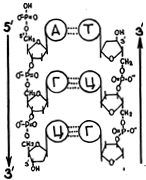
На рис. эти направления для обеих нитей указаны стрелками. Они противоположны. Отсюда следует, что переходя от звена к звену в двунитевой ДНК (т. е. от одной пары связанных нуклеиновых оснований к следующей паре) мы неизбежно будем двигаться по одной нити в направлении 5'-3', а по второй в направлении 3'-5'. Это обстоятельство следует запомнить.
На рис. для простоты нуклеиновые основания были обозначены кружками. Рис. 2 изображает полную структуру двухзвенного фрагмента ДНК. Штриховка означает, что плоскости оснований перпендикулярны направлению параллельных сахаро-фосфатных цепочек.
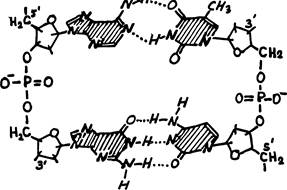
Рис. 2
Сопоставив суммарные элементарные составы пар А—Т и Г— Ц легко подсчитать, что они почти одинаковы. Если же учесть, что усреднение элементарного состава всей молекулы ДНК происходит по более, чем миллиону пар, можно с еще большей уверенностью, чем для белков, утверждать, что элементарный состав всех ДНК одинаков. Он оказался совсем иным, чем у белков (см. выше), а именно: 33,1% С; 38,2% О; 15,7% N и 9,3% Р.
Подробно рассмотрев состав и структуру ДНК, можно для РНК ограничиться лишь краткими замечаниями. С учетом того, что замена тимина на урацил в составе РНК ведет к отличию всего в двух атомах водорода, а дезоксирибозы на рибозу — в одном атоме кислорода, можно утверждать, что элементарный состав РНК такой же, как у ДНК. Главное отличие в том, что большинство РНК — однонитевые молекулы. Им ведь нет нужды реплицироваться. (Впрочем, последнее замечание сегодня, в 2001 году, звучит не столь категорически, как даже год назад, ввиду совсем недавнего обнаружения возможности репликации РНК с помощью фермента «рйбозима».)
Давно известно, что на отдельных небольших участках нить РНК может спариваться сама на себя, благодаря разнесенным по ее длине комплементарным участкам. Такое образование двунитевых участков, по-видимому, важно для структурирования и функционирования больших (например, рибосомальных) молекул РНК и малых молекул тРНК.
Наконец, следует иметь в виду, что растворы РНК, благодаря гибкой однонитевой структуре ее молекул, обладают гораздо меньшей вязкостью, чем растворы ДНК. Еще, в отличие от ДНК, РНК можно расцепить до нуклеотидов обработкой слабым щелочным раствором. Способность к поглощению света в УФ-области у обоих типов молекул нуклеиновых кислот одинакова.
Механизм редупликации ДНК
Здесь целесообразно лишь вкратце повторить основные представления о фундаментальных жизненных процессах: редупликации ДНК, транскрипции ее с переносом наследственной информации на иРНК и механизме трансляции — синтезе белка, направляемом этой информацией.
Разумеется, по ходу дела мы сделаем некоторые уточнения и добавим ряд деталей к описаниям названных процессов.
Редупликацию ДНК (копирование «материнской» ДНК при делении клетки) осуществляет особый фермент ДНК-полимераза. Посадке этого фермента на одну из нитей ДНК предшествует строго локализованный разрыв кольца, если ДНК кольцевая (у бактерий) и некоторое расплетание концевого участка ее гигантской двунитевой спирали. Заметим сразу, что ДНК-полимераза может садиться на любой из двух концов спирали, но обязательно на ту нить, для которой этот конец является 3'-концом (будь то «кодирующая» или «защитная» нить). Продвижение фермента вдоль «матрицы» материнской нити всегда идет в направлении от 3'-конца к 5'-концу. Отсюда следует, что синтезируемая по этой матрице, «комплементарная» к ней новая нить ДНК будет начинаться своим 5'-концом и наращиваться в направлении своего будущего 3'-конца. Эти два направления нельзя путать. В случае сомнения достаточно вспомнить, что наращивание новосинтезируемой нити происходит путем последовательного присоединения нуклеотидов, уже несущих фосфатную группу, связанную с 5'-уг-леродом дезоксирибозы. Следовательно к предыдущему, уже стоящему на своем месте нуклеотиду он должен присоединяться по его ОН-группе, связанной с 3'-углеродом дезоксирибозы. А это и означает, что наращивание новой нити ДНК идет в направлении б'-3' . Здесь же уместно напомнить, что работа продвижения ДНК-полимеразы осуществляется за счет энергии разрыва химической связи между первым и вторым фосфатами соответствующего нуклеозидтрифосфата — предшественника присоединяемого нуклеотида.
Теперь перейдем к добавлениям и уточнениям. Начнем с того, что в клетке кишечной палочки (E.coli) обнаружилась не одна, а целых три ДНК-полимеразы. Они заметно отличаются друг от друга по молекулярному весу и по числу молекул каждой из них, содержащихся в клетке. А также по их роли в процессе редупликации ДНК.
Исторически первой была обнаружена и очищена ДНК-поли-мераза I (фермент Корнберга). Потом появились ДНК-полимеразы II и III, Молекулярные веса этих трех ферментов, соответственно, 109, 90 и 300 тыс. дальтон, а их представительство в одной клетке: 300, 40 и 20 штук. Различие функций будет видно из дальнейшего.
Описание 1-го этапа редупликации начнем с того, что первоначальное расплетание конца двунитевой материнской молекулы ДНК осуществляется с помощью специального белка «топоизоме-разы». Он продвигается по двунитевой молекуле, ослабляя ее водородные связи настолько, что на пройденном им коротком концевом участке эти связи разрываются уже при температуре 37°С. Вслед за топоизомеразой на материнскую ДНК садится и начинает продвигаться по ней другой белок ДНК-геликаза, которому предстоит сыграть свою роль позже. Затем особая РНК-полимера-за, работающая только с конца нити ДНК, именуемая «праймаза» строит очень короткую цепочку рибонуклеотидов (получившую название «праймера») комплементарно к началу нити ДНК. У бактерий это всего 5 нуклеотидов, а у эукариотов — порядка 40. (На рис. 28 все праймеры показаны тонкой линией, а все нити ДНК — жирной.)
Только теперь, сразу за праймером на ту же нить ДНК (условимся, для простоты, называть ее «первой») садится ДНК-поли-мераза, которая может начать строительство комплементарной нити ДНК только начиная от праймера, присоединяясь к нему(«танцует от печки»). Это — ДНК-полимераза III, самая крупная, состоящая из 6-ти субъединиц и по своей функции главная — она будет вести «комплементарный синтез» ДНК по этой первой материнской нити ДНК до самого конца. Первоначальное движение этой ДНК-полимеразы ограничено 1-2-мя тысячами нуклеотидов первой нити (у эукариотов — всего на 200 нуклеотидов).
Вторая материнская нить (еще пустая) формирует вместе с первой нитью «вилку редупликации».
Между геликазой и ДНК-полимеразой III образуется некоторый участок обнаженной 1-й нити. 2-я нить тоже еще ничем не прикрыта. Эти две нити после ухода геликазы могут вновь сомкнуться. Чтобы этого не происходило, вплотную за геликазой на 1-ю нить садятся четыре, так называемых «ДНК-связывающих белка». Им не приписывают иных функций, кроме защиты от восстановления двойной спирали ДНК близ вершины вилки...
Каждому, кто пробовал просто раздвинуть концы двух скрученных в спираль веревок (противоположные концы которых закреплены) знает, что сначала это удается сделать легко, потом все труднее, а затем становится и невозможно. Это происходит потому, что концы веревок именно раздвигаются, а не раскручиваются. Остальная часть спирали при этом уплотняется, сохраняя прежнее число витков, что и создает напряжение, мешающее дальнейшему раздвижению концов. Достаточно теперь у вершины вилки перерезать одну веревку, как плотно скрученная ее часть начинает вращаться до тех пор, пока напряжение уплотнения витков не будет снято. Одновременно будет вращаться вокруг своей оси и не перерезанная ветвь раздвинутой веревки. Отрезанная ветвь, если она сохранила какое-то сцепление, хотя бы в одной точке, с другой ветвью, будет без напряжения вращаться вместе с нею...
Подобный описанному процесс, вероятно, осуществляется и в ходе репликации ДНК. Пройдя до вершины вилки разошедшихся нитей материнской ДНК 'тандем геликаза-ДНК-связывающие белки — ДНК-полимераза III останавливается (см. рис. 28). Топо-изомераза уходит дальше по двухнитевой материнской ДНК, а ге-ликаза разрывает сахаро-фосфатную связь на 2-й нити. Уплотненные на участке, прилегающем к вилке, витки двойной спирали расправляются, 1-я нить ДНК вместе с сидящими на ней белками вращается вокруг своей оси, а вокруг этой нити поворачивается и отрезанный кусок 2-й нити, временно связанный с геликазой. Этот кусок называют «фрагментом Оказаки» — по имени ученого, обнаружившего появление таких фрагментов при редупликации. После снятия напряжения нити двойной спирали материнской ДНК снова могут начать расходиться. Но до этого с отрезанного конца фрагмента Оказаки другая праймаза начинает на нем построение нового рибонуклеотидного праймера. Затем геликаза освобождает фрагмент и уходит вперед, а специальный фермент «лигаза» пришивает начало фрагмента Оказаки на прежнее место — ко 2-й нити материнской ДНК. Заметим, что лигаза (М=96 тыс.) в клетке E.coli представлена наиболее многочисленной популяцией — около 200 молекул. Из чего следует, что она выполняет не случайные «ремонтные» работы, а является полноправным членом совокупности ферментов, обеспечивающих редупликацию ДНК (подобно значению ниток для хирурга).
Когда праймер готов, впереди него, по направлению к 5'-кон-цу 2-й материнской нити ДНК садится ДНК-полимераза I. Начинается строительство нити, комплементарной к этому фрагменту 2-й нити, опять в направлении 3'— 5', считая по этой нити. ДНК-полимераза I доходит до конца фрагмента Оказаки и снимается. Этим заканчивается 1-й этап редупликации (рис. 28).
Между тем праймер, оставшийся в начале 1-й нити разрушается некой «рибонуклеазой Н», — ферментом, рвущим нить РНК, находящуюся в комплексе с нитью ДНК. На его место ДНК-полимераза II ставит «правильные дезоксирибонуклеотиды. В то же время топоизомераза, геликаза, а за ними и ДНК-полимераза III продвигаются вперед. Начинается 2-й этап редупликации. Вилка репликации тоже продвигается, прилегающий к ней участок материнской двунитевой ДНК уплотняется и весь синтезирующий тандем останавливается. Геликаза опять разрезает 2-ю нить, образуя второй фрагмент Оказаки. Так же, как раньше, на обрезанном (временно) конце фрагмента создается праймер, к нему «подсаживается» ДНК-полимераза I и начинает копировать второй фрагмент Оказаки, т.е. 2-ю нить материнской ДНК. Отличие второго этапа будет только в том, что на пути этой полимеразы встретится праймер, оставшийся от копирования 1-го фрагмента Оказаки. Но ДНК-полимераза I, в отличие от всех прочих ДНК-полимераз, обладает еще и 5'—3' экзонуклеазной активностью, т. е. в направлении своего движения. Она разрушает праймер и доходит до того
места, с которого начинала копирование 1-го фрагмента Оказаки ее предшественница. Остается только связать фосфодиэфирной связью эти два куска новосинтезированной комплементарной нити. Естественно, что это делает вездесущая ДНК-лигаза.
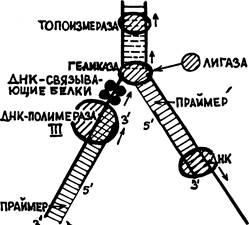
Тем временем в районе образования уже третьей вершины вилки редупликации происходят точно такие же события, как на 2-м этапе редупликации. Скорость этого процесса оценивается как, примерно, 1000 нуклеотидов в секунду у бактерий 100 — у животных и 20 — у растений. '
Весьма вероятно, что в то же самое время аналогичные процессы расплетания двойной спирали с образованием фрагментов Оказаки и комплементарного построения новых нитей ДНК идут и с противоположного конца материнской ДНК. Разумеется, там ДНК-полимераза III непрерывно двигается вдоль той нити ДНК, которую мы назвали 2-й, а на фрагменты Оказаки разрезается 1-я нить. Когда два движения встречаются, две дочерние копии исходной ДНК оказываются готовы. (Их «сошьет» все та же лига-за.) Кстати оказалось, что длина фрагментов Оказаки у E.coli (1—2 тысячи нуклеотидов) значительно больше, чем у эукариотов (меньше 200). Не лишено интереса совпадение этой последней цифры с длиной ДНК в нуклеосоме.
Конечно, все описанные механизмы были установлены для E.coli и подтверждены для других микроорганизмов. Имела ли смысл вся эта огромная работа, с точки зрения познания процессов передачи наследственной информации у высших организмов? Безусловно имела! Во-первых, исследователи не раз убеждались в том, что фундаментальные реакции на разных уровнях развития организмов протекают сходным образом. Во-вторых, ферменты, выделенные из бактерий, хорошо ведут in vitro редупликацию ДНК, полученных из любых источников, и потому представляют собой прекрасный инструмент для исследования этих ДНК.
Что до механизма редупликации у эукариотов, то, хотя он изучен хуже, тем не менее и здесь найдены и охарактеризованы целых пять ДНК-полимераз, которые принято обозначать греческими буквами: ос, Р, у, 5 и е. Основным ферментом, подобным ДНК-полимеразе III у бактерий, является ДНК-полимераза 5. ДНК-полимераза а отвечает за построение праймеров (из рибонуклеотидов). ДНК-полимераза Р — копирует фрагменты Оказаки и отвечает за репарацию ДНК. ДНК-полимераза у ведет синтез ДНК в митохондриях. Функция ДНК-полимеразы с пока неизвестна.
Конечно, гигантские ДНК высших организмов начинают редупликацию не только с концов молекулы, но и во множестве промежуточных точек (как — непонятно). Считают, что у дрожжей таких точек начала репликации около 300. Они отстоят друг от друга на 40 тысяч пар нуклеотидов. В ДНК человека насчитывают до 20 000 точек начала, расположенных с интервалом в 150 тысяч пар нуклеотидов. По-видимому, местами начала расплетания и посадки ДНК-полимеразы 5 служат последовательности относительно слабо связанных А—Т пар оснований.
Отметим еще, что все ДНК-полимеразы, ведущие комплеметарный синтез, как у бактерий, так и у эукариотов, обладают еще и экзонуклеазной активностью в направлении 3'-5', — обратном направлению синтеза. Они способны как бы «обернуться» и удалить только что ими же присоединенный нуклеотид. Это — очень важный механизм устранения ошибок в комплементарном синтезе. Ошибку ведь можно обнаружить только тогда, когда она уже сделана. И немедленно исправить! Это «умеют» делать ДНК-полимеразы. Такая коррекция не редкость, а норма. Считают, что без нее при редупликации ДНК из E.coli 5—10% нуклеотидов были бы присоединены неправильно. Благодаря коррекции 1 ошибка в этой ДНК приходится на десять миллионов пар оснований. Всем бы корректорам такую тщательность!
Литература:
Ахмеджанов М.Ю., Гуз С.Я., Архангельский В.В. Динамика содержания триглицеридов, общего холестерина и его фракций в сыворотке крови больных, перенесших инфаркт миокарда и при санаторно-курортном лечении. // Новое в лабораторной диагностике хронических болезней внутренних органов. - Ужгород, -1983. -С.52.
Бабенко Н.А., Натарова Ю.А. Роль тиреоидных гормонов в регуляции сфинголипидов в печени // Биохимия.1999. -Т.64. -вып. 8. -С.- 1085-1089.
Бабич Л.Г., Шлыков С.Р., Борисова И.А. Энергозависимый транспорт Са+2 во внутриклеточных структурах гладкой мышцы. //Биохимия -1994. -Т.59. -вып.8. -С. 1218 - 1222.
Баев В.П., Булах Е.П. Определение кетоновых тел в крови и тканях. // Лабораторное дело. -1974. -N 9. -С.545.