| Скачать .docx | Скачать .pdf |
Курсовая работа: Кинетика клеток
Содержание
Введение. 2
1История химической кинетики. 4
2Биокинетика. 6
3Движение клеток. 7
4Системы подвижности эукариотических клеток. 10
4.1Микротрубочки. 10
4.1.1Микротрубочки в ресничках и жгутиках. 11
4.1.2Самосборка микротрубочек и ее регуляция. 14
4.2Микрофиламенты.. 15
4.2.1Свойства актинов немышечных клеток. 17
4.3Микротрубочки, микрофиламенты и клеточные мембраны.. 18
5Генерация движения микрофиламентами. 23
5.1Регуляция скольжения белками микрофиламентов. 25
Список используемой литературы.. 29
Введение
Первый признак, который отличает живое от неживого, это движение, постоянное развитие во времени. Мы заведомо знаем, что события складываются из последовательностей весьма определенных стадий и циклов, разворачиваются во времени. Каждая стадия события имеет продолжительность, определенность и значимость.
Очевидно, что в ритмах живого лежат последовательности превращений молекул. Что определяет протекание биологических процессов во времени? Каковы пути и возможности ускорений биохимических реакций? Какая стадия определяет скорость того или иного биологического явления? Какие события на молекулярном уровне задают динамику развития в целом? Постановка такого рода вопросов связана с развитием области количественных исследований, которая называется биологической кинетикой (биокинетикой).
Исследование количественных закономерностей развития биологических процессов на молекулярном уровне во времени составляет предмет биологической (биохимической) кинетики. В задачи биокинетики входит выяснение механизмов, определяющих скорости и природу процессов.
Изучение динамики биологических процессов охватывает большой круг явлений. Многие из них уже в настоящее время могут быть интерпретированы на молекулярном уровне. За последние десятилетия существенный прогресс в данной области в значительной степени связан с интенсивным изучением ферментов и ферментных систем. Именно ферменты в большинстве случаев являются кинетическими элементами, определяющими скорости и направления развития биопроцессов. Самосогласованность биологических процессов на молекулярном уровне существенным образом определяется отработанными эволюцией процессами обмена информацией с помощью сигнальных молекул и белковых рецепторов. Эти процессы характеризуются вполне определенными кинетическими закономерностями, анализу которых посвящен значительный раздел называемый молекулярной рецепцией.
Наконец, большой и важный раздел современной биокинетики связан с анализом кинетики роста и эволюции клеточных популяций. Клетка как элементарная ячейка жизни представляет собой высокоорганизованный реактор, обладающий свойством полностью воспроизводить себя во всей сложности состава и структуры. Понимание динамики клеточного роста принципиально важно как при решении задач микробиологии, биотехнологии и управляемого биосинтеза, так и для развития количественной медицины, онкологии, для понимания и управления механизмами старения.
Итак, Биокинетика — наука, изучающая на молекулярном уровне закономерности развития биологических процессов в системах in vitro, живых органах и тканях, клеточных популяциях
1. История химической кинетики
Выделение химической кинетики в самостоятельную научную дисциплину произошло в 80-е гг. XIX в. Это оказалось возможным благодаря накоплению и систематизации громадного фактического материала по изучению механизмов химических превращений.
Вероятно, одной из самых первых попыток изучения механизма химической реакции являются работы Р. Бойля, который в 1680 г. наблюдал яркое свечение фосфора при уменьшении давления воздуха. Несколько позже, в начале – середине XVIII в. А. Лавуазье изучал образование оксида ртути при взаимодействии этого металла с кислородом. В рамках этой работы в 1770 г. Лавуазье провел один из самых длинных за всю историю химии опыт, длившийся 100 дней.
Вслед за Лавуазье ученые-химики не раз предпринимали попытки исследовать закономерности химических превращений, найти материальный субстрат химических реакций
В 1789 г. английский ученый У. Хиггинс предположил, что процесс растворения железа в кислоте является сложной реакцией, протекающей с образованием промежуточных соединений. Эти промежуточные соединения распадаются к моменту окончания реакции из-за действия «силы сродства». Благодаря работе Хиггинса в химию было впервые введено понятие времени протекания химической реакции.
В 1894 г. ученица Хиггинса У. Фульгем показала, что вода способна образовывать промежуточные соединения с реагентами при протекании химических реакций в водных растворах. Фульгем предположила, что вода катализирует реакции, происходящие в растворах.
Гипотезы английских ученых У. Хиггинса и У. Фульгем о наличии промежуточных продуктов химической реакции соответствуют современным представлениям, согласно которым любая химическая реакция представляет собой совокупность элементарных актов (превращений). Эти превращения происходят из-за взаимодействия между собой молекул исходных веществ (субстратов реакции) и приводят к образованию продуктов реакции. При этом любое химическое превращение обратимо.
Впервые предположил обратимый характер химических превращений Д.И. Менделеев в книге «Основы химии» (1869). В этой книге он уделяет особое внимание влиянию внешних условий на закономерности протекания химических реакций и подчеркивает сложный характер химических превращений.
Дальнейшее развитие учения о механизмах химических реакций связано с именами A.M. Бутлерова и М.Д. Львова. В 1861 г. Бутлеров сформулировал теорию химического строения, в которой подчеркивал, что закономерности протекания химических реакций зависят от строения веществ, вступающих в реакцию. Теория химического строения Бутлерова была развита и дополнена его учеником М.Д. Львовым (1884).
Один из основных разделов кинетики изучает кинетику биологических реакций; этот раздел принято называть биокинетикой. Биокинетика является пограничной наукой, возникшей на стыке биохимии и химической кинетики (см рис. 1). Выделение биокинетики в отдельную дисциплину неслучайно, оно логически оправдано и связано с исключительной значимостью кинетических процессов для всех живых организмов. Биокинетика — относительно молодая наука. Термин «Биокинетика» был введен И.В. Березиным и С.Д. Варфоломеевым в 1979 г. Традиционно в курсе биокинетики рассматриваются ферментативные реакции, процессы взаимодействия лигандов с рецепторами и процессы клеточного роста.
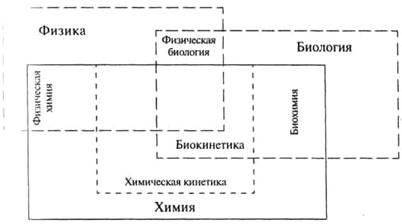
Рисунок 1. « Соотношение между основными естественнонаучными дисциплинами».
При изучении живых систем часто говорится о движении: движутся хромосомы к полюсам клетки во время митоза, перемещаются вакуоли клеточных органелл, движется клеточная поверхность. Кроме того, в клетках растений и животных наблюдаются токи цитоплазмы (например, в растительных клетках или у амебы). Более того, отдельные клетки (свободноживущие одноклеточные организмы или специфические типы клеток в многоклеточных животных организмах) обладают способностью активно перемещаться, «ползать» (см. рис. 3.). Некоторые клетки имеют специализированные структуры, реснички или жгутики, которые позволяют им или самым перемещаться, или перемещать окружающую их жидкость. Наконец, у многоклеточных животных организмов есть специализированные клетки, мышечная работа которых позволяет производить различные движения органов, отдельных его частей и всего организма. В основе всех этих многочисленных двигательных реакций лежат общие молекулярные механизмы. Кроме того, наличие каких-либо двигательных аппаратов должно сочетаться и структурно связываться с существованием опорных, каркасных или скелетных внутриклеточных образований. Поэтому можно говорить (описывать и изучать) об опорно-двигательной системе клеток.
Само понятие о цитоскелете или скелетных компонентах цитоплазмы разных клеток было высказано Н.К.Кольцовым, выдающимся русским цитологом еще в начале XX века. К сожалению, они были забыты и только в конце 1950-х годов с помощью электронного микроскопа эта скелетная система была переоткрыта.
Огромный вклад в изучение цитоскелета внес метод иммунофлуоресценции, который помог разобраться в химии и динамике этого чрезвычайно важного компонента клетки. Цитоскелетные компоненты представлены нитевидными, неветвящимися белковыми комплексами, или филаментами (тонкими нитями).
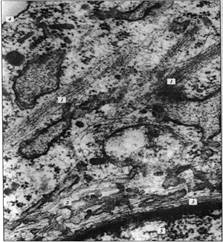
Рисунок 2. «Микрофотография элементов цитоскелета, полученная с помощью электронного микроскопа». 1 - пучки микрофиламентов; 2 - микротрубочки; 3 - промежуточные филаменты 4 плазматическая мембрана; 5 – ядро.
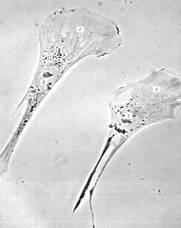
Рисунок 3. «Поляризованные движущиеся фибробласты в культуре ткани». 1 – ламеллоплазма; 2 — ядро
Существуют три системы филаментов, различающихся по химическому составу, ультраструктуре и функциональным свойствам. Самые тонкие нити — это микрофиламенты; их диаметр составляет около 6 мм, и состоят они в основном из белка актина. К другой группе нитчатых структур относятся микротрубочки, которые имеют диаметр 25 нм и состоят в основном из белка тубулина. Третья группа представлена промежуточными филаментами с диметром около 10 нм (промежуточным по сравнению с 6 и 25 нм), образующимися из разных, но родственных белков (рис. 2 и 4).
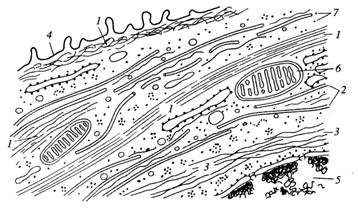
Рисунок 4. «Схематическое изображение цитоскелетных компонентов клеток». 1 – микрофиламенты; 2 – микротрубочки; 3 – промежуточные филаменты; 4 – плазмалитическая мембрана; 5 – ядро; 6 – митохондрии; 7 – рибосомы.
4. Системы подвижности эукариотических клеток
Микротрубочки предстали как особые внутриклеточные структуры (особенно внутри жгутика) благодаря применению методов электронной микроскопии. В 1946 г. Джакус и Холл продемонстрировали наличие одинаковых по диаметру трубчатых структур в ресничках Paramecium. Впоследствии трубчатые структуры обнаружили почти во всех клетках. Стали считать, что они характерны для эукариотичсского уровня организации. Однако микротубулярные структуры были обнаружены и у прокариот — в цитоплазме спирохет. Морфологически и биохимически эти микротрубочки подобны тем, которые наблюдаются в эукариотических клетках. Это открытие поддержало «экзогенную гипотезу» происхождения микротрубочек эукариот, согласно которой реснички и жгутики клеток высших организмов считаются приобретенными извне—путем симбиоза клеток, ранее не имевших жгутиков или ресничек, со спирохетами, содержавшими микротрубочки.
Микротрубочки представляют собой длинные полые цилиндры, наружный диаметр которых около 24 нм, а внутренний—15 нм. В большинстве клеток длина трубочек обычно не превышает нескольких микрон, хотя в некоторых специализированных клетках, например в моторных нейронах центральной нервной системы, трубочки могут быть длиной в несколько сантиметров. Стенка микротрубочки (около 5 нм толщиной) построена из продольно ориентированных протофибрилл. Протофибриллы состоят из глобулярных субъединиц (их очень хорошо видно в электронный микроскоп при негативном окрашивании), содержащих только один белок – тубулин (рис. 5). Помимо того, что микротрубочки имеют прямое отношение к подвижности клетки, они участвуют и в других процессах, более или менее связанных с подвижностью, например в поддержании формы клетки, во внутриклеточном транспорте веществ, в секреции клеточных продуктов, в движении хромосом при делении клетки и, возможно, в осуществлении сенсорных связей, а также в перемещении компонентов клеточной мембраны. Микротрубочки могут быть рассеяны по всей цитоплазме, а могут быть собраны в организованные структуры.

Рисунок 5. «Электронные, микрофотографии микротрубочек в краевых областях эритроцитов тритона»: а — продольный срез, на котором: видна линейная упаковка субъединиц в микротрубочках; б —поперечный срез; видно, что стенка каждой микротрубочки образована тринадцатью субъединицами. Негативное окрашивание. ×200 000.
Чтобы понять, как клетка движется с помощью микротрубочек, мы рассмотрим их организацию в ресничках жгутиках эукариотических клеток, в сократимом аксостиле некоторых жгутиконосцев, а также в микротубулярной сети цитоплазмы животных клеток.
4.1.1 Микротрубочки в ресничках и жгутиках
Эукариотические реснички и жгутики представляют собой специализированные структуры, выступающие за пределы клеточной поверхности и способные двигаться. Эти два типа органелл идентичны по своей структуре; в функциональном отношении между ними обнаруживаются лишь незначительные, различия; Жгутики и реснички удается легко и в довольно больших количествах отделить от клеток, иногда просто энергичным встряхиванием клеточной суспензии. Это облегчило исследование их структуры и химического состава.
Структурная организация ресничек и жгутиков практически во всех исследовавшихся клетках отличается удивительным единообразием: центральную пару микротрубочек окружает кольцо из микротрубочек (обычно парных). Микротрубочки связаны между собой поперечными мостиками; вся конструкция называется аксонемой жгутика. Расположение микротрубочек и мостиков в аксонеме изучено в настоящее время довольно подробно; оно схематически представлено на рис. 6. В центре находится пара микротрубочек, окруженных общей оболочкой из белковых колец, плоскость которых наклонена к оси аксонемы. Стенка каждой из центральных микротрубочек образована тринадцатью продольными рядами (протофибриллами) белковых субъединиц. Эту центральную структуру, окружают девять расположенных по кольцу пар микротрубочек. В каждом дублете одна микротрубочка (основная или А) имеет полностью замкнутую стенку, состоящую из 13 протофибрилл (рис. 7); другая микротрубочка (микротрубочка В) имеет серповидное сечение и примыкает к микротрубочке А, так что ее стенка состоит из меньшего числа протофибрилл, обычно из 10. Каждая микротрубочка А соединена с микротрубочкой В соседнего дублета с помощью мостика из белка, называемого нексином. От обращенной внутрь поверхности микротрубочки А отходит по направлению к центру белковый выступ — так называемая радиальная спица. Вдобавок каждая микротрубочка А имеет двойной ряд боковых ручек (их длина около 14 нм), построенных из белка (динеина) и направленных к соседнему дублету. Все эти выступы расположены с определенной периодичностью относительно оси микротрубочки А и, как будет видно из дальнейшего, играют очень важную роль в генерации движения. Описанное выше строение аксонемы соответствует всей средней части жгутика, т. е. за исключением его концевой части и основания, где аксонема устроена несколько иначе. На самом конце центральная пара микротрубочек обычно длиннее, чем окружающие ее периферические дублеты, так что жгутик на конце заострен. Нередко микротрубочки В бывают короче микротрубочек А, и периферические дублеты вблизи конца жгутика переходят в одиночные трубочки (рис. 6). На одиночных микротрубочках А в этой области жгутика выступов нет.
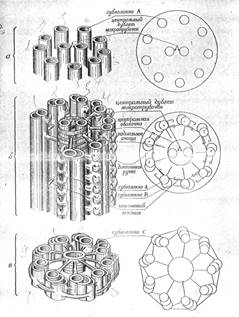
Рисунок 6. «Схематическое изображение аксонемы жгутика». Показана взаимосвязь между элементами структуры на разных уровнях: а — на конце жгутика, б — в средней части и в — на уровне базального тела.
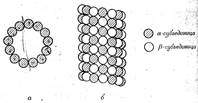
Рисунок 7. «Схематическое изображение расположения тубулиновых субъединиц в микротрубочке»: а — поперечный срез; б — вид сбоку.
Специализированная структура — базальное тело , — из которой исходит жгутик, находится в цитоплазме клетки у основания жгутика. Она может быть различна в клетках разных типов, однако при этом основные, общие черты сохраняются. В базальном теле ресничек Tetrahymena piriformis наряду с микротрубочками А и В имеются еще микротрубочки С (они образованы десятью протофибриллами), так что по периферии реснички располагаются не дублеты, а триплеты микротрубочек, Эти триплеты связаны между собой боковыми выступами (поперечными мостиками) и прикреплены к центральному кольцу радиальными спицами (рис. 6). Базальное тело реснички или жгутика по своей структуре, похоже на центриоль животных клеток. И базальное тело, и центриоль действуют как организаторы ассоциации микротрубочек; в первом случае/образуется ресничка или жгутик, во втором — мнтотическое веретено.
Еще одна интересная часть жгутика — это так называемый воротничок, кольцевой валик, окружающий основание жгутика там, где он выходит из клетки. В электронный микроскоп па поперечных срезах воротничка в этой области видны волокнистые соединения между периферическими дублетами жгутика и внутренними слоями клеточной мембраны. Они жестко закрепляют микротрубочки в клеточной мембране и участвуют в движении жгутика.
Подвижность ресничек и жгутиков обусловлена свойствами именно аксонемы, но не других компонентов этих органелл (т. е. клеточной мембраны, цитоплазмы и др.).
4.1.2 Самосборка микротрубочек и ее регуляция
В любой живой клетке происходит агрегация тубулина с образованием микротрубочек и распад этих структур. Вопрос о том, как регулируются процессы сборки и деструкции микротрубочек имеет первостепенное значение для понимания многих проявлений жизнедеятельности клетки.
Изучение invitro полимеризации тубулина, выделенного из нервных клеток, оказалось ценным источником информации для понимания механизма сборки микротрубочек invivo и путей управления этим процессом. Если нейротубулин находится в растворе в виде димеров в присутствии связанных с ним белков, то в определенных условиях легко идет процесс самосборки. Полимеризация происходит в условиях умеренной ионной силы, при слабо кислом рН (около 6,6—0,7), в присутствии нуклеотидов (GTP) и ионов, магния. Направление реакции зависит также, от температуры: invitro при физиологических температурах идет полимеризация тубулина с образованием микротрубочек, а снижение температуры до 0°С вызывает их быструю деполимеризацию (рис. 8).
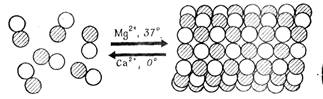
Рисунок 8. «Реакция полимеризации димеров тубулина с образованием микротрубочек. Указано влияние двухвалентных ионов и температуры».
4.2 Микрофиламенты
Микрофиламенты эукариотических клеток представляют собой длинные нитевидные структуры толщиной 5-7 нм, находящиеся в цитоплазме. Они состоят главным образом из актина (хотя можно обнаружить и другие белки), в связи, с чем их часто называют актиновыми филаментами (или нитями). Они иногда образуют группы или пучки, а в некоторых высоко специализированных клетках, например в мышечных волокнах, упорядоченные и стабильные структуры. Чаще, однако из микрофиламентов формируются нестабильные пучки или тонкие сетчатые структуры, их форма и локализация в клетке изменяются в зависимости от фазы жизненного цикла клетки, ее движения и т. п. Это означает, что, подобно цитоплазматическим микротрубочкам, микрофиламенты немышечных клеток представляют собой изменяемые структуры, способные к самосборке или деполимеризации на составляющие их молекулы в зависимости от нужд клетки.
Морфологию микрофиламентов и их поведение в процессе клеточного движения и цикла развития клетки много изучали на культуре животных клеток с помощью флуоресцирующих антител против актина, а также против других белков (миозин, тропомиозин и а-актинин), которые часто обнаруживаются в микрофиламентах. Когда клетки находятся в состоянии покоя и прикреплены к стенкам сосуда, в котором живут, в них можно наблюдать длинные пучки микрофиламентов, тянущиеся сквозь всю клетку, по-видимому, непосредственно под клеточной мембраной. В «возбужденные» выступы клетки с нестабильными, как бы волнующимися краями они обычно не заходят. Пучки микрофиламентов называются стрессовыми, типичны для нормальных клеток и отсутствуют в клетках, трансформированных онкогенными вирусами. Когда клетки движутся, в дополнение к стрессовым волокнам появляются топкие пучки актиновых нитей, рассеянные в цитоплазме; они особенно заметны в направленных вперед выступах клеток.
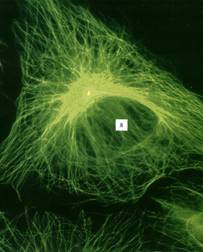
Рисунок 9. «Сеть микротрубочек, окрашенная мечеными антителами к тубулину в клетке культуры ткани в Gj -периоде». В клеточном центре желтым окрашена центриоль, связанная с антителами к γ -тубулину, Я – ядро.
4.2.1 Свойства актинов немышечных клеток
Актин широко распространен во всех эукариотических клетках. Его можно обнаружить как в растворимой мономерной форме, так и в полимерной — в виде филаментов. В некоторых активно передвигающихся клетках (амебы, макрофаги, тромбоциты) актин преобладает среди белков клеточного экстракта: его содержание может достигать 20—30% от общего белка, В менее подвижных клетках содержание актина заметно меньше—1—2% от общего белка. Однако если вспомнить, что в эукариотической клетке одновременно присутствуют многие, тысячи различных белков, то такое небольшое количество весьма существенно.
Благодаря относительно высокому его содержанию актин удается выделить из клеточных систем. Более акриламидном геле в присутствии додецилсульфата натрия. Выявились три основных полипептида, один из которых соответствует актину. Два других белка (мол. массы 230 000 и 250000), по-видимому, ответственны за I поддержание актина в неполимеризованном состоянии. Обработка протеолитическим ферментом (трипсином) позволяет отделить мономерный актин от высокомолекулярных белков. Акросомальную реакцию у сперматозоидов Thyone можно вызвать различными актива- I торами; повышение, внутреннего рН освобождает актин от связанных с ним белков, делая тем самым возможной полимеризацию. Процесс полимеризации инициирует 1 специфическая структура, которую можно увидеть на дне акросомальной чаши и которая называется актомером. Она представляет собой пучок из примерно 25 коротких микрофиламентов, заключенный в плотное аморфное вещество. В момент активации актомер начинает действовать как ядро полимеризации, из которого вырастают микрофиламенты.
Па основании этих и других данных высказано предположение, что полимеризацией и ростом микрофиламентов управляют какие-то организующие центры, подобно тому, как центр, управляющий жгутиковым синтезом, регулирует сборку микротрубочек.
Специфические точки роста актиновых филаментов обнаружены в семенных клетках Mitylus и Limulus, a также при формировании кишечных ворсинок; вполне возможно, что они есть и во многих других объектах. Действительно, в немышечных клетках часто видно, что микрофиламенты прикреплены к определенным точкам клеточной мембраны, однако пока неясно, управляют ли эти точки сборкой микрофиламентов или же прикрепление к ним происходит уже после.

Рисунок 10. «Пучки актиновых микрофиламентов в клетках культуры ткани, окрашенных флуоресцирующими антителами Ядра – фиолетовые».
4.3 Микротрубочки, микрофиламенты и клеточные мембраны
Есть основания считать цитоплазматические микротрубочки и микрофиламенты тесно взаимосвязанными структурами одной и той же системы. Они играют сходную роль в генерации движения, встречаются в одних и тех же клетках в роли опорных структур, ведут себя сходным образом в зависимости от фазы клеточного цикла и образуют морфологически похожие структуры. К сожалению, приходится отметить, что, несмотря на приведенные выше соображения, у нас нет прямых доказательств взаимосвязи между микротрубочками и микрофиламентами. Многие данные, полученные различными методами, косвенно, но с большой вероятностью указывают на то, что между этими двумя системами есть связь, так что почти все признают, что такая связь на самом деле существует.
Например, с морфологической точки зрения исследовали распределение в клетке микротрубочек и микрофиламентов, особенно в процессе митоза. Фудживара и Поллард с помощью антител к тубулину, меченных флуоресцеином, и антител к миозину, меченных родамином, сравнили распределение микротрубочек и микрофиламентов в одной и той же клетке. В полностью распластанных клетках они не обнаружили морфологических связей между этими двумя типами волокон. Однако в митотическом веретене обусловленная миозином флуоресценция так сочеталась с флуоресценцией микротрубочек, что это указывало на тесную взаимосвязь между микрофиламентами и микротрубочками.
Более косвенные данные получены в опытах с ингибиторами микротубулярных структур (колхицин или его производное колцемид) и ингибиторами микрофиламентов (цитохалазин В). В качестве примера такого подхода мы рассмотрим влияние ингибиторов микротрубочек и микрофиламентов на явление образования «шапки» (capping), наблюдаемое в клетках млекопитающих. Реакция образования «шапки» возникает после обработки клетки бивалентными антителами или другими лигандами, способными взаимодействовать с белками на клеточной мембране. Она заключается в том, что связываемые молекулы лиганда скапливаются на ограниченном участке клеточной поверхности, образуя как бы шапку. Образование такой шапки угнетается цитохалазином-В, откуда следует, что микрофиламенты играют активную роль в перемещении молекул в мембране клетки. Во многих системах угнетение образования шапки удается снять колхицином или другими веществами, вызывающими деполимеризацию микротрубочек. Если разрушить и микротрубочки, и микрофиламенты комбинированным действием колхицина и цитохалазина-В, образовавшаяся шапка, которая обычно довольно стабильна, распадается, так что молекулы, из которых она состояла, распределяются по всей поверхности клетки. Эти результаты указывают на то, что и микротрубочки, и микрофиламенты участвуют в регуляции перемещения молекул в клеточной мембране: микрофиламенты проявляют сократительную активность, а микротрубочки служат для них опорой. Можно также предполагать, что для латерального перемещения белков в липидном бислое мембраны необходимо прямое взаимодействие между микрофиламентами и мембранными белками.
Есть, однако, и более прямые данные о том, что микрофиламенты соединены с поверхностными белками, т. е. с клеточными мембранами. В разных лабораториях различными методами показано, что непосредственно под шапкой, образующейся на лимфоцитах (после обработки антителами) или на клетках гранулезы яичников (после обработки конканавалином А), собирается много микрофиламентов. Там же накапливаются миозин и тубулин. В более поздней работе применили метод осаждения актина из лизатов клеток мышечным миозином, после чего идентифицировали вещество, преципитирующее вместе с актином; показано, что актин осаждается вместе с молекулами, обычно находящимися на внешней поверхности клеточных мембран, такими, как антигены Н-2 и иммуноглобулины. Количество иммуноглобулинов, связанных с актином, больше в клетках с «шапками», чем в клетках с неизмененным распределением антигенов по поверхности. Исходя из полученных результатов, можно предположить, что агрегация поверхностных белков в «шапку» вызывается возникновением новых связей с актином. Это может служить дополнительным свидетельством в пользу того представления, что микрофиламенты контролируют, по крайней мере, некоторые типы перемещения компонентов мембраны.
Приведенные выше данные демонстрируют также, что актиновые филаменты прямо связаны с мембранами. Результаты, полученные с помощью других методов и на других объектах, также указывают на это. Например, микрофиламенты часто выделяются при очистке цитоплазматических мембран. Впервые это показано для Acanthamoeba caslellani , а впоследствии подтвердилось практически для всех типов клеток. В очищенных препаратах мембран микрофиламенты можно увидеть в электронный микроскоп (причем они прикреплены к внутренней поверхности мембраны); кроме того, при электрофорезе препаратов мембран в полиакриламидном геле в присутствии додецилсульфата натрия обнаруживается актин.
Далее, есть прямые микроскопические наблюдения примембранных областей, в которых много микрофиламентов. Иногда, например, в случае ворсинок, очень четко видно, что микрофиламенты прикреплены к мембране на конце ворсинки. На электронных микрофотографиях поперечных сколов ворсинок центральный пучок микрофиламентов соединен с мембраной ворсинки как бы спицей.
Как микрофиламенты прикрепляются к мембране, пока неясно. Непосредственно ли присоединены микрофиламенты к мембранам или же их связь осуществляется через какие-то другие белки? Второе предположение кажется более правдоподобным; наиболее вероятные кандидаты на роль такого связующего белка — а-актинин или подобные ему белки. Показано, что а-актинин присутствует в кончике ворсинки — там, где микрофиламенты прикрепляются к ее мембране; кроме того, он обнаружен в цитоплазматической мембране и в мембранах секреторных везикул вблизи микрофиламентов. Таким образом, предполагается, что α-актинин играет одну и ту же роль и в мышечных волокнах (где он связывает актиновые нити с Ζ-мембранами) и в немышечных клетках. Возможно, микрофиламенты взаимодействуют с мембранами также через посредство миозина, и есть данные, указывающие на то, что миозин может выступать как трансмембранный белок.
Менее убедительны эксперименты, поставленные с целью выявить прямое взаимодействие, между микротрубочками и мембранами. По данным многих исследователей, колхицин связывается с мембранами, однако из-за того, что такому связыванию могут способствовать самые разные факторы, это нельзя расценивать как доказательство присутствия тубулина в клеточной мембране. Более прямые данные, указывающие на то, что тубулин входит в состав мембран, получены на синаптосомах головного мозга и на мембранах ресничек некоторых моллюсков. Во всяком случае, связь с мембранами для тубулина, по-видимому, не столь обычное явление, как для актина.
Кратко рассмотрев данные о взаимодействиях между микротрубочками, микрофиламентами и мембранами, и помня о распределении элементов цитоскелета в животной клетке, мы можем обсудить роль цитоскелета и клеточной мембраны (рис. 11).
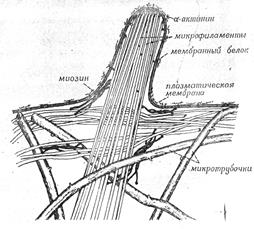
Рисунок 11. «Предполагаемые взаимоотношения между микротрубочками, микрофиламентами и клеточной мембраной».
Согласно предполагаемой модели, в интерфазных клетках микротрубочки образуют внутренний каркас, расходясь от центра к периферии клетки. Этот каркас не участвует непосредственно в генерации движения, а служит опорой для расположенных ближе, к поверхности клетки структур.
5. Генерация движения микрофиламентами
Микрофиламенты могут генерировать движение двумя различными способами: путем скольжения — согласно этому механизму актиновые и миозиновые нити скользят друг относительно друга – или же просто путемсборки и дезагрегации пучков микрофиламентов. Опять мы сталкиваемся с удивительным соответствием между системой микрофиламентов и системой микротрубочек: обе системы генерируют движенце, одними и тещи же способами. Это представляет интерес с эволюционной точки зрения, поскольку указывает на то, что системы, которые возникли, по-видимому, независимо друг от друга, решили проблему генерации движения одинаково, хотя и с помощью различных материалов.
Модель скольжения микрофиламентов, описывающая зависимые от актомиозина движения немышечных клеток, исходит из данных о сокращении мышц, модель которого можно применить с незначительными модификациями к микрофиламентам.
В немышечных клетках такая система подвижности состоит из актиновых микрофиламентов, один конец которых прикреплен к каким-либо структурам клетки (к клеточной мембране, микротрубочкам или другим органеллам, а другой конец свободен;, менаду свободными концами двух противолежащих актиновых микрофиламентов находятся биполярные миозиновые нити. Когда два противолежащих актиновых микрофиламента скользят вдоль миозиновой нити, их свободные концы сближаются, а закрепленные концы тянут за собой те структуры, к которым они присоединены. В мышцах всё это происходит в структурах, специально предназначенных для генерации движения (саркомерах). Саркомер состоит из двух пучков актиновых нитей (они прикреплены к Z -мембранам, ограничивающим саркомер) с миозиновыми нитями между ними. В результате скольжения нитей саркомер укорачивается, что соответствует сокращению мышцы. В немышечных клетках такое скольжение приводит к сближению структур, к которым прикреплены противоположные концы микрофиламентов
Рассмотрим теперь, как же при взаимодействии актиновых и миозиновых нитей возникает скольжение. Инициатором является миозин, точнее головки его молекулы, где находятся центры АТРазной активности. Миозиновые головки отличаются большим сродством к АТР, и при его избытке каждая головка связывает одну молекулу АТР.
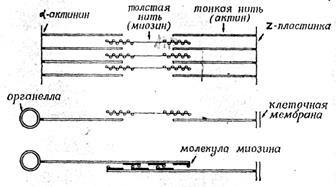
Рисунок 12. «Модели взаимодействия актина и миозина. Вверху, мышце; посередине и внизу: в немышечных клетках».
Связав АТР, миозиновая головка сразу же переходит в активированное состояние с высоким сродством к актину и прикрепляется к одной из актиновых субъединиц ближайшего микрофиламента. Связывание с актином немедленно вызывает гидролиз АТР, за счет выделившейся - при этом энергии головка поворачивается на небольшой угол, что немного перемещает актиновый филамент, к которому головка прикреплена. При утилизации новых порций АТР такой цикл повторяется многократно, скольжение становится заметным. Мы привели весьма упрощенное описание процессов, связанных с актомиозинзависимой сократимостью мышечных волокон. Более полное описание читатель найдет в другой книге этой же серии (R. Μ. SimmonsMuscleContraction"). Этот основной механизм взаимодействия актина и миозина можно, по-видимому, распространить также и • на немышечные клеточные системы.
5.1 Регуляция скольжения белками микрофиламентов
В мышечных клетках актиновые нити содержат (кроме, актина) два регуляторных белка — тропомиозин и тропонин, благодаря которым скольжение чувствительно к концентрации ионов Са2+ . Связывание комплекса миозин — АТР с актином возможно только в присутствии Са2+ , т. е. Са2+ служит, регулятором мышечного сокращения. Концентрация Са2+ внутри саркомеров регулируется высвобождением его из саркоплазматического ретикулума при деполяризации мембраны.
Тот факт, что миозин действительно присутствует в микрофиламентах животных клеток, был продемонстрирован с помощью антител к миозину, меченных флуоресцеином. Возможно также, что в немышечных клетках миозиновые молекулы существуют не в виде типичных нитей с выступающими головками, а просто как двуглавые мономеры, сохраняющие способность связывать актиновые микрофиламенты. Другая модель, которую предложили Марута и Корн, предполагает, что одиночные, миозиновые молекулы присоединены к актиновому филаменту стержневыми участками тяжелых цепей, так что свободные головки могут взаимодействовать с соседними актиновыми филаментами. В этом случае подвижность обеспечивалась бы непосредственно скольжением двух актиновых микрофиламентов друг относительно друга, т, е. так, как скользят микротрубочки в ресничках и жгутиках. Согласно этой модели, одноглавый миозин, который, по-видимому, не способен образовывать биполярные нити (как, например, миозин Acanthamoeba) , тоже мог бы участвовать в генерации движения.
Поскольку подвижность зависит от взаимодействия актина и миозина, факторы, регулирующие это взаимодействие, можно рассматривать как регуляторы клеточной подвижности. В мышечных клетках управление сокращением осуществляется с помощью ионов Са2+ и системы тропомиозин — тропонин, связанной с актиновыми нитями. В немышечных клетках регуляция еще недостаточно изучена. Ясно, однако, что в клетках различных типов может, быть много разных регуляторных систем — одни из них основаны на действии Са2+ , а Другие реализуют другие механизмы.
Ферментативная активность при взаимодействии очищенных препаратов актина и миозина из немышечных клеток не зависит, как правило, от концентрации Са2+ , однако известны примеры Са2+ чувствительной АТРазной активности актомиозина из тканей мозга, из лейкоцитов, тромбоцитов и плазмодия миксомицета Physarum polycephalum . Чувствительность к Са2+ можно определить, регистрируя сокращение актомиозиновых нитей в клеточных экстрактах (это сделано на амебах и некоторых других клетках). Данные invivo о подвижности, чувствительной к Са2+ , в которой участвуют микрофиламенты, получены при исследовании токов цитоплазмы у Amoeba proteus, Chaos carolinensis и Physarum, а также АТР-зависимого сокращения изолированных полосок щеточной каемки кишечного эпителия.
В мышечных клетках сокращение регулируется Са2+ -связывающим белком системы тропомиозин — тропонин, поэтому некоторые исследователи искали подобные белки и в немышечных клетках. Белки, подобные тропомиозину, удалось найти в тромбоцитах, в тканях мозга, в поджелудочной железе и в культуре фибробластов мышц; Са2+ -связывающий белок, похожий на тропонин мышц, недавно выделили из мозга куриных эмбрионов. Белки, придающие, актомиозиновому комплексу чувствительность к Са2+ , выделены из Physarum и Dictyosteиит, однако эти данные нуждаются в дальнейшей проверке.
Наряду с кальциевой регуляцией, несомненно, существуют и другие регуляторные системы, контролирующие взаимодействие актина и миозина.
Одной из них может быть регуляция фосфорилирования миозина. Было показано, например, что в тромбоцитах АТРазная активность миозина, стимулированная актином, возрастает приблизительно в 5 раз, когда легкая (17 000) цепь миозина фосфорилируется особой протеинкиназой в присутствии АТР. Это позволяет предполагать, что фосфорилирование прямо влияет на взаимодействие актина и миозина. Однако относительно этой системы пока еще преждевременно делать окончательные выводы.
Из приведенных выше примеров должно быть ясно, что в настоящий момент еще нет единой теории регуляции актомиозинового взаимодействия в немышечных клетках. Отчасти это обусловлено сравнительной скудостью сведений, которыми мы располагаем по этому вопросу, а отчасти—сложностью самой проблемы. Вероятно, что в регуляции взаимодействия актина с миозином в немышечных клетках участвует многих систем. Для некоторых клеток важное значение имеют ионы Са2+ , о других механизмах регуляции известно пока еще слишком мало.
Есть еще один способ генерировать движение, который используется не для перемещения всей клетки как целого, а для движения отдельных ее частей (например, мембран); речь идет о подвижности, обусловленной полимеризацией и деполимеризацией пучков актиновых филаментов. В этом случае движение обусловлено не скольжением, а ростом пучков микрофиламентов, которые при этом отталкивают ту часть клетки, которая контактирует с зоной их роста (обратный процесс, как можно представить себе, происходит при деструкции микрофиламентов).
Пример движения такого типа — уже упоминавшаяся акросомальная реакция. В процессе этой реакции менее чем за 10 с, формируется прямой пучок микрофиламентов, выпячивающий мембрану сперматозоида в направлении яйца.
В цитоплазме некоторых немышечных клеток нередко обнаруживают другой тип надмолекулярной организации актиновых мономеров: вместо обычных пучков микрофиламенты образуют тонкую трехмерную сеть.
Это явление можно воспроизвести invitro; оно известно под названием процесса желатинизации. Такие, как их еще, называют, переходы золь — гель имеют, по-видимому, существенное значение для регуляции вязкости цитоплазмы и изменения формы клетки, и, хотя эти функции могут быть косвенно связаны с движением клетки, их нельзя считать истинной подвижностью.
Список используемой литературы
1.Биологический энциклопедический словарь. / Гл. ред. М. С. Гилярон; Редкол.: А. А. Баев, Г. Г. Винберг, Г. А. Заварзип и др. — 2-е изд., исправл. — М.: Сов. энциклопедия, 1989. — 864 с, ил.
2.Варфоломеев С. Д., Гуревич К. Г. Биокинетика: Практический курс. – М.: ФАИР-ПРЕСС, 1999.– 720 с: ил.
3.Мецлер Д. Биохимия: Химические реакции в живой клетке. В 3-х томах том 1, 2, 3. Пер. с англ. – М.: Изд-во «Мир», – 1980.
4.Ченцов Ю.С. Введение в клеточную биологию: Учебник для вузов. – 4-е изд., перераб, и доп. / Ю.С. Ченцов. – М.: ИКЦ «Академкнига», 2004. – 495 с: ил.
5.Юрина Н. А., Радостина А. И. Гистология: Учебник. – М.: Медицина, 1995. – 256 с; ил.
6.Гистология: Учебник. 2-е изд., перераб, и доп. / Под ред. Э.Г. Улумбекова, Ю./ Челышева. – М.: ГЭОТАР-МЕД, 2002. – 672 с: ил.