| Похожие рефераты | Скачать .docx | Скачать .pdf |
Курсовая работа: Основные типы взаимодействий генов
Содержание
Введение
1. Проявление действия гена
2. Форми взаимодействия аллельных генов: полное и неполное доминирования, кодоминирование
3. Типы взаимодействия неаллельных генов
3.1 Комплементарность
3.2 Эпистаз
3.3 Полимерия
3.4 Гены-модификаторы
4. Плейотропное действие генов
5. Пенетрантность, экспрессивность, норма реакции
6. Влияние факторов внешней среды на действие генов
Выводы
Список использованной литературы
Введение
В большинстве случаев один признак развивается под влиянием не одного, а двух и более параллелей. Последние вступают во взаимодействие в процессе развития признака, что приводит к его изменению, а отсюда к изменению соотношения фенотипов во втором поколении. Впервые возможность подобного явления была отмечена Г. Менделем, наблюдавшим при скрещивании белоцветковых сортов фасоли с пурпурноцветковыми появление во втором поколении из растения только одного с белыми цветками. Мендель объяснил это тем, что окраска цветков и семян у фасоли «слагается из двух или нескольких совершенно самостоятельных красок, которые в отдельности подчиняются тем же правилам, которым подчиняется каждый другой константный признак у растений». Предположение Менделя было подтверждено дальнейшими исследованиями.
На основании изучения взаимодействия разных пар генов, влияющих на один и тот же признак, установлено несколько основных in нов их взаимодействия: новообразование, комплементарные, или дополнительные, факторы, эпистаз и гипостаз и полимерия. Следует отметить, что при всех типах взаимодействия наследование признаком происходит в строгом соответствии с установленными Менделем правилами наследования; меняется лишь характер расщепления по фенотипу, так как в зависимости от типа взаимодействия отдельные генотипы обнаруживают сходство между собой, в результате чего число различных фенотипов уменьшается против обычного по правилам Менделя. Рассмотрим более подробно тему аллельных и неаллельных взаимодействий генов.
Цель работы : рассмотреть основные типы взаимодействий генов.
Предмет исследования – основные типы и формы взаимодействия генов.
Для достижения поставленной цели необходимо решить основные задачи , которые заключаются в следующем:
1) особенности проявления действия гена;
2) охарактеризовать основные формы взаимодействия аллельных генов;
3) рассмотреть типы взаимодействия неаллельных генов;
4) рассмотреть особенности плейотропного взаимодействия генов;
5) установить основные закономерности модифицирующего действия генов;
6) выяснить влияние факторов внешней среды на действие генов.
В ходе работы над данной темой мы использовали научные методы исследования:
- анализ научной литературы;
- сравнение;
- анализ и синтез.
1. Проявление действия гена
Ген как единица наследственности, детерминирующая признаки, или свойства, организма, имеет определенные характеристики:
1) в своем действии ген дискретен; он определяет присутствие или отсутствие отдельной биохимической реакции, степень развития или подавления определенного признака организма;
2) ген действует градуально: накопление дозы его в соматических клетках может приводить к усилению или ослаблению проявления признака;
3) каждый ген специфичен в своем действии, т.е. отвечает за синтез первичной структуры белковой молекулы;
4) ген может опосредованно воздействовать на течение разных реакций и развитие многих признаков организма, т.е. действовать множественно; это явление носит название множественного, или плейотропного эффекта гена;
5) разные гены, находящиеся в различных парах хромосом, могут действовать однозначным образом на развитие одного и того же органа или свойства организма, усиливая или ослабляя его; это так называемые множественные, или полимерные, гены (полигены):
6) ген вступает во взаимодействие с другими генами, и в силу этого его эффект может изменяться;
7) проявление действия гена зависит от факторов внешней среды;
8) гены взаимодействуют теми продуктами, которые они детерминируют и контролируют в процессе их синтеза.
Как мы увидим дальше, гены действуют на двух уровнях: на уровне самой генетической системы, определяя состояние генов и их работу, скорость репликации ДНК, стабильность и изменчивость генов, и на уровне работы клеток в системе целого организма.
Прежде чем перейти к рассмотрению указанных особенностей действия генов, необходимо условиться о системе их обозначения. До сих пор для обозначения гена мы пользовались любыми, большей частью начальными, буквами латинского алфавита. Поскольку в дальнейшем нам придется касаться характеристики действия генов на определенные признаки, свойства, реакции различных организмов, в ряде случаев ген удобнее обозначать в зависимости от характера его действия.
Для сокращенного написания формул генотипа используются первые буквы названия признака, определяемого данным геном. Чаще всего эти названия даются на английском или латинском языках. Например, рецессивный ген, определяющий развитие признака белого цвета глаз у дрозофилы, называют геном white (или сокращенно w). Доминантная аллель этого гена обозначается как +, но чаще для этого объекта доминантные аллели, свойственные мухам, встречающимся в природе, обозначают той же буквой, но со знаком плюс - w+ . Признаки у организмов, встречающихся в природе, обычно являются доминантными и называются признаками дикого типа, а определяющие их аллели - аллелями дикого типа. Для разных видов организмов принята различная номенклатура генов, хотя это и не очень удобно. Так, ген, определяющий развитие желтой Окраски эндосперма у кукурузы, назван yeliow- у, его доминантная аллель - Y.
Рассмотрим сначала характер действия одной аллельной пары. Одна аллельная пара может определять присутствие или отсутствие признака, изменять количественное или качественное его проявление. Накопление в клетке нескольких доз аллели одного и того же гена приводит к усилению либо к ослаблению признака. Например, количество витамина А и его активность в эндосперме зерна кукурузы зависят от многих генов, и в том числе от доминантных аллелей гена Y.
Клетки эндосперма содержат три набора хромосом. Следовательно, путем скрещивания можно получить четыре различных по генотипу эндосперма кукурузы, содержащих разное количество доминантных аллелей У. Количество витамина А (в единицах активности) при разных дозах одного и того же гена Y оказывается следующим:
При генотипе эндосперма YYY0,05
» > > YYY2,25
» > > УУу5,00
> > > УУУ7,50
Как видно из приведенных данных, действие одной дозы доминантного гена у соответствует примерно 2,25 единиц активности витамина А. С увеличением дозы гена его действие суммируется, или кумулируется. Такой тип действия гена называют кумулятивным, или аддитивным, т.е. суммирующимся. Подобные гены могут определять различные признаки. Так, у кукурузы от них зависят степень окраски зерна, вес и размеры растения, его плодовитость и уровень отдельных физиологических и биохимических процессов.
Гены классифицируют по их проявлению: гены морфологических и биохимических признаков, гены плодовитости (фертильности), гены жизнеспособности и т.д. Эта классификация является чисто условной, так как гены с однозначным действием встречаются редко; например, ген, изменяющий морфологический признак зерна кукурузы - его форму, размер и толщину алейронового слоя, одновременно может быть связан и с различиями в химическом составе. Изучение действия генов в онтогенезе производят различными методами: биохимическими, анатомическими, физиологическими, эмбриологическими и др. Однако проблема изучения функционирования гена осложняется тем, что гены в своем действии не изолированы, а взаимосвязаны. В этом легко убедиться, рассмотрев некоторые типы или системы взаимодействия генов при сочетании нескольких генов, находящихся в негомологичных хромосомах.
2. Формы взаимодействия аллельных генов: полное и неполное доминирования, кодоминирование
Более пристальное рассмотрение элементарных признаков, т.е. признаков, альтернативные состояния которых наследуются по моногибридной схеме, позволяет подойти к проблеме доминирования, исследовать его механизм. И не только механизм доминирования, но и механизм действия и взаимодействия аллеломорфных пар вообще.
Казалось бы, с точки зрения рассмотрения генетически детерминированной активности фермента (или отсутствия активности), явление доминирования не представляет проблемы. Ферментативная активность должна доминировать над ее отсутствием.
У дрожжей Saccharomyces cerevisiae есть формы, наследственно различающиеся по окраске колоний: красные и белые. Красная пигментация - рецессивный признак. Она возникает вследствие генетического блока в биосинтезе пуринов: отсутствует активность фермента фосфорибозиламиноимидазолкарбоксилазы и поэтому дрожжи для своего роста нуждаются в экзогенном аденине. Субстрат реакции (аминоимидазолриботид) накапливается в клетке и конденсируется в красный пигмент. У белых дрожжей упомянутый фермент работает нормально, пигмент не накапливается и дрожжи не нуждаются в аденине. Они синтезируют его сами. Подобные примеры можно найти в описании любого метаболического пути, генетический контроль которого хорошо изучен.
Явление доминирования не исчерпывает все случаи взаимодействия аллелей. Уже упоминалось явление неполного доминирования. Кроме того, известны случаи отсутствия доминантно-рецессивных отношений или, точнее, случаи кодоминирования. Типичный пример такого взаимодействия аллелей - наследование антигенных группкрови человека: А, В, АВ и О, детерминируемых геномІ .
Известны три типа аллелей этого гена: І А , І в , і °. При гомозиготности І А І А эритроциты имеют только поверхностный антиген А (группа крови А, или II). При гомозиготности І в І в эритроциты несут только поверхностный антиген В (группа В, или III). В случае гомозиготности ІВ ІВ эритроциты лишены А и В антигенов (группа О или I). В случае гетерозиготности ІА іО или ІВ іО группа крови определяется, соответственно, А (II) или В (III). Эритроциты имеют, соответственно, антигены только А или только В. Это уже известный случай полного доминирования.
Если же человек гетерозиготен ІА І в , его эритроциты несут оба антигена: А и В (группа крови АВ, или IV). Это и есть случай кодоминирования. Аллели І А и І в работают в гетерозиготе как бы независимо друг от друга, что и определяют с помощью иммунологических методов.
Знание генетического контроля групп крови имеет большое практическое значение. Дело в том, что у людей с группой О в плазме крови присутствуют гемагглютинины α и β, с группой А - гемагглютинин β, с группой В - α. У людей группы АВ в плазме нет ни α- ни β-гемагглютининов. При этом агглютинин а специфически связывает и осаждает эритроциты с антигеном А, агглютинин β - эритроциты с антигеном В. На этих взаимоотношениях основана система переливания крови. Кровь группы О можно переливать всем людям, кровь группы А - людям с группами крови А и О, группы В - людям с группами В и О, а кровь группы АВ - только людям с той же группой. Нарушение этих правил приводит к геморрагическому шоку вследствие связывания эритроцитов гемагглютининами плазмы.
Знание этих закономерностей используется также в судебной медицине для идентификации пятен крови и при установлении отцовства.
Пример наследования групп крови иллюстрирует и проявление множественного аллелизма: ген І может быть представлен тремя разными аллелями, которые комбинируются в зиготах только попарно.
Явление множественного аллелизма широко распространено в природе. Известны обширные серии множественных аллелей, определяющих тип совместимости при опылении у высших растений, при оплодотворении у грибов, детерминирующих окраску шерсти животных, глаз у дрозофилы, форму рисунка на листьях белого клевера, наконец, у растений, животных и микроорганизмов известно много примеров так называемых аллозимов или аллельных изоэнзимов - белковых молекул, различия между которыми определяются аллелями одного гена.
Во многих случаях попарные взаимодействия членов серии аллелей приводят к тому, что исследуемый признак проявляется иначе, чем у гомозиготных родительских форм. В качестве примера можно привести наследование узора на листьях белого клевера (рис. 1). На рисунке показаны формы, гомозиготные по различным аллелям гена V (по вертикали), и формы, несущие разные сочетания этих аллелей.
В некоторых случаях механизм взаимодействия аллелей расшифрован. Вернемся к примеру с красными и белыми дрожжами. Существует большое число красных аденинзависимых мутантов дрожжей. Большинство из них несет изменения одного и того же гена. Во всех случаях потребность в аденине и красная окраска колоний рецессивны по отношению к белой окраске и, соответственно, к отсутствию потребности в аденине. Аллель, определяющая доминантный признак, обозначается, как это принято в генетике, прописными буквами: ADE - сокращенное наименование признака. Такую аллель условно называют нормальной или аллелью дикого типа. Поскольку путь биосинтеза аденина состоит из многих (двенадцати) этапов, каждый ген, контролирующий отдельный этап, имеет свой номер. Интересующий нас ген - ADE 2.

Рис. 1 - Множественная серия алелей, определяющих рисунок на листях белого клевера и их взаимодействие в гетерозиготе
Для обозначения аллелей этого гена, определяющих мутантный, рецессивный признак (рецессивных аллелей), используют те же буквы, но строчные - ade , а поскольку таких аллелей много, справа приписывают номер аллели: ade 2-1, ade 2-2 и т.д. В гомозиготном состоянии (или в гомозиготе) все они определяют характерный рецессивный или мутантный фенотип - красную окраску колонии и потребность в аденине вследствие отсутствия активности фермента фосфорибозиламиноимидазолкарбоксилазы.
В некоторых случаях при объединении в гибриде двух разных аллелей независимого происхождения, рецессивных по отношению к дикому типу, наблюдают восстановление нормы, т.е. признака дикого типа. При этом частично восстанавливается и ферментативная активность. Такое восстановление дикого фенотипа происходит весьма специфично - только в некоторых комбинациях аллелей.
Это загадочное, на первый взгляд, явление описано для многих объектов: дрозофилы, мышей, зеленой водоросли Chlamydomonas , многих грибов и т.п. Лучше всего оно изучено у микроорганизмов.
Примерно у 50 % генов, которые исследованы таким образом, обнаружен данный тип взаимодействия, получивший название межаллельной комплементации. Механизм его расшифрован. Оказалось, что все гены, аллели которых взаимодействуют подобным образом, контролируют структуру ферментов, построенных из одинаковых белковых субъединиц, т.е. одна и та же полипептидная цепь повторена в них несколько раз: от двух до восьми.
Если исследовать гомозиготы по рецессивным аллелям, то в этом случае в белковой молекуле повторяются одинаковые и одинаково испорченные субъединицы. А вот если изучать гибриды, гетерозиготные по разным рецессивным аллелям (такие гибриды также называют компаундами) : ade 2-1/ade 2-2; ade 2-1/ ade 2-3 и т.д., то в этом случае фермент будет содержать субъединицы, испорченные немного по-разному, и иногда эти субъединицы, взаимодействуя, приводят к восстановлению ферментативной активности. Как происходит это взаимодействие?
Для ответа на этот вопрос необходимо обратиться к современным представлениям о структуре белков, характере складывания в них непрерывной полипептидной цепи. В каждом белке есть несколько функциональных центров: для связывания субстрата, для взаимодействия с коферментами, с регуляторными молекулами, с мембранами клетки и т.д. Каждый функциональный центр обычно представлен полуавтономным участком специфически уложенной полипептидной цепи - доменом .
Разные рецессивные аллели одного и того же гена отличаются друг от друга тем, что кодируют полипептиды с повреждениями различных доменов. Если в гетерозиготе (компаунде) объединятся аллели с разными повреждениями таким образом, что в молекуле фермента, состоящей из субъединиц - продуктов одного и того же гена, соберутся все необходимые функциональные центры, то ферментативная активность будет восстановлена.
Итак, рассмотрев различные типы взаимодействия аллелей, отметим, что во всех случаях, где наблюдается та или иная форма доминирования, одну аллель можно рассматривать как активную, т.е. детерминирующую Проявление Признака, а другую - как неактивную. Лучше всего эти взаимоотношения иллюстрируют специфические функции белков - носителей элементарных признаков, или фенов. При этом следует подчеркнуть, что свойства доминантности - рецессивности относятся, строго говоря, не к самим аллелям, а к признакам. Таким образом, может быть принято рациональное зерно гипотезы «присутствия - отсутствия» У. Бэтсона, который предложил ее тогда, когда не только не было известно, как действуют гены, но и самого термина «ген» еще не существовало. По У. Бэтсону, рецессивные аллели представляют собой результат утраты доминантных аллелей. Теперь известно, что на самом деле мутации, приводящие к появлению рецессивных аллелей, далеко не всегда сопровождаются физической потерей генетического материала. Тем не менее они, как правило, возникают в результате утраты какой-либо нормальной функции.
Важно также отметить, что несмотря на различные и порой сложные по своим механизмам взаимодействия аллелей, которые уже рассмотрены, все они подчиняются первому закону Менделя - закону единообразия гибридов первого поколения.
3. Типы взаимодействия неаллельных генов
Закон независимого наследования генов еще раз демонстрирует дискретный характер генетического материала. Это проявляется в независимом комбинировании аллелей разных генов и в их независимом действии - фенотипическом выражении. Однако в ряде случаев идентификация фенов сопряжена с некоторыми трудностями. Обратимся к примеру.
3.1 Комплементарность
У популярного генетического объекта плодовой мушки Drosophila melanogaster имеется большое число форм, наследственно различающихся по окраске глаз. У мух так называемого дикого типа или типа, распространенного в природе, глаза темно-красные. Существуют формы с ярко-красными глазами. Этот признак рецессивен по отношению к дикому типу. Он наследуется по моногибридной схеме при скрещивании нормальных мух и мух с ярко-красными глазами. Соответствующий ген обозначается: st (scarlet) - рецессив; st + - доминант.
Существуют также мухи с коричневыми глазами. Это тоже рецессивный признак, наследующийся по моногибридной схеме при скрещивании диких мух и мух с коричневыми глазами. Соответствующий ген обозначается bw (brown) - рецессив; bw + - доминант1 .
Если скрестить мух с ярко-красными глазами и мух с коричневыми глазами, то получаются следующие результаты. В Fi все мухи имеют темно-красные глаза (дикий тип), а при скрещивании гибридов первого поколения в F 2 появляются четыре класса расщепления: мухи с темно-красными, ярко-красными, коричневыми и белыми глазами в соотношении 9:3:3:1. Для объяснения этого результата обратимся к логике генетического анализа.
Результаты, получившиеся в F1 , показывают, что существует некоторый тип взаимодействия. Можно предположить, что это взаимодействие аллелей одного гена при моногибридном скрещивании. Однако в F 2 появляются четыре класса в соотношении, характерном для дигибридного скрещивания при полном доминировании по обоим признакам, и среди них формируется самый малочисленный класс - мухи с белыми глазами.
Если справедливо предположение о том, что это скрещивание дигибридное, то генотипы родительских форм записывают:
![]()
где знак «+» соответствует нормальным (доминантным) аллелям генов: bw и st . Тогда самки и самцы образуют по одному типугамет - соответственно st bw + и st + bw , а генотип гибридов F1 будет: st + st bw + bw .
Такие дигетерозиготные мухи должны образовать четыре типа гамет, которые во всевозможных сочетаниях при скрещивании между собой гибридов F 1 дадут в F 2 следующее расщепление по генотипу.
С помощью фенотипических радикалов можно написать следующее расщепление по фенотипу:
9 st+ - bw + - - с темно-красными глазами
3 st+ - bw bw - с коричневыми глазами
3 st st bw + - - с ярко-красными глазами
1 st st bw bw - с белыми глазами
Учитывая, что аллели st и bw рецессивны, можно объяснить появление первых трех фенотипических классов при расщеплении. При наличии нормальных аллелей st + и bw + мухи должны принадлежать к дикому типу по окраске глаз (9 с темно-красными глазами). При гомозиготности только по рецессивной аллели bw bw мухи должны быть с коричневыми глазами (3), так же как при гомозиготности только по st st мухи должны быть с ярко-красными глазами (3). Наконец, остается последний класс - двойной гомозиготный рецессив (1 st st bw bw ), который соответствует мухам с белыми глазами. Все эти выводы можно проверить, исследуя далее расщепление при анализирующих скрещиваниях и скрещиваниях между особями F 1 .
Таким образом, предположение о дигибридном расщеплении в рассмотренном скрещивании подтверждается, а новообразование - белоглазые мухи в F 2 - результат взаимодействия рецессивных аллелей st и bw .
В рассмотренной схеме также наблюдалось взаимодействие генов в F 2 , в результате которого дрозофилы имели нормальный цвет глаз. Такой тип взаимодействия носит название комплементарности или комплементарного (взаимно дополнительного) действия, когда доминантные аллели обоих генов обусловили нормальный (или дикий) фенотип (под комплементарностью обычно подразумевают именно этот тип взаимодействия генов).
В F2 , рецессивные аллели тех же генов обусловили появление белоглазых мух. Был приведен пример того, что носит название формально-генетического анализа, при котором полностью абстрагируются от механизмов действия исследуемых генов. Если анализ проведен верно и формальные отношения генов и аллелей выявлены правильно, то последующее выяснение физиологических механизмов, лежащих в основе генных взаимодействий, только подтверждает и конкретизирует выводы.
Биохимический механизм взаимодействия аллелей генов st и bw исследован достаточно подробно. Известно, что у дрозофилы окраска глаз обусловлена синтезом двух пигментов - красного и бурого. Рецессивная аллель bw в гомозиготе прерывает синтез красного пигмента, поэтому глаза содержат только бурый пигмент. Рецессивная аллель st в гомозиготе блокирует синтез бурого пигмента, вследствие чего в глазах мух содержится только красный пигмент.
Когда в дигетерозиготе оказываются нормальные аллели обоих генов, синтезируются оба пигмента. Результат - комплементарное взаимодействие нормальных аллелей, наблюдаемое в F 1 . Если в F2 в гомозиготе оказываются и bwbw , и stst , то не синтезируются ни красный, ни бурый пигменты, и глаза оказываются белыми, неокрашенными.
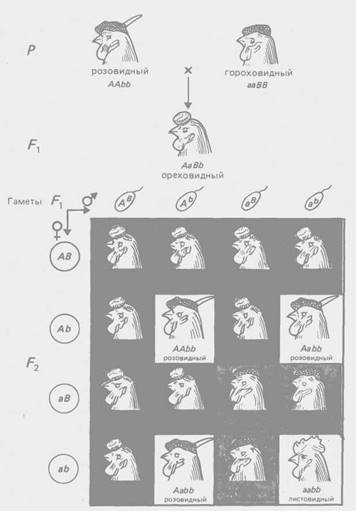
Рис. 2 - Комплементарное взаимодействие генов, определяющих форму гребня у кур (соотношение 9:3:3:1)
Можно рассмотреть и другой тип комплементарного взаимодействия генов у дрозофилы, если идти от метаболического эффекта генов к схеме их взаимодействия. Вновь обратимся к биосинтезу пигментов глаза у дорозофилы (рис. 3). Известно, что кроме рецессивных аллелей st синтез бурого пигмента блокируют и рецессивные аллели гена purple ( pr ). Фенотип гомозигот stst и ргрт - ярко-красные глаза. При скрещивании таких мух в F 1 глаза нормальные - темно-красные, поскольку работают оба гена - комплементарно взаимодействуют их доминантные аллели. В F 2 наблюдается следующее соотношение фенотипов: 9 с темно-красными и 7 с ярко-красными глазами. Это объясняется тем, что выход в гомозиготу любой из двух рецессивных аллелей достаточен для блокирования синтеза пигмента, тем более, когда и рг, и st находятся в гомозиготе. Это тоже пример комплементарного взаимодействия, но без новообразования в F 2 .
По типу комплементарное™ взаимодействуют гены, контролирующие разные этапы одного и того же метаболического пути. Однако для многих морфологических признаков неизвестен биохимический механизм их реализации, поэтому приходится ограничиваться констатацией формально-генетической схемы их наследования. Так, по типу комплементарности взаимодействуют гены, определяющие форму гребня кур (рис. 2), форму плода у тыквы (рис. 3) и др.
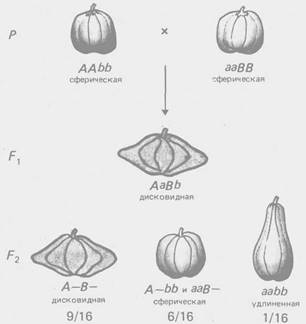
Рис. 3 - Комплементарное взаимодействие генов, определяющих форму плода тыквы (соотношение 9:6: 1)
3.2 Эпистаз
Вернемся к анализу взаимодействия генов р r и st у дрозофилы. Соотношение фенотипических классов в F 2 можно представить себе и как следствие того, что рецессивная аллель р r в гомозиготе препятствует проявлению доминантной аллели st + . Точно так же рецессивная аллель st в гомозиготе препятствует проявлению доминантной аллели р r + . Действительно, то, что известно о генетическом контроле синтеза бурого глазного пигмента у дрозофилы, вполне соответствует предложенному здесь объяснению. Такой тип взаимодействия носит название эпистатического, или эпистаза, и условно изображается: р r > st + и st > pr + . В данном случае рецессивная аллель р r эпистатична по отношению к доминантной аллели st + , ast эпистатична по отношению к р r + . Данный случай взаимодействия генов называют также двойным рецессивным эпистазом .
По изменению числа и соотношения классов дигибридного расщепления в F2 рассматривают несколько типов эпистатических взаимодействий: простой рецессивный эпистаз (а > В; а> Ь или b > А; Ь > а) , который выражается в расщеплении 9:3:4; простой доминаданый эпистаз (А> В; А> b или В > А; B > а) с расщеплением 12:3:1 и т.д.
Один ген, подавляющий действие другого, называют эпистатическим геном, ингибитором или супрессором. Подавляемый ген носит название гипостатического.
Как уже показано, констатация того или иного типа взаимодействия генов в дигибридном скрещивании условна. Тем не менее, при кажущемся нарушении закона независимого наследования (появлении неожиданных классов в расщеплении или уменьшении числа классов), связанного с взаимодействием двух генов, всегда можно свести наблюдаемые соотношения в F2 к классическому 9:3:3:1. При этом важно понять, какие классы объединились, и тогда интерпретировать тип взаимодействия.
Необходимо также отметить, что само словосочетание «взаимдействие генов» условно. В действительности взаимодействуют продукты генов, а не сами гены, так что правильнее было бы говорить о взаимодействии фенов, а не о взаимодействии генов. Отсюда понятно, что судить о том, с каким скрещиванием имеет дело экспериментатор: моногибридным, дигибридным или полигибридным - можно только на основании результатов полного гибридологического анализа.
3.3 Полимерия
Наряду с комплементарным и эпистатическим принято также рассматривать взаимодействие генов по типу полимерии. В этом случае разные гены как бы дублируют действие друг друга, и одной доминантной аллели любого из взаимодействующих генов достаточно для проявления изучаемой фенотипической характеристики. Так, при скрещивании растений пастушьей сумки с треугольными плодами (стручками) и с овальными плодами в F 1 образуются растения с плодами треугольной формы. При их самоопылении в F2 наблюдается расщепление на растения с треугольными и овальными стручками в соотношении 15:1. Это объясняется тем, что существуют два гена, действующих эднозначно. В этих случаях их обозначают одинаково (А1 и А2 ). Тогда все генотипы: А1 - А2 -, А1 - а2 а2 , а2 а2 А2 - будут иметь одинаковую фенотипическую характеристику - треугольные стручки, и только растения а1 а1 а2 а2 будут отличаться - образовывать овальные стручки. Это случай так называемый некумулятивной полимерии.
Однозначные, или полимерные, гены могут действовать и по типу кумулятивной полимерии. Так, шведский генетик Г. Нильсон-Эле в 1908 г. описал серию однозначно действующих генов, которые определяют окраску эндосперма зерен пшеницы. При этом интенсивность окраски зерен оказалась пропорциональной числу доминантных аллелей разных генов в тригибридном скрещивании. Наиболее окрашенными были зерна А1 А1 А2 А2 А3 А3 , а зерна а1 а1 а2 а2 а3 а3 не имели пигмента. Между этими крайними типами при расщеплении в F2 наблюдались промежуточные варианты в соотношении 1:6:15:20:15:6:1.
По типу кумулятивной полимерии наследуются многие количественные признаки, например цвет кожи у человека; молочность, яйценоскость, масса и другие признаки сельскохозяйственных животных; длина колоса у злаков, содержание сахара в корнеплодах сахарной свеклы и др. Изучением наследования таких признаков занимается специальный раздел генетики - генетика количественных признаков, которая важна прежде всего для селекции и разработки проблем микроэволюции.
3.4 Гены-модификаторы
Основателем генетики количественных признаков в нашей стране был Ю.А. Филипченко. Он изучал наследование размеров черепа крупного рогатого скота, длины колоса пшеницы и даже умственных способностей у человека. В одной из работ 1928 г. он опубликовал данные о наследовании длины колоса при скрещивании двух форм пшеницы. В F 2 он наблюдал распределение по длине колоса, хорошо согласующееся с гипотезой о моногенном различии по этому признаку. Однако последующий анализ показал, что наряду с «основным» геном, определяющим длину колоса, существует ряд генов-модификаторов этого признака. Подобный тип наследования встречается часто. Таким образом, фенотип, как правило, представляет собой результат сложного взаимодействия генов. Природа генов-модификаторов до сих пор вызывает споры: в частности, не ясно, существуют ли специальные модификаторы, функция которых заключается в изменении действия других - «основных» генов или модифицирующее действие гена - результат его плейотропии.
4. Плейотроп ное действие генов
Учитывая данные, изложенные в этой главе, следует заключить, что не бывает однозначного соотношения между генотипом и фенотипом. Справедливость этого положения подчеркивает и тот факт, что один и тот же ген может в конечном итоге действовать на различные признаки организма.
Первый пример такого множественного, или плейотропного, действия гена содержится в работе Менделя, а именно: окраска цветков и окраска семенной кожуры зависели в его опытах от одного наследственного задатка. У высших растений гены, обусловливающие красную (антоциановую) окраску цветков, одновременно контролируют красную окраску стебля. У человека известен доминантный ген, определяющий признак «паучьи пальцы» (арахнодактилия или синдром Марфана). Одновременно он определяет аномалии хрусталика глаза и порок сердца. В Западном Пакистане обнаружены люди - носители гена, определяющего отсутствие потовых желез на отдельных участках тела. Это одновременно определяет и отсутствие некоторых зубов.
Признак платиновой окраски шерсти у лисиц контролируется доминантным геном, который существует только в гетерозиготе, поскольку обладает рецессивным летальным действием. При скрещивании платиновых лис наблюдали расщепление на платиновых и серебристо-черных в соотношении 2:1. Такое соотношение может получаться, если платиновые лисицы гетерозиготны (Аа), а черные гомозиготны по рецессивной аллели того же гена (аа). При этом не выживают гомозиготы по доминантной аллели (АА). Такое предположение подтверждается результатами скрещивания платиновых и серебристо-черных лис. Как и следует ожидать, при анализирующем скрещивании получается расщепление на платиновых и серебристо-черных в отношении 1:1. По этой же схеме наследуется наличие (аа) и отсутствие (Аа) чешуи у зеркального карпа, серая (Аа) и черная (аа) окраска каракулевых овец и т.д.
Множественное или плейотропное действие генов связывают с тем, на какой стадии онтогенеза проявляются соответствующие аллели. Чем раньше проявится аллель, тем больше эффект плейотропии.
Учитывая плейотропный эффект многих генов, можно предположить, что часто одни гены выступают в роли модификаторов действия других генов.
5 . Пенетрантность, экспрессивность, норма реакции
Рассматривая действие гена, его аллелей, необходимо учитывать не только генные взаимодействия и действие генов-модификаторов, но и модифицирующее действие среды, в которой развивается организм. Известно, что у примулы окраска цветка розовая (Р-) - белая (рр) наследуется по моногибридной схеме, если растения развиваются в интервале температур 15-25°С. Если же растения F 2 вырастить при температуре 30-35°С, то все цветки у них оказываются белыми. Наконец, при выращивании растений F 2 в условиях температуры, колеблющейся около 30°С, можно получить разнообразные соотношения от 3 Р :1рр до 100% растений с белыми цветками. Такое варьирующее соотношение классов при расщеплении в зависимости от условий внешней среды или от условий генотипической среды (так назвал С.С. Четвериков варьирование генотипа по генам-модификаторам) носит название варьирующей пенетрантности: Это понятие подразумевает возможность проявления или непроявления признака у организмов, одинаковых по исследуемым генотипическим факторам.
Уже упоминался пример плейотропного действия гена - доминантная платиновая окраска лисиц с рецессивным летальным действием. Как показал Д.К. Беляев с сотрудниками, можно добиться рождения живых щенков, гомозиготных по доминантной аллели платиновой окраски, если варьировать длину дня для беременных самок. Таким образом, пенетрантность проявления летального эффекта может быть снижена (уже не будет 100%-ной).
Пенетрантность выражается долей особей, проявляющих исследуемый признак среди всех особей одинакового генотипа по контролируемому (изучаемому) гену.
От внешней среды и генов-модификаторов может зависеть и степень выраженности признака. Например, дрозофила, гомозиготная по аллели vgvg (зачаточные крылья), более контрастно проявляет этот признак при понижении температуры. Другой признак дрозофилы - отсутствие глаз (еуеу) варьирует от 0 до 50% от числа фасеток, характерного для мух дикого типа.
Степень проявления варьирующего признака называется экспрессивностью. Экспрессивность обычно выражают количественно в зависимости от уклонения признака от дикого типа.
Оба понятия - пенетрантность и экспрессивность - были введены в 1925 г. Н.В. Тимофеевым-Ресовским для описания варьирующего проявления генов (рис. 4).

Рис. 4 - Схема, поясняющая варьирование экспрессивности и пенетрантностипризнака
Тот факт, что признак может проявиться или не проявиться у особей данного генотипа в зависимости от условий или варьировать в различных условиях среды, убеждает в том, что фенотип - это результат действия (и взаимодействия) генов в конкретных условиях существования организма.
Способность генотипа так или иначе проявляться в различных условиях среды отражает норму его реакции - способность реагировать на варьирующие условия развития. Норму реакции генотипа необходимо учитывать как при экспериментах, так и при выведении новых форм хозяйственно ценных организмов. Отсутствие изменений в проявлении признака указывает на то, что используемое воздействие не влияет на данную норму реакции, а гибель организма - на то, что оно уже за пределами нормы реакции. Селекция высокопродуктивных форм растений, животных и микроорганизмов в значительной степени представляет собой отбор организмов с узкой и специализированной нормой реакции на такие внешние воздействия, как удобрения, обильное кормление, характер выращивания и др.
Искусственное сужение или сдвиг нормы реакции используют для маркирования многих жизненно важных генов. Так, были исследованы гены, контролирующие воспроизведение ДНК и синтез белка у бактерий и дрожжей, гены, контролирующие развитие дрозофилы, и др. При этом получали мутанты, нежизнеспособные при повышенной температуре культивирования, т.е. условно-летальные.
Таким образом, материал, рассмотренный в этой главе, показывает, что генотип представляет собой систему взаимодействующих генов, которые проявляются фенотипически в зависимости от условий генотипической среды и условий существования. Только благодаря использованию принципов менделеевского анализа можно условно разложить эту сложную систему на элементарные признаки – фены и тем самым идентифицировать отдельные, дискретные единицы генотипа – гены.
6. Влияние факторов внешней среды на действие генов
Влияние внешней среды: условий питания, температуры, света, химического состава и структуры почвы, влажности и т.п. - на индивидуальное развитие организма огромно. Человеку, занимающемуся сельскохозяйственным производством, очевидна роль внешней среды в формировании организмов. Внешняя среда определяет процессы отбора, изменчивости и наследственности, т.е. факторы эволюции. В соответствии с изменениями внешней среды развивался весь органический мир. Благодаря этому каждый вид организма приобрел своеобразное внутреннее строение и присущий ему тип индивидуального развития, закрепленные наследственностью.
Поскольку у организмов разных видов благодаря совместному действию указанных трех факторов эволюции выработались приспособления к определенным условиям внешней среды, их нормальное развитие стало возможно только при данных условиях. Изменение последних может приводить к изменению наследственных факторов (мутации), с чем мы познакомимся позднее, и к изменению процессов индивидуального развития, вызывающему видоизменение фенотипа, т.е. к изменению проявления действии генов. Реализация наследственного признака или свойства организма является результатом взаимодействия генотипа и условий внешней среды.
Создавая определенные внешние условия, можно направлять действие генов в нужную нам сторону, т.е. управлять процессами индивидуального развития организма. Генетики давно обратили на это внимание. Проблема управления индивидуальным развитием привлекла внимание И.В. Мичурина. Он показал, что наследственная потенция организма огромна, и, подставляя соответствующие внешние условия для развития гибридов плодовых и ягодных растений, можно изменять доминирование и выявлять наиболее ценные наследственные качества и свойства организма.
По этому поводу И.В. Мичурин в 1929 г. писал: «В общем влияние всей совокупности внешних факторов на строение организма гибридов настолько велико, что в большинстве случаев значительно подчиняет себе действия наследственной передачи качеств и свойств растений-производителей. В особенности такое влияние резко отражается на состоянии материнского растения при закладке у него в строении семени зачатков будущего организма гибрида и на полученном гибриде в самой ранней стадии его развития, благоприятствуя одним и являясь непреодолимым препятствием для проявления других наследственных признаков». При этом И. В. Мичурин добавлял, что «в законе Менделя нисколько не отвергаю его достоинств, напротив, я лишь настаиваю на необходимости внесения в него поправок и дополнений...».
Одним из ярких примеров влияния внешней среды на действие гена и выражение признака является изменение доминирования. Так, И.В. Мичурин на гибриде груши маслинолистной (Pyruselaeagnifolia) и сорта Бессемянка и на гибриде вишен Самарской и Владимирской показал, что в гибридном организме будут доминировать те свойства и признаки, которые встречают наиболее благоприятные условия для своего осуществления. Для той же цели управления доминированием им применялся «спартанский режим воспитания» гибридных сеянцев, а также метод ментора.
Изучение влияния внешних факторов на действие отдельных генов у животных также показало возможность изменять фенотип организма. Так, например, встречаются кролики с желтым и белым жиром. Кролики с желтым жиром имеют в своем генотипе рецессивную аллель у в гомозиготном состоянии. Наличие доминантной аллели гена У определяет белый жир. Этот ген контролирует выработку энзима, который разрушает желтый пигмент - ксантофилл, содержащийся в зеленом корме. Если кроликам с желтым жиром (по генотипу уу) давать корм, который не содержит ксантофилла, то у них будет образовываться белый жир, так как для реализации действия гена не будет создано подходящих условий.
У гибридов от скрещивания озимых и яровых форм пшеницы или ржи в первом поколении доминирует свойство яровости. Во втором поколении у ржи наблюдается расщепление в отношении 3:1, у пшениц могут быть расщепления 3:1 и 15:1 и более сложные. Если семена второго поколения посеять весной, то озимые формы будут куститься, но не будут колоситься, что даст возможность обнаружить расщепление. Если же эти семена предварительно прояровизировать, т.е. подвергнуть предпосевной обработке низкими температурами, то все растения второго поколения будут колоситься в год посева. В этом случае мы не обнаружим расщепления.
У одноклеточной зеленой водоросли хлореллы известны наследственные изменения, обусловливающие отсутствие хлорофилла. Эти формы не могут жить и размножаться в обычных условиях на свету, так как они не способны осуществлять фотосинтез. Но если в питательную среду для таких форм добавить глюкозу, то они смогут размножаться. Подобные опыты проводились и с бесцветными альбинотическими проростками кукурузы.
Можно твердо сказать, что бессмысленно представлять себе проявление наследственности вне конкретных условий внешней среды. Знание механизмов действия гена и особенностей влияния внешней среды на его проявление обеспечивает возможность управления развитием врожденных свойств. Благодаря этому развитие наследственных заболеваний у человека перестает быть фатальной неизбежностью. Изучение влияния внешних условий на породные и сортовые (т.е. наследственные) хозяйственно ценные свойства организмов расширяет возможности повышения продуктивности животных, растений и микроорганизмов.
Выводы
Итак, подводя итоги нашей работы можно сказать, что генетический анализ наследования и взаимодействия отдельных генов у организмов независимо от их филогенетического уровня и сложности организации служит необходимым начальным приемом, соответствующим качественному и количественному анализу в химии. Когда исследователь имеет дело с неизвестным ему химическим веществом, он в определенной последовательности исследует сначала группы элементов, а затем сами элементы и, наконец, их количественное соотношение.
Однако следует иметь в виду, что сам по себе генетический анализ взаимодействия генов является формальным, так как наследственные признаки используются в нем лишь в качестве «маркеров»для прослеживания судьбы генов в ряду поколений и их взаимосвязи на «финише» индивидуального развития, т.е. в фенотипе. Взаимодействие генных продуктов при формировании признаков данному анализу оказывается недоступным.
В ходе изложения неоднократно обращалось внимание на то, что все перечисленные закономерности поведения генов в хромосомах оказываются общими для животных и человека, для высших и низших растений, а также для одноклеточных организмов.
Из анализа расщепления, независимого комбинирования генов, а также их взаимодействия видно, что комбинация генов и их взаимодействие создают огромную наследственную изменчивость. В результате комбинации генов могут возникать новые наследственные признаки. Такого типа изменчивость называется комбинативной изменчивостью. Она играет существенную роль эволюционном процессе как источник изменчивости, как механизм, обеспечивающий сочетание наиболее приспособительных признаков для существования организмов.
Список использованной литературы
1. Актуальные вопросы современной генетики. /Под ред. С.И. Алиханяна. Изд. МГУ, 1966.
2. Алиханян С.И., Акифъев А.П., Чернил Л.С. Общая генетика. М., 1985.
3. Биологический энциклопедический словарь. М., 1986.
4. Ватти К.В., Тихомирова М.М. Руководство к практическим занятиям по генетике. М., 1979.
5. Гуляев Г.В. Генетика. М., 1977.
6. Дубинин Н.П. Генетика. Кишенев, 1985.
7. Инге-Вечтомов С.Г. Введение в молекулярную генетику. М., 1983.
8. Медведев Н.Н. Практическая генетика. М., 1968.
9. Лобашев М.Е. Генетика. Изд. ЛГУ, 1967.
10. Лобашев М.Е., Ватти К.В., Тихомирова М.М. Генетика с основами селекции. М., 1979.
11. Морган Т. Структурные основы наследственности. Москва - Петроград, 1924.
12. Натали В.Ф. Основные вопросы генетики. М., 1967.
13. Рокицкий П.Ф. Введение в статистическую генетику. Минск, 1978.
14. Серебровский А.С. Генетический анализ. М., 1970.
15. Уильяме У. Генетические основы и селекция растений. М., 1968.
16. Филипченко Ю.А. Генетика. Госиздат, 1929.
Похожие рефераты:
Литература - Другое (книга по генетике)
Аллельные варианты генов-кандидатов подверженности туберкулезу у русского населения Западной Сибири
Математические модели естествознания
Шпаргалки (биология) для выпускных экзаменов в 11 классе
Закономерности передачи генетической информации
Психолого-педагогические аспекты изучения генетики в школе
Что такое ген? Генетическая точка зрения
Генотипическое и фенотипическое разнообразие населения Украины по группам крови