| Скачать .docx | Скачать .pdf |
Курсовая работа: Межклеточные контакты
ФЕДЕРАЛЬНОЕ АГЕНСТВО ПО ОБРАЗОВАНИЮ
ПЕНЗЕНСКИЙ ГОСУДАРСТВЕННЫЙ ПЕДАГОГИЧЕСКИЙ УНИВЕРСИТЕТ ИМ. В.Г. БЕЛИНСКОГО
Естественно-географический факультет
Кафедра биохимии
КУРСОВАЯ РАБОТА
ПО БИОХИМИИ МЕМБРАН:
«МЕЖКЛЕТОЧНЫЕ КОНТАКТЫ»
Выполнила студентка IV курса группы БХ-41
Гребенникова Ирина
Научный руководитель Соловьев В.Б.
ПЕНЗА, 2009г.
Содержание:
Введение
1.Классификация межклеточных контактов
2. Замыкающие межклеточные контакты
2.1.Рыхлые или простые контакты
2.2.Плотный замыкающий контакт
2.3.Замыкающие межклеточные контакты
3.Адгезионные или сложные межклеточные контакты
3.1.Точечные межклеточные контакты
3.2.Адгезионные пояски
3.3.Адгезионные соединения между клеткой и матриксом
3.4.Десмосомы
4.Гемидесмосома
5.Заболевание аутоиммунное, поражающее десмосомы
6.Проводящие межклеточные контакты
6.1.Синапсы
6.2.Нексусы
7.Рецепторы
Вывод
Приложение 1
Список литературы
Введение
Межклеточные контакты возникают в местах соприкосновения клеток в тканях и служат для межклеточного транспорта веществ и передачи сигналов, а также для механического скрепления клеток друг с другом. Основные типы межклеточных контактов: а) рыхлые, или простые, контакты — между плазматическими мембранами соседних клеток имеется щель шириной 10—20 нм, заполненная гликокалликсом, специализированных структур на мембранах нет; б) межклеточные «замки» — мембраны соседних клеток разделены таким же расстоянием, но изгибаются, образуя на поверхности клеток впячивания; в) десмосомы; г) плотные контакты (встречаются в основном в эпителиальных клетках) — разделяются на зону замыкания и зону слипания (промежуточный контакт); в зоне замыкания две соседние мембраны сливаются своими наружными слоями, эта зона непроницаема для макромолекул и ионов, в зоне слипания мембраны разделены щелью в 10—20 нм, заполненной плотным веществом, вероятно, белковой природы; д) щелевидные (высокопроницаемые) контакты, свойственные всем типам эпителиальной и соединительной тканей,— плазматической мембраны разделены промежутком в 2— 4 нм, пронизанным каналами, по которым низкомолекулярные вещества попадают из цитоплазмы одной клетки в другую, минуя межклеточную среду. В большинстве случаев межклеточные контакты разрушаются при удалении из среды ионов Са2+ . Особыми формами межклеточных контактов являются синапсы, а также плазмодесмы растит, клеток.
1. Классификация межклеточных контактов
1.Замыкающие межклеточные контакты.
а) простой или рыхлый контакт;
б) плотный замыкающий контакт.
2.Адгезионные межклеточные контакты.
а) точечные контакты;
б) адгезионные пояски;
в) адгезионные соединения между клеткой и внутриклеточным матриксом;
г) десмосомы.
3.Проводящие.
а) нексусы;
б) синапсы.
2. Замыкающие межклеточные контакты
2.1 Рыхлые или простые контакты
Простой контакт — соединение клеток за счет пальцевидных впячиваний и выпячиваний цитомембран соседних клеток. Специфических структур, формирующих контакт, нет.
Простые контакты занимают наиболее обширные участки соприкасающихся клеток. Расстояние между билипидными мембранами соседних клеток составляет 15-20 нм, а связь между клетками осуществляется за счет взаимодействия макромолекул соприкасающихся гликокаликсов.
Посредством простых контактов осуществляется слабая механическая связь - адгезия, не препятствующая транспорту веществ в межклеточных пространствах. Разновидностью простого контакта является контакт "типа замка", когда плазмолеммы соседних клеток вместе с участком цитоплазмы как бы впячивается друг в друга (интердигитация), чем достигается большая поверхность соприкосновения и более прочная механическая связь.
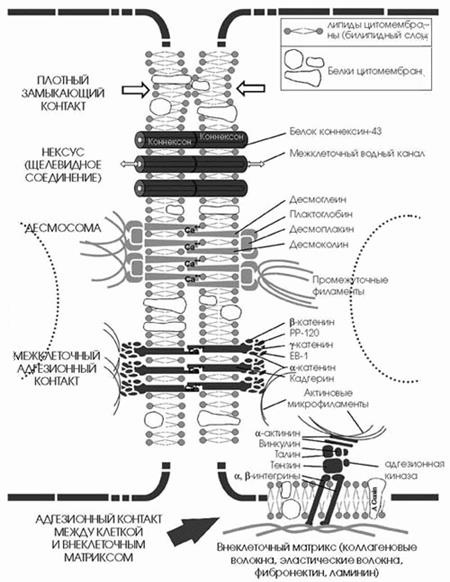
2.2 Плотный замыкающий контакт
Плотный замыкающий контакт — соприкасаются билипидные слои мембран соседних клеток. В области зоны плотных контактов между клетками не проходят практически никакие вещества.
Постоянные клеточные контакты скрепляют клетки в эпителиальном клеточном слое таким образом, что предотвращается перетекание даже малых молекул с одной стороны слоя на другую. Латеральная подвижность многих мембранных белков ограничена. Ограничение подвижности достигается с помощью барьеров, образованных при участии плотных контактов.
Клоны эпителиальных тканей (эпителии) функционируют в качестве избирательно-проницаемых барьеров, разделяющих жидкости с разным химическим составом по обе стороны слоя. В выполнении этой функции плотные контакты играют две роли.
Осуществляемый эпителиальными клетками трансклеточный транспорт (например, питательных веществ полости тонкого кишечника во внутриклеточную жидкость по другую сторону слоя) зависит от двух групп мембранных белков-переносчиков: одна находится на апикальной (обращенной в полость) поверхности клетки и активно транспортирует отдельные молекулы в клетку; другая находится на базолатеральной поверхности клетки и позволяет тем же молекулам покидать клетку путем облегченной диффузии . Для поддержания этого направленного транспорта не должно происходить перемещения апикальных белков-переносчиков на базолатеральную поверхность и наоборот.
Кроме того, промежутки между эпителиальными клетками должны быть скреплены таким образом, чтобы транспортированные молекулы не могли бы продиффундировать назад в полость через межклеточные промежутки.
Плотные контакты и выполняют эти две функции: барьеров для диффузии мембранных белков между апикальной и базолатеральной поверхностями и скрепления соседних клеток вместе так, что водорастворимые молекулы не могут перетечь на другую сторону слоя. При этом плотные контакты непроницаемы для макромолекул, а их проницаемость для малых молекул сильно варьирует в разных эпителиях. Эпителиальнные клетки могут временно модифицировать плотные контакты с тем, чтобы допустить увеличенный ток жидкости через бреши в контактных барьерах. Такой параклеточный транспорт особенно важен при абсорбции аминокислот и моносахаридов из полости тонкого кишечника.
Важнейшим элементом в структуре избирательно проницаемых барьеров эпителиальных и эндотелиальных являются плотные контакты. Избирательная проницаемость варьирует от ткани к ткани, пропуская или целые клетки и макромолекулы, или только протоны и ионы. Плотный контакт выглядит как пояс из переплетающихся скрепляющих нитей, который полностью окружает апикальный конец каждой клетки эпителиального слоя. Полагают, что скрепляющие нити состоят из длинных рядов специфических трансмембранных белков в каждой из двух взаимодействующих плазматических мембран, и которые (белки) соединяются напрямую друг с другом, что приводит к закупориванию межклеточного пространства. Интегральным мембранным белком плотного соединения оказался окклудин (взаимодействует с двумя цитоплазматическими белками, ZO-1 и ZO-2 ( zonula occludence 1, 2 ). Их функция окончательно не ясна. Возможно, их роль заключается в локализации оккулдина в сайтах между апикальной и базолатеральной поверхностями клетки. Некоторые ассоциированные с цитоскелетом белки были также обнаружены в участках плотных контактов. Среди них зингулин, антиген и актин (по данным электронной микроскопии, актиновые филаменты состоят из двух цепей глобулярных молекул, диаметром 4 нм и образующих двойную спираль, на каждый виток которой приходится 13,5 молекулы). Эти цепи составляют основу тонких филаментов скелетных мышц, которые кроме актина содержат также несколько других белков; глобулярный актин имеет молекулярную массу около 42 кД. Он содержит одну полипептидную цепь, состоящую из 375 или 374 аминокислотных остатков; различия в аминокислотной последовательности у разных актинов, как в пределах одного вида, так и межвидовые, крайне незначительны. Они составляют не более 25 аминокислотных замен; в настоящее время у позвоночных животных различают 6 изоформ актина, в зависимости от изоэлектрической точки они делятся на 3 класса - альфа, бета и гамма; бета- и гамма-актины характерны для немышечных клеток , а альфа-актины - для мышечных). Ras играет определенную роль в регулировании функционирования плотных соединений. Таким образом, в клетках имеются, по-видимому, сходные механизмы построения и регуляции адгезионных структур, и эти механизмы тесно взаимосвязаны с изменениями в цитоскелете. Однако, каким образом перестройки цитоскелета влияют на процессы межклеточной адгезии, пока окончательно не ясно. Механизмы адгезии и межклеточной сигнализации тесно сопряжены с давно известным феноменом контактного торможения , природа которого до сих пор до конца не выяснена.
2.3 Замыкающие межклеточные контакты
Простое межклеточное соединение — сближение плазмолемм соседних клеток на расстояние 15—20 нм. При этом происходит взаимодействие слоев гликокаликса соседних клеток. Гликопротеиды соседних клеток при образовании простого контакта «узнают» клетки одного типа. Наличие этих белков-рецепторов (кадгерины, интегрины и др.) характерно для определенных тканей. Они реагируют только с соответствующими им клетками. Например, кадгерины участвуют в образовании контактов только между эпителиальными клетками, обеспечивая их соединение практически по всей поверхности контактирующих клеток.
Интегрины - представляют собой поверхностные гетеродимерные белки, которые обеспечивают адгезию клеток к компонентам внеклеточного матрикса и иногда к другим клеткам. Многие интегрины проявляют сродство к гликопротеидам и базальной мембраны, и внеклеточного матрикса. Утрата некоторых интегринов (при раке молочной железы, раке предстательной железы, раке толстой кишки) или их избыток (при меланоме, плоскоклеточном раке полости рта, носоглотки, гортани) сопряжены с высокой степенью злокачественности опухоли. Связывание интегринов с лигандами и сближение клеток необходимы для перестройки базальной мембраны, идущей при ангиогенезе . Взаимодействие интегринов с белками внеклеточного матрикса в некоторых случаях препятствует апоптозу . Так, клетки меланомы избегают апоптоза в дерме за счет связывания альфа(ню)-бета3-интегрина с коллагеном. Нейтрализация этого интегрина антителами, напротив, способствует апоптозу. Таким образом, информация, которую интегрины передают от внеклеточного матрикса внутрь клетки, в одних случаях стимулирует адгезию и миграцию опухолевых клеток, в других - приводит к их гибели. Иными словами, интегрины играют роль своеобразного "переключателя", определяющего дальнейшую судьбу опухолевой клетки.
Интегрины - семейство родственных белков с молекулярной массой 100-160 кД, способных узнавать в матриксных белках пептид RGD(аминокислотная последовательность ARG-GLY-ASP, узнаваемая в белках интегринами, встречается в белках клеточных мембран, внеклеточного матрикса и т.д.).
Это большое семейство трансмембранных линкерных белков, являющихся расположенными на клеточной поверхности рецепторами большинства белков внеклеточного матрикса, включая коллаген, фибронектин, витронектин, ламинин . Одновременное множественное, но слабое связывание интегринов со своими лигандами позволяет клеткам исследовать свое окружение, сохраняя способность двигаться, что было бы невозможно при слишком прочных взаимодействиях. Интегрины работают как рецепторы клеток и ЕСМ белков. Клеточное соединение с помощью интегринов быстрое - в течение минут.
Интегрины - это молекулы межклеточной адгезии, которые присутствуют на поверхности различных клеток, в том числе и лейкоцитов. Они участвуют в адгезии лейкоцитов к внеклеточному матриксу и к эндотелию. Все белки, входящие в это крупное семейство, состоят из двух нековалентно связанных полипептидых цепей (альфа и бета). Обе цепи пронизывают клеточную мембрану. Альфа цепь содержит 3 или 4 тандемных повтора мотива связывающего двухвалентные ионы и нуждаются в Mg и Ca для функционирования. Альфа цепи при связывании с бета цепью дают функциональный рецептор. Бета цепь имеет функциональное значение и интегрины классифицируются по ним. Так интегрины с бета 1 или бета 3 цепью преимущественно вовлечены во взаимодействие клетки - ЕСМ. Интегрины с бета 2 цепью преимущественно вовлечены во взаимодействие лейкоцитов между собой. Семейство интегринов делят на три основных подсемейства по типу бета-цепи (бета1, бета2 и бета3). Тип альфа-цепи не так важен для функциональной активности.
Клеточно-матриксные контакты, образованные с помощью интегринов хорошо изучены в гладкомышечных клетках и в местах прикрепления культивируемых фибробластов к внеклеточному матриксу.
Кадгедрин. На наружной поверхности плазматической мембраны гладкомышечных клеток (ГМК), выделенных из кровеносных сосудов человека, выявлен белок, который осуществляет Са-зависимое гомофильное межклеточное взаимодействие. Этот белок очищен до гомогенного состояния, подвергнут ограниченному протеолизу, определена первичная структура полученных пептидов. Установлено, что он является рецептором межклеточного взаимодействия Т-кадгерином, относящимся к семейству фосфатидилинозитолгликан-заякоренных белков. Обнаружено, что в ГМК Т-кадгерин локализован в кавеолах вместе с Src-киназой и α-субъединицами G-белков. Показано, что Т-кадгерин способен связывать липопротеиды низкой плотности, а антитела к Т-кадгерину подавляют это связывание. При связывании липопротеидов ослабляется взаимодействие ГМК, стимулируется их пролиферация и синтез белка. Обнаружена стимуляция липопротеидами фосфоинозитидного обмена и выхода Са2+ из саркоплазматического ретикулума в цитоплазму. Все эти регуляторные эффекты липопротеидов подавляются коклюшным токсином, который АДФ-рибозилирует в ГМК β-субъединицы Gi2 - и Gi3 -белков. Экспрессия Т-кадгерина в ГМК снижается при повышении плотности клеток в культуре. Приведенные данные свидетельствуют в пользу того, что нами обнаружен и выделен новый рецептор гомофильного межклеточного взаимодействия, который также способен связывать липопротеиды низкой плотности. При взаимодействии с этим рецептором липопротеиды запускают зависимую от G-белка внутриклеточную сигнализацию. В результате этого связывания частично устраняется также контактное торможение деления клеток, что приводит к активации их деления и подавлению экспрессии Т-кадгерина.
3. Адгезионные или сложные межклеточные контакты
Сложные или адгезионные межклеточные соединения представляют собой небольшие парные специализированные участки плазматических мембран двух соседних клеток. Они подразделяются на запирающие (изолирующие), сцепляющие (заякоривающие) и коммуникационные (объединяющие) контакты.
К запирающим (изолирующим) относится плотный контакт (запирающая зона — zona occuludens). В этом соединении принимают участие специальные интегральные белки, расположенные на поверхности соседних клеток, образующие подобие ячеистой сети. Эта ячеистая сеть окружает в виде пояска весь периметр клетки, соединяясь с такой же сетью на поверхности соседних клеток. Эта область непроницаема для макромолекул и ионов и, следовательно, она запирает, отграничивает межклеточные щели (и вместе с ними собственно внутреннюю среду организма) от внешней среды. Этот тип соединений характерен для клеток однослойных эпителиев и эндотелия.
К сцепляющим, или заякоривающим, соединениям относятся адгезивный (сцепляющий) поясок и десмосомы. Общим для этой группы соединений является то, что к участкам плазматических мембран со стороны ци-топлазмы подходят фибриллярные элементы цитоскелета, которые как бы заякориваются на их поверхности.
Адгезивный (сцепляющий) поясок — парное образование в виде ленты, опоясывающей апикальную часть клетки однослойных эпителиев. Здесь клетки связаны друг с другом интегральными гликопротеидами, к которым со стороны цитоплазмы и той и другой клетки примыкает слой примембранных белков, включающих характерный белок винкулин. К этому слою подходит и связывается с ним пучок актиновых микрофиламентов. Кооперативное сокращение актиновых микрофиламентов во многих соседствующих клетках может привести к изменению рельефа всего эпителиального пласта.
К сцепляющим соединениям может быть отнесен так называемый фокальный контакт, характерный для фибробластов. В этом случае клетка соединяется не с соседней клеткой, а с элементами внеклеточного субстрата. В образовании фокального контакта также принимают участие актиновые микрофиламенты. К заякоривающим межклеточным соединениям относятся и десмосомы. Это тоже парные структуры, представляющие собой небольшую площадку или пятно диаметром около 0,5 мкм. Со стороны цитоплазмы к плазматической мембране прилежит слой белков, в состав которого входят десмоплакины. В этом слое заякореваются пучки цитоплазматических промежуточных филаментов. С внешней стороны плазмолеммы соседних клеток в области десмосом соединяются с помощью трансмембранных доменов белков — десмоглеинов. Каждая клетка эпидермиса кожи может иметь до нескольких сотен десмосом.
Функциональная роль десмосом заключается главным образом в механической связи между клетками. Десмосомы связывают друг с другом клетки в различных эпителиях, в сердечных и гладких мышцах. Полудесмосомы связывают эпителиальные клетки с базальной мембраной.
3.1 Точечные межклеточные контакты
Точечные — контакт образуется на небольшом по площади участке цитомембран соседних клеток.
3.2 Адгезионные пояски
Адгезионные пояски — контакт окружает по периметру всю клетку в виде пояса, располагается в верхних отделах боковых поверхностей эпителиальных клеток.
В области контакта в цитомембрану встроены специальные трансмембранные белки — кадгерины, которые соединяются с кадгеринами другой клетки. Для соединения кадгеринов нужны ионы кальция.
Со стороны цитоплазмы к кадгеринам присоединяются белки, β-катенин, α-катенин, γ-катенин, PP-120, EB-1, и к ним присоединяются актиновые микрофиламенты.
3.3 Адгезионные соединения между клеткой и матриксом
Контакт образуется на небольшом по площади участке. В месте контакта в цитомембрану встроены трансмембранные белки α- и β-интегрины, которые соединяются с элементами межклеточного матрикса.
Со стороны цитоплазмы к интегринам присоединяются несколько промежуточных белков (тензин, таллин, α-актинин, винкулин, паксиллин, фокальная адгезионная киназа), к которым присоединяются актиновые микрофиламенты.
Винкулин - молекула белка размером 1066 аминокислот (что характерно для человека и цыпленка) содержит 2 домена, которые получаются при протеолизе белка. N-концевой глобулярный домен, 95кДа. С-концевой удлиненный домен, 30кДа. Обе части белка соединены пролин-богатым участком длиной 41 аминокислота, где и находятся 2 сайта действия протеазы. Винкулин-α и Винкулин-β - изоформы винкулина с молекулярной массой 130 кДа , названные метавинкулином , который экспрессируется в гладких, поперечно-полосатых и сердечной мышцах. Обе формы получаются путем альтернативного сплайсинга 19 экзона. Он присутствует в метавинкулине между 915 и 916 положением γ- винкулина.
Таллин - связывается непосредственно с цитоплазматическим доменом интегрина. Субъединица талина с массой 215-235 кД димеризуется с образованием длинной гибкой молекулы. На молекуле имеется сужение, чувствительное к Са-зависимой протеазе II. При этом талин разрезается на два неравных фрагмента - 200кД и 47 кД. Участок связывания интегрина находится на малом фрагменте. На большом фрагменте находится участок связывания винкулина.
Тензин - белок, кепирующий актиновые филаменты с их плюс концов.
Паксиллин- молекулярная масса 68 кД.
α-актинин- в клетках хороидного плексуса(однослойный секреторный эпителий, выстилающий желудочки мозга) альфа-актинин локализован главным образом в апикальной мембране и связываются предпочтительно с Na/К-АТФазой , но не обнаруживаются вблизи базолатеральной мембраны, где находится белок полосы 3. α-актинин связывает актиновые филаменты в пучки и сети in vitro и локализуется в ламеллиподии. Различные изоформы частично разделяются пространственно между разными областями локализации актина, причем актинин 1 и актинин 4 локализуется в раффлах. Клетки Dictyostelium , лишенные α-актина, не обнаруживают дефектов двигательной активности за исключением клеток с отсутствием гомолога филамина, что заставляет предположить структурную совместимость этих кросслинкеров в ламеллиподии. В синтетических кометных хвостах актина, недостаток α-актинина проявляется в образовании менее компактной структуры хвоста, подтверждая кросслинкующую функцию этого белка.
3.4 Десмосомы
Десмосомы - кнопковидные межклеточные контакты, скрепляющие клетки друг с другом. С цитоплазматической стороны к ним прикрепляются промежуточные филаменты, которые формируют структурный остов цитоплазмы, выдерживающий большие силы натяжения. Таким образом, через десмосомы промежуточные филаменты соседних клеток опосредованно объединяются в непрерывную сеть по всей ткани. Таким образом десмосомы действуют в качестве заклепок, распределяющих силы натяжения или разрыва по эпителиальному слою.
Тип промежуточных филаментов, прикрепленных к десмосомам, зависит от типа клеток: в большинстве эпителиальных клеток к десмосомам прикреплены кератиновые промежуточные филаменты; в клетках сердечной мышцы - десминовые промежуточные филаменты . Сеть промежуточных филаментов в десмосоме ассоциирована с плотной бляшкой на цитоплазматической поверхности контактной плазматической мембраны . Десмосомы - наиболее распространенные адгезионные элементы в эпителиях и сердечной мышце . В отличии от кадхериновой адгезии, десмосомы связаны с промежуточными филаментами (в эпителии - с цитокератинами , а в сердце - с десминовыми филаментами). Вместе десмосомы и промежуточные филаменты формируют в тканях непрерывную сеть. Адгезионные рецепторы в десмосомах - члены суперсемейства кадгеринов, десмоколлины и десмоглеины (это трансмембранный гликопротеин с молекулярной массой около 150 кД., цитоплазматическая негликозилированная часть десмоглеина входит в состав бляшки десмосомы, а наружная гликозилированная часть достигает центрального диска и врастает в него), среди которых встречаются тканеспецифически экспрессирующиеся изоформы.
Десмоглеины и десмоколлины прикреплены к промежуточным филаментам при помощи нескольких цитоплазматических белков, таких как десмоплакины и плакоглобин. Десмоплакины имеют определенную гомологию с белками промежуточных филаментов и, по- видимому, связаны непосредственно с ними.
Плакоглобин (белок с молекулярной массой 83 кД, обнаруживающийся в адгезионных межклеточных контактах) связывается с цитоплазматическим участком некоторых десмоглеинов и десмоколлинов (белок с молекулярной массой 240 кД, вероятно, непосредственно участвующий в заякоривании промежуточных филаментов) и возможно является центральным пунктом в формировании десмосомы и прикреплении цитокератиновых филаментов. Плакоглобин, имеющий гомологию с β-катенином, также участвует в трансдукции сигналов.
Десмосомы и цитокератины обеспечивают механическую прочность, необходимую для поддержания целостности эпидермиса. Система десмосом и промежуточных филаментов в других тканях, по-видимому, имеет сходную роль.
4. Гемидесмосома
Гемидесмосому называют также полудесмосомой.
В отличие от десмосом, соединяющих мембраны соседних эпителиальных клеток, гемидесмосомы присоединяют базальную поверхность эпителиальных клеток к подлежащей базальной мембране, тем самым, однако, также, как и десмосомы , функционируя в качестве заклепок, распределяющих силы натяжения или разрыва, но уже на подлежащую эпителий соединительную ткань . В то время как промежуточные филаменты, ассоциированные с десмосомами, латерально прикрепляются к десмосомным бляшкам, многие из промежуточных филаментов, ассоциированных с гемидесмосомами, своими концами погружены в бляшку. Внутриклеточные прикрепляющие белки гемидесмосом отличны от подобных белков десмосом. Трансмембранные линкерные белки гемидесмосом принадлежат к интегриновому семейству рецепторов внеклеточного матрикса.
Как и десмосомы, гемидесмосомы прикрепляют промежуточные филаменты, однако основным адгезионным рецептором в данном случае является альфа-6 бета-4-интегрин, прикрепляющий ламинин (на ранних этапах развития базальная мембрана состоит в основном из сети ламинина и не содержит (или содержит мало) коллагена типа IV); ламинин, адгезивный гликопротеин - большой (молекулярная масса 850000) гибкий комплекс из длинных полипептидных цепей, ассоциированных в форме асимметричного креста и удерживаемых вместе при помощи дисульфидных связей. Содержит несколько функциональных доменов: связывающиеся с коллагеном типа IV, с гепаран сульфатом, с энтактином, c рецепторами ламинина на клеточной поверхности к базальной ламине. Остальные белки, составляющие гемидесмосому, также уникальны, хотя и отчасти гомологичны десмосомальным белкам.
5. Заболевание аутоиммунное, поражающее десмосомы
Некоторые формы кожного заболевания pemphigus характеризуются образованием аутоиммунных антител против одного из десмосомных кадхериновых белков. Такие антитела разрушают десмосомы между кератиноцитами - клетками эпидермиса. Кератиноциты называются так потому, что характерной чертой их дифференцированного состояния является синтез кератинас. От слоя к слою эти клетки изменяют свой внешний вид. Самый глубокий из внутренних слоев образован базальными клетками. В основном именно эти клетки делятся путем митоза. Над базальными клетками находится несколько слоев более крупных шиповатых клеток. ) , что приводит к образованию волдырей в результате поступления жидкостей тела в разрыхленный эпителий. Разрушение десмосом только в эпидермисе предполагает, что эти десмосомы биохимически отличны от десмосом в других тканях.
6. Проводящие межклеточные контакты
Коммуникационные соединения в клетках животных представлены так называемыми щелевыми контактами и синапсами.
Щелевое соединение, или нексус, представляет собой область протяженностью 0,5—3 мкм, где плазмолеммы разделены промежутком в 2—3 нм. Со стороны цитоплазмы никаких специальных примембранных структур в данной области не обнаруживается, но в структуре плазмолемм соседних клеток друг против друга располагаются специальные белковые комплексы (коннексоны), которые образуют как бы каналы из одной клетки в другую. Этот тип соединения встречается во всех группах тканей.
Функциональная роль щелевого соединения заключается в переносе ионов и мелких молекул (молекулярная масса 2 -10) от клетки к клетке. Так, в сердечной мышце возбуждение, в основе которого лежит процесс изменения ионной проницаемости, передастся от клетки к клетке через нексус.
Синоптические соединения, или синапсы. Этот тин соединений характерен для нервной ткани и встречается в специализированных участках контакта как между двумя нейронами, так и между нейроном и каким-либо иным элементом, входящим в состав рецептора или эффектора (например, нервно-мышечные, нервно-эпителиальные синапсы).
6.1 Синапсы
Синапсы — участки контактов двух клеток, специализированных для односторонней передачи возбуждения или торможения от одного элемента к другому.
Обеспечивают передачу потенциала действия (нервного импульса) с нервной клетки на другую нервную или иную клетку.
Синаптические контакты или синапсы - специфические контакты между нервными клетками (межнейронные синапсы) или между нервными и другими клетками (нервно-мышечные синапсы и другие). Функциональная роль синаптических контактов заключается в передаче возбуждения или торможения с одной нервной клетки на другую или с нервной клетки на иннервируемую клетку.
6.2 Нексусы
Нексусы или щелевидные контакты.
Контакт образуется на небольшом по площади участке.
В месте контакта в цитомембрану встроены трансмембранные белки коннексины, которые соединяются между собой и образуют водный канал в толще мембраны — конексон.
Коннексоны контактирующих клеток соединяются (или сопоставляются), в результате чего между соседними клетками образуется канал, с помощью которого из одной клетки в другую (в обоих направлениях) свободно проходит вода, малые молекулы и ионы, а также электрический ток.
Щелевидные контакты или нексусы ограниченные участки контакта соседних цитолемм, диаметром 0,5-3,0 мкм, в которых билипидные мембраны сближены на расстояние 2-3 нм, а обе мембраны пронизаны в поперечном направлении белковыми молекулами коннексонами, содержащими гидрофильные каналы. Через эти каналы осуществляется обмен ионами и микромолекулами соседних клеток, чем и обеспечивается их функциональная связь (например, распространение биопотенциалов между кардиомиоцитами, их содружественное сокращение в миокарде).
7. Рецепторы
Биохимические рецепторы мембран клетки - разновидность биохимических рецепторов, структуры, как правило, молекулы белка, расположенные на наружной поверхности мембраны.
Биохимический рецептор имеет высокую степень сродства к определенным химическим соединениям, обладает свойством стереоспецифически связывать их. Эти химические соединения, служат средством передачи информации через мембрану. Содержание информации «закодированной» в химическом соединении может быть различным: информация о цели системы, управляющие сигналы, информация о среде, информация о состоянии объекта управления, информация о результатах управления. В числе таких химических соединений переносчиков информации могут быть: нейромедиаторы, гормоны, иммуноглобулины и ряд других веществ.
Химический рецептор может считаться первичным посредником передачи информации от управляющего звена или регулятора к объекту управления. Этот рецептор может быть функционально связан с системой мембранных белков - вторичных посредников передачи информации через мембрану. Примером такой системы является аденилатциклатная группа. В ней внешний химический сигнал, например гормон, образует комплекс с рецептором мембраны, расположенном на наружной её поверхности. Образовавшийся комплекс гормон-рецептор активирует систему белков, управляющую активностью фермента аденилатциклазы, расположенной на внутренней поверхности мембраны. Активированная аденилциклаза катализирует образование циклического аденозинмонофосфата из аденозинтрифосфата. Циклический аденозинмонофосфат влияет на метаболические реакции.
Таким образом, рецепторы живых мембран являются структурами, принимающими участие в механизмах управления функциями клеток посредством медиаторов, гормонов и других активных веществ.
Вывод
Межклеточные соединения – соединения между клетками, образованные при помощи белков.
Межклеточные связи сводятся не только к электрическим взаимодействиям. Взаимосвязь между клетками является более сложной. Клетки органов и тканей вырабатывают ряд химических веществ, действующих на другие клетки и вызывающих включение/выключение (усиление/ослабление) функции механического сцепления между клетками, изменение интенсивности обмена веществ и процесса синтеза клеткой белков.
В тех тканях, в которых клетки или их отростки плотно прилежат друг к другу (эпителий, мышечная ткань и пр.) между мембранами контактирующих клеток формируются связи – межклеточные контакты. В большинстве случаев межклеточные соединения разрушаются при удалении из среды ионов Ca2+ .
Межклеточные соединения возникают в местах соприкосновения клеток в тканях и служат для межклеточного транспорта веществ и передачи сигналов (межклеточное взаимодействие), а также для механического скрепления клеток друг с другом.
Межклеточные соединения выполняют ряд важных функций: обмен ионами и микромолекулами соседних клеток; передача возбуждения или торможения с одной нервной клетки на другую или с нервной клетки на иннервируемую клетку; обеспечивают механическую прочность, необходимую для поддержания целостности эпидермиса; связь друг с другом клеток в различных эпителиях, в сердечных и гладких мышцах.
Межклеточные соединения сильно зависят от присутствия Са2+ . При недостатке ионов в среде контакты разрушаются.
Приложение 1
Список литературы
1.Геннис, Биомембраны. Молекулярная структура и функции. М., 1997.
2.Евгеньва Т.П., Межклеточные взаимодействия и их роль в эволюции. М., 1976.
3.Межклеточные взаимодействия, перевод с англ. М.,1980.
4.Ленинджер А., 1985.
5.Марри, Греннер, 1993.
6.Ткачук В.А., Бочкова В.Н., Филиппова М.П., Стамбольский Д.В., Т-кадгедрин – новый белок межклеточного взаимодействия: структура, локализация, механизмы сигнализации. М., Российский кардиологический научный центр.