| Скачать .docx | Скачать .pdf |
Курсовая работа: рДНК-биотехнология. Способы биотрансформации клеток
СОДЕРЖАНИЕ
Введение
1 Основные группы ферментов генетической инженерии
1.1 Рестриктазы
1.1.1 Механизм действия рестриктаз
1.1.2 Построение рестрикционных карт
1.3 Лигазы
2 Введение нового гена в клетку
2.1 Регуляция экспрессии гена у прокариот
2.2 Способы прямого введения гена в клетку
2.3 Введение генов в клетки млекопитающих
2.4 Генетическая трансформация соматических клеток млекопитающих
2.5 Генотерапия
2.6 Получение трансгенных животных
Заключение
Список литературы
Введение
Генетическая инженерия - конструирование in vitro функционально активных генетических структур (рекомбинантных ДНК), или иначе - создание искусственных генетических программ (Баев А. А.). По Э. С. Пирузян генетическая инженерия - система экспериментальных приемов, позволяющих конструировать лабораторным путем (в пробирке) искусственные генетические структуры в виде так называемых рекомбинантных или гибридных молекул ДНК.
Речь идет о направленном, по заранее заданной программе конструировании молекулярных генетических систем вне организма с последующим введением их в живой организм. При этом рекомбинантные ДНК становятся составной частью генетического аппарата рецепиентного организма и сообщают ему новые уникальные генетические, биохимические, а затем и физиологические свойства.
Цель прикладной генетической инженерии заключается в конструировании таких рекомбинантных молекул ДНК, которые при внедрении в генетический аппарат придавали бы организму свойства, полезные для человека.
Технология рекомбинантных ДНК использует следующие методы:
- специфическое расщепление ДНК рестрицирующими нуклеазами, ускоряющее выделение и манипуляции с отдельными генами;
- быстрое секвенирование всех нуклеотидов очищенном фрагменте ДНК, что позволяет определить границы гена и аминокислотную последовательность, кодируемую им;
- конструирование рекомбинантной ДНК;
- гибридизация нуклеиновых кислот, позволяющая выявлять специфические последовательности РНК или ДНК с большей точностью и чувствительностью, основанную на их способности связывать комплементарные последовательности нуклеиновых кислот;
- клонирование ДНК: амплификация in vitro с помощью цепной полимеразной реакции или введение фрагмента ДНК в бактериальную клетку, которая после такой трансформации воспроизводит этот фрагмент в миллионах копий;
- введение рекомбинантной ДНК в клетки или организмы.
История генетической инженерии
Генная инженерия появилась благодаря работам многих исследователей в разных отраслях биохимии и молекулярной генетики. На протяжении многих лет главным классом макромолекул считали белки. Существовало даже предположение, что гены имеют белковую природу. Лишь в 1944 году Эйвери, Мак Леод и Мак Карти показали, что носителем наследственной информации является ДНК. С этого времени начинается интенсивное изучение нуклеиновых кислот. Спустя десятилетие, в 1953 году Дж. Уотсон и Ф. Крик создали двуспиральную модель ДНК. Именно этот год принято считать годом рождения молекулярной биологии.
На рубеже 50 - 60-х годов были выяснены свойства генетического кода, а к концу 60-х годов его универсальность была подтверждена экспериментально. Шло интенсивное развитие молекулярной генетики, объектами которой стали E. coli, ее вирусы и плазмиды. Были разработаны методы выделения высокоочищенных препаратов неповрежденных молекул ДНК, плазмид и вирусов. ДНК вирусов и плазмид вводили в клетки в биологически активной форме, обеспечивая ее репликацию и экспрессию соответствующих генов. В 70-х годах был открыт ряд ферментов, катализирующих реакции превращения ДНК. Особая роль в развитии методов генной инженерии принадлежит рестриктазам и ДНК-лигазам.
Историю развития генетической инженерии можно условно разделить на три этапа. Первый этап связан с доказательством принципиальной возможности получения рекомбинантных молекул ДНК in vitro. Эти работы касаются получения гибридов между различными плазмидами. Была доказана возможность создания рекомбинантных молекул с использованием исходных молекул ДНК из различных видов и штаммов бактерий, их жизнеспособность, стабильность и функционирование.
Второй этап связан с началом работ по получению рекомбинантных молекул ДНК между хромосомными генами прокариот и различными плазмидами, доказательством их стабильности и жизнеспособности.
Третий этап - начало работ по включению в векторные молекулы ДНК (ДНК, используемые для переноса генов и способные встраиваться в генетический аппарат клетки-рецепиента) генов эукариот, главным образом, животных.
Формально датой рождения генетической инженерии следует считать 1972 год, когда в Стенфордском университете П. Берг, С. Коэн, Х. Бойер с сотрудниками создали первую рекомбинантную ДНК, содержавшую фрагменты ДНК вируса SV40, бактериофага и E. coli.
1 Основные группы ферментов генетической инженерии
Генетическая инженерия - потомок молекулярной генетики, но своим рождением обязана успехам генетической энзимологии и химии нуклеиновых кислот, так как инструментами молекулярного манипулирования являются ферменты. Если с клетками и клеточными органеллами мы подчас можем работать микроманипуляторами, то никакие, даже самые мелкие микрохирургические инструменты не помогут при работе с макромолекулами ДНК и РНК. Что же делать? В роли "скальпеля", "ножниц" и "ниток для сшивания" выступают ферменты.
Только они могут найти определенные последовательности нуклеотидов, "разрезать" там молекулу или, наоборот, "заштопать" дырку в цепи ДНК. Эти ферменты издавна работают в клетке, выполняя работы по репликации (удвоению) ДНК при делении клетки, репарации повреждений (восстановлению целостности молекулы), в процессах считывания и переноса генетической информации из клетки в клетку или в пределах клетки. Задача генного инженера - подобрать фермент, который выполнил бы поставленные задачи, то есть смог бы работать с определенным участком нуклеиновой кислоты.
Следует отметить, что ферменты, применяемые в генной инженерии, лишены видовой специфичности, поэтому экспериментатор может сочетать в единое целое фрагменты ДНК любого происхождения в избранной им последовательности. Это позволяет генной инженерии преодолевать установленные природой видовые барьеры и осуществлять межвидовое скрещивание.
Ферменты, применяемые при конструировании рекомбинантных ДНК, можно разделить на несколько групп:
- ферменты, с помощью которых получают фрагменты ДНК (рестриктазы);
- ферменты, синтезирующие ДНК на матрице ДНК (полимеразы) или РНК (обратные транскриптазы);
- ферменты, соединяющие фрагменты ДНК (лигазы);
- ферменты, позволяющие осуществить изменение структуры концов фрагментов ДНК.
1.1 Рестриктазы
Общепринято термины "рестриктаза", "эндонуклеаза рестрикции" и "сайт специфическая эндодезоксирибонуклеаза" считать синонимами.
Все рестрикционные эндонуклеазы бактерий узнают специфические, довольно короткие последовательности ДНК и связываются с ними. Этот процесс сопровождается разрезанием молекулы ДНК либо в самом сайте узнавания, либо в каком-то другом, что определяется типом фермента. Наряду с рестрикционной активностью бактериальный штамм обладает способностью метилировать ДНК; для этого процесса характерна такая же специфичность в отношении последовательностей ДНК, как и для рестрикции. Метилаза добавляет метильные группы к адениновым или цитозиновым остаткам в том же сайте, в котором связывается рестрикционный фермент. В результате метилирования сайт становится устойчивым к рестрикции. Следовательно, метилирование защищает ДНК от разрезания.
Различают 3 основных класса рестриктаз: 1, 2 и 3.
Все рестриктазы узнают на двуспиральной ДНК строго определенные последовательности, но рестриктазы 1-го класса осуществляют разрывы в произвольных точках молекулы ДНК, а рестриктазы 2-го и 3-го классов узнают и расщепляют ДНК в строго определенных точках внутри сайтов узнавания или на фиксированном от них расстоянии.
Ферменты типов 1 и 3 имеют сложную субъединичную структуру и обладают двумя типами активностей - модифицирующей (метилирующей) и АТФ-зависимой эндонуклеазной.
Ферменты второго класса состоят из 2 отдельных белков: рестрицирующей эндонуклеазы и модифицирующей метилазы, поэтому в генной инженерии используются исключительно ферменты 2-го класса. Они нуждаются в ионах магния в качестве кофакторов.
В настоящее время выделено более 500 рестриктаз класса 2, однако среди ферментов, выделенных из различных микроорганизмов, встречаются такие, которые узнают на ДНК одни и те же последовательности. Такие пары или группы называют изошизомерами. Различают истинную изошизомерию, когда ферменты узнают одну и ту же последовательность нуклеотидов и разрывают ДНК в одних и тех же точках, и ложную, когда ферменты, узнавая один и тот же сайт на ДНК, производят разрывы в разных точках в пределах того же сайта.
Большинство рестриктаз класса 2 узнают последовательности, содержащие от 4 до 6 нуклеотидных пар, поэтому рестриктазы делят на мелко- и крупнощепящие. Мелкощепящие рестриктазы узнают тетрануклеотид и вносят в молекулы гораздо больше разрывов, чем крупнощепящие, узнающие последовательность из шести нуклеотидных пар. Это связано с тем, что вероятность встречаемости определенной последовательности из четырех нуклеотидов гораздо выше, чем последовательности из шести нуклеотидов. Например, в ДНК бактериофага Т7, состоящей из 40000 пар оснований, отсутствует последовательность, узнаваемая рестриктазой R1 из E. coli.
К мелкощепящим относятся рестриктазы Hpa II и Alu (из Arthrobacter luteus), к крупнощепящим - Eco R I (из Escherichia coli) и Hind III. Если предположить, что участки узнавания рестриктаз распределены вдоль цепи ДНК случайно, то мишень для ферментов, узнающих последовательность (сайт) из четырех нуклеотидов, должна встречаться в среднем 1 раз через каждые 256 пар оснований, а для ферментов, узнающих шесть нуклеотидов, - через 4096 пар оснований. Если сайт рестрикции окажется внутри гена, то обработка ДНК-рестриктазой приведет к его инактивации. Вероятность такого события очень велика при обработке мелкощепящими рестриктазами и незначительна при применении крупнощепящих эндонуклеаз. Поэтому с целью получения неповрежденного гена расщепление проводят поочередно несколькими крупнощепящими рестриктазами, либо применяют прием "недорестрикции", т.е. рестрикцию проводят в таких условиях, когда происходит расщепление лишь в одном сайте.
1.1.1 Механизм действия рестриктаз
В качестве мишеней (мест узнавания) часто выступают палиндромы из 4-6 пар оснований - сайты рестрикции. Точки узнавания рестриктазами симметричны относительно поворота на 180оС, то есть последовательность нуклеотидов слева направо в одной нити такая же, как справа налево в другой. Симметрия подразумевает, что те из них, которые должны быть метилированы, встречаются на обеих цепях ДНК. В результате сайт-мишень может быть полностью метилирован (обе цепи модифицированы), полуметилирован (только одна цепь метилирована) или не метилирован.
Полностью метилированный сайт не подвержен ни рестрикции, ни модификации. Полуметилированный сайт не узнается ферментом рестрикции, но может быть превращен с помощью метилазы в полностью метилированный. У бактерий метилирование, как правило, связано с сохранением имеющегося состояния модификации. Репликация полностью метилированной ДНК ведет к образованию полуметилированной ДНК. Вероятно узнавание полуметилированных сайтов представляет собой обычный этап функционирования метилазы in vivo.
Неметилированный сайт-мишень пред¬ставляет собой субстрат либо для рестрикции, либо для модификации in vitro. В клетке немодифицированная ДНК с большей вероятностью рестрицируется. Реакция разрезания осуществляется в две ступени. Сначала разрезается одна цепь ДНК, а затем рядом разрезается другая. В областях, прилегающих с каждой стооны к сайту разрезания, может иметь место экзонуклеотическая деградация. Происходит эффективный гидролиз АТФ, роль которого еще не выяснена.
Каким образом фермент узнает один сайт, а разрезает другой, достаточно удаленный? Важно отметить, что белок никогда не отделяется от молекулы ДНК, с которой он первоначально связался. Если фермент инкубировать со смесью модифицированной и немодифицированной ДНК, он предпочтительно разрезает немодифицированную ДНК. Следовательно, узнавая сайт связывания, белок не отделяется от неметилированной ДНК для того, чтобы найти сайт разрезания.
Существуют две альтернативные модели, объясняющие взаимосвязь между сайтами узнавания и разрезания: в соответствии с одной из них движется фермент, согласно другой модели, перемещается ДНК. Если движется фермент, то его перемещение вдоль ДНК будет продолжаться до тех пор, пока он не сделает выбор сайта разрезания. Если же движется ДНК, то фермент остается прикрепленным в сайте узнавания, а ДНК протаскивается через второй сайт связывания на ферменте, и это продолжается до тех пор, пока фермент не достигает области разрезания (пока не охарактеризованной). Получены электронно-микроскопические данные, свидетельствующие, что фермент вызывает образование петли в ДНК и остается, по-видимому, связанным с сайтом узнавания после разрезания; эти данные подтверждают вторую модель.
1.1.2 Построение рестрикционных карт
Ферменты рестрикции стали эффективным инструментом исследования. Они позволяют превращать молекулы ДНК очень большого размера в набор фрагментов длиной от нескольких сотен до нескольких тысяч оснований. С помощью метода электрофореза в агарозном геле (см. раздел 1) фрагменты ДНК, различающиеся по размеру, можно легко разделить, а затем исследовать каждый фрагмент отдельно.
Короткие фрагменты мигрируют намного быстрее, чем длинные. При сравнительно высокой концентрации агарозы большие фрагменты вообще не могут проникнуть в гель. В процессе миграции рестрикционные фрагменты не деградируют, их можно элюировать (вымывать) в виде биологически активных двуцепочечных молекул. При окрашивании гелей красителями, связывающимися с ДНК, выявляется набор полос, каждая из которых отвечает рестрикционному фрагменту, молекулярную массу которого можно определить, проведя калибровку с помощью ДНК с известными молекулярными массами.
Сравнение размеров фрагментов ДНК, полученных после обработки определенного участка генома набором рестрицирующих нуклеаз позволяет построить рестрикционную карту, на которой указано положение каждого сайта рестрикции относительно других участков.
Молекулу ДНК длиной 5000 пар нуклеотидов (п. н.). обрабатывают отдельно рестриктазами А и В. Фрагменты разделяют электрофорезом. Фермент А разрезал ДНК на 4 фрагмента размером 2100, 1400, 1000 и 500 п. н. Обработка рестриктазой В дала 3 фрагмента: 2500, 1300 и 1200 п. н. (рис. 37). Для определения расположения сайтов рестрикции этих ферментов на следующем этапе применяют процедуру двойного расщепления – обрабатывают ДНК двумя эндонуклеазами. Обработка изучаемого фрагмента одновременно двумя рестриктазами дала 6 фрагментов: 1900, 1000, 800, 600, 500, 200 п. н.
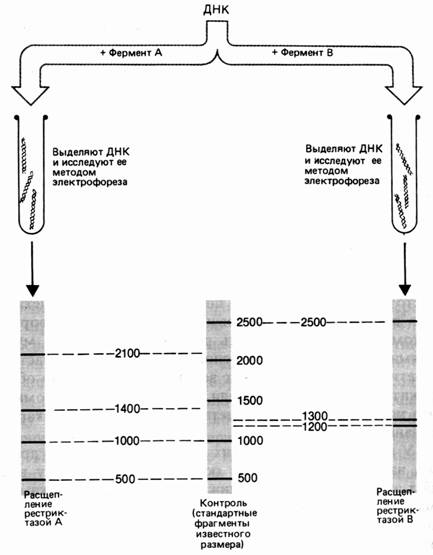
Рис. 1. Результаты электрофореза после обработки фрагмента ДНК разными рестриктазами
Наиболее полный вариант – элюировать каждый фрагмент, образующийся в результате расщепления одной рестриктазой, а затем обработать его второй. Смесь фрагментов, полученных после такой обработки, также анализируют с помощью электрофореза. В нашем примере были получены следующие результаты:
Обработка каждого из 4-х А-фрагментов рестриктазой В
2100 - 1900 и 200,
1400 - 800 и 600,
1000 - 1000 (изменений нет)
500 - 500 (изменений нет)
Обработка каждого из 3-х В-фрагментов рестриктазой А
2500 - 1900 и 600
1300 - 800 и 500
1200 - 1000 и 200
Анализ полученных результатов показывает, что каждый из ферментов, полученный при расщеплении А-фрагментов рестриктазой В можно обнаружить в образцах, полученных при расщеплении В-фрагментов рестриктазой А. Ключом к рестрикционному картированию являются перекрывающиеся фрагменты. Такими в рассматриваемом примере являются В-фрагмент 2100 и А-фрагмент 2500. При обработке другой рестриктазой они дают фрагмент 1900.
Из данных о расщеплении этих фрагментов мы предполагаем, что с одной стороны на расстоянии 200 п. н. от фрагмента 1900 находится следующий А-сайт, а с другого конца, на расстоянии 600 п. н. – следующий В-сайт (рис. 38). При обработке двумя эндонуклеазами фрагмент 200 п. н. образуется 1 раз, при обработке рестриктазой А из В-фрагмента 1200, т. е. фрагмент 1200 лежит слева. Остается определить, как продолжается карта вправо. Очевидно, это А-фрагмент 1400, так как он рассечен рестриктазой В на фрагменты 600 и 800. Вправо от фрагмента 2500 следует отложить, очевидно, фрагмент 1300. Тогда логично наличие А-фрагмента 500 и деления В-фрагмента 1300 рестриктазой А на 800 и 500.
При построении рестрикционных карт обычно используют несколько рестриктаз, поэтому приходится анализировать сложные соотношения между фрагментами, полученными при действии разных ферментов. Для упрощения процедуры картирования можно применять неполное расщепление. В определенных условиях рестриктаза узнает и расщепляет не все сайты в молекуле ДНК. Например, при частичном расщеплении ДНК ферментом А могут образовываться фрагменты 3100 п. н., 1400 п. н. и 500 п. н. Сопоставив их с данными полного расщепления (2100, 1400, 1000 и 500), можно сразу поставить рядом 2100 и 1000 (фрагмент 3100). А получив фрагмент 3500 – расположить рядом 2100 п. н. и 1400 п. н.

Рис. 2. Анализ фрагментов рестрикции и карта фрагмента ДНК
Другой прием – введение радиоактивной концевой метки. Концевые фрагменты определяются в этом случае по включению метки. Можно также сопоставить фрагменты путем гибридизации нуклеиновых кислот. Перекрывающиеся фрагменты (в данном случае 2100 и 2500) будут гибридизоваться.
Первая карта была получена для вируса SV40 (обезьяний вирус, вызывающий злокачественную трансформацию), содержащего 5423 пары оснований. Использовали рестриктазу Hind-II, расщепляющую кольцевую ДНК вируса на 11 фрагментов. Порядок их расположения в ДНК был установлен путем исследования наборов фрагментов, образующихся по мере того, как расщепление доходит до конца. Первый разрыв превращал кольцевую молекулу в линейную, которая затем расщеплялась на все меньшие и меньшие фрагменты. Исследовали вначале наборы перекрывающихся фрагментов, а затем продукты полного расщепления. Таким образом была получена рестрикционная карта кольцевой вирусной ДНК, на которую были нанесены сайты расщепления рестриктазой. Повторив подобные эксперименты с другой рестриктазой можно получить более подробную карту, где отмечено много сайтов рестрикции.
Располагая такой информацией, можно идентифицировать на ДНК биологически важные участки. Поскольку рестрикционная карта отражает расположение определенной последовательности нуклеотидов в данном участке, сравнение таких карт для двух или более родственных генов позволяет оценить гомологию между ними. Анализируя рестрикционные карты, можно сравнивать определенные участки ДНК разных видов животных без определения их нуклеотидной последовательности. Таким образом, например, было установлено, что хромосомные участки, кодирующие цепи гемоглобина у человека, орангутанга и шимпанзе сохранились в практически неизменном виде в течение последних 5 - 10 млн. лет (с тех пор как виды дивергировали).
Метод рестрикционного картирования позволяет увидеть крупные генетические изменения, такие как делеции или инсерции. При этом происходит уменьшение или увеличение рестрикционных фрагментов, а также исчезновение или возникновение сайтов рестрикции.
Один из приемов картирования – фингерпринт («метод отпечатков пальцев» или DNA-fingerprint). Он подразумевает использование неупорядоченных и неполных наборов фрагментов, которые являются характеристикой генома, хотя описывает его не полностью.
1.2 Обратная транскриптаза
Обратная транскриптаза используется для транскрипции м-РНК в комплементарную цепь ДНК. При изучении ретровирусов, геном которых представлен молекулами одноцепочечной РНК, было обнаружено, что в процессе внутриклеточного развития ретровирус проходит стадию интеграции своего генома в виде двухцепочечной ДНК в хромосомы клетки-хозяина. В 1964 г. Темин выдвинул гипотезу о существовании вирусспецифичного фермента, способного синтезировать на РНК-матрице комплементарную ДНК. Усилия, направленные на выделение такого фермента, увенчались успехом, и в 1970 г. Темин с Мизутани, а также независимо от них Балтимор открыли искомый фермент в препарате внеклеточных вирионов вируса саркомы Рауса. Данная РНК-зависимая ДНК-полимераза получила название обратная транскриптаза, или ревертаза.
Наиболее детально изучена ревертаза ретровирусов птиц. Каждый вирион содержит около 50 молекул этого фермента. Обратная транскриптаза состоит из двух субъединиц — a (65 кДа) и b (95 кДа), присутствующих в эквимолярном количестве. Обратная транскриптаза обладает, по крайней мере, тремя ферментативными активностями:
1) ДНК-полимеразной, использующей в качестве матрицы как РНК, так и ДНК;
2) активностью РНКазы Н, гидролизующей РНК в составе гибрида РНК—ДНК, но не одно- или двухцепочечную РНК;
3) ДНК-эндонуклеазной активностью.
Первые две активности необходимы для синтеза вирусной ДНК, а эндонуклеаза, по-видимому, важна для интеграции вирусной ДНК в геном клетки-хозяина. Очищенная обратная транскриптаза синтезирует ДНК как на РНК-, так и на ДНК-матрицах. Чтобы начать синтез, ревертазе, как и другим полимеразам, необходим короткий двухцепочечный участок (праймер). Праймером может служить одноцепочечный сегмент как РНК, так и ДНК, которые в процессе реакции оказываются ковалентно связанными с новосинтезированной цепью ДНК.

Рис. 3. Схема синтеза двухцепочечных ДНК-копий молекул РНК
Обратную транскриптазу преимущественно используют для транскрипции матричной РНК в комплементарную ДНК (кДНК). Реакцию обратной транскрипции проводят в специально подобранных условиях с использованием сильных ингибиторов РНКазной активности. При этом удается получать полноразмерные ДНК-копии целевых молекул РНК. В качестве праймера при обратной транскрипции поли (А)-содержащих мРНК используют олигo (dT), а для молекул РНК, не имеющих З'-поли (А) концов, — химически синтезированные олигонуклеотиды, комплементарные З'-концу изучаемой РНК. После синтеза на мРНК комплементарной цепи ДНК и разрушения РНК (обычно применяют обработку щелочью) осуществляют синтез второй цепи ДНК. При этом используют способность ревертазы образовывать на 3'-концах одноцепочечных кДНК самокомплементарные шпильки, которые могут выполнять функции праймера.
Матрицей служит первая цепь кДНК. Данная реакция может катализироваться как ревертазой, так и ДНК-полимеразой I E. coli. Показано, что сочетание этих двух ферментов позволяет повысить выход полноценных двухцепочечных молекул кДНК. По окончании синтеза первая и вторая цепи кДНК остаются ковалентно связанными петлей шпильки, служившей праймером при синтезе второй цепи. Эту петлю расщепляют эндонуклеазой S1, специфически разрушающей одноцепочечные участки нуклеиновых кислот. Образующиеся при этом концы не всегда оказываются тупыми, и для повышения эффективности последующего клонирования их репарируют до тупых с помощью фрагмента Кленова ДНК-полимеразы I E. coli. Полученную двухцепочечную кДНК можно затем встраивать в клонирующие векторы, размножать в составе гибридных молекул ДНК и использовать для дальнейших исследований.
1.3 Лигазы
В 1961 г. Мезельсон и Вейгл на примере фага l показали, что рекомбинация включает разрыв и последующее воссоединение молекул ДНК. Это положило начало поискам фермента, участвующего в сшивании фрагментов ДНК. В 1967 году такой фермент был найден и получил название ДНК-лигаза. Он катализирует синтез фосфодиэфирной связи в 2-х цепочечной молекуле нуклеиновой кислоты.
Иными словами, ДНК-лигазы сшивают рядом расположенные нуклеотиды, образуя связь между остатками сахаров. ДНК-лигазы абсолютно необходимы в процессах репарации ДНК, в процессах репликации - при удвоении цепи ДНК.
Существует 2 типа ДНК-лигаз, отличающихся по потребностям в кофакторах и способу действия. ДНК-лигаза E. coli в качестве кофактора использует дифосфопиридиннуклеотид, а лигаза фага Т4 - АТФ в присутствии Mg2+. Лигаза фага Т4 более универсальна, так как помимо лигирования липких концов способна катализировать реакцию воссоединения двухцепочечных фрагментов ДНК с тупыми концами. Она используется чаще.
2 Введение нового гена в клетку
Ввести рекомбинантный ген в клетку можно 2 способами: используя вектора или путем прямого введения.
Требования к векторной ДНК, ее состав
Вектор - молекула ДНК или РНК, состоящая из двух компонентов: векторной части (носителя) и клонируемого чужеродного гена. Задача вектора – донести выбранную ДНК в клетку-рецепиент, встроить ее в геном, позволить идентификацию трансформированных клеток, обеспечить стабильную экспрессию введенного гена.
Таким образом, вектор должен быть небольшим, способным поддерживаться в клетке-хозяине (реплицироваться), многократно копироваться (ампфлицироваться), экспрессировать соответствующий ген (содержать соответствующие регуляторные последовательности), должен иметь маркерный ген, позволяющий различать гибридные клетки для эффективной селекции их; должен быть способен передаваться в клетку соответствующего организма.
Регуляторные последовательности, отвечающие за стабильную экспрессию гена, будут рассмотрены позднее.
Можно выделить 2 группы маркерных генов, позволяющие отличить трансформированные клетки:
1. Селективные гены, отвечающие за устойчивость к антибиотикам (канамицину, тетрациклину, неомицину и др.), гербицидам (у растений). Это могут быть гены ауксотрофности по какому-либо субстрату и т.д. Основной принцип работы такого маркера – способность трансформированных клеток расти на селективной питательной среде, с добавкой определенных веществ, ингибирующих рост и деление нетрансформированных, нормальных клеток.
2. Репортерные гены, кодирующие нейтральные для клеток белки, наличие которых в тканях может быть легко тестировано.
Чаще всего в качестве репортерных используются гены β-глюкуронидазы (GUS), зеленого флюоресцентного белка (GFP), люциферазы (LUC), хлорамфениколацетилтрансферазы (CAT). К настоящему времени из этого арсенала наиболее часто используют гены GUS и GFP и, в меньшей степени, LUC и CAT. Используемый в настоящее время как репортерный ген GUS является модифицированным геном из Escherichia coli, кодирующим β-глюкуронидазу с молекулярной массой 68 кД. GUS активен в широком диапазоне условий среды с оптимумом при рН 5-8 и 37°С. Он может гидролизовать обширный спектр природных и синтетических глюкуронидов, что позволяет подбирать соответствующие субстраты для спектрофотометрического или флюориметрического определения активности фермента, а также для гистохимического окрашивания тканей in situ (например, в синий цвет). Фермент достаточно стабилен: он устойчив к нагреванию (время полужизни при 55°С составляет около 2 ч) и к действию детергентов. В процессе замораживания-оттаивания потери активности GUS не происходит. В составе химерных белков, созданных генно-инженерными методами, GUS обычно сохраняет свою функциональную активность. В живых клетках белок GUS также весьма стабилен и активен от нескольких часов до нескольких суток.
GFP (green fluorescent protein - зеленый флюоресцентный белок, или белок зеленой флюоресценции) был обнаружен Shimomura с соавт. в 1962 г. у люминесцирующей медузы Aequorea victoria. Ген GFP был клонирован в 1992 г. Prasher и соавт., и уже через несколько лет началось активное использование этого гена как репортерного в работах с самыми разными про- и эукариотическими организмами. В настоящее время ген GFP применяется в сотнях работ во всем мире, и число их стремительно нарастает. Столь быстрый рост вызван особыми свойствами белка GFP, а именно его способностью флюоресцировать в видимой (зеленой) области спектра при облучении длинноволновым УФ. Эта флюоресценция обусловлена непосредственно белком, для ее проявления не требуется субстратов или кофакторов. Благодаря этому свойству ген GFP является очень перспективным репортерным геном, позволяющим проводить разнообразные прижизненные (недеструктивные) исследования с трансгенными организмами.
Многочисленные производные GFP получили общее название AFP (autofluorescent proteins - автофлюоресцентные белки). Из морской анемоны Discosoma sp. недавно выделен еще один белок DsRed, флуоресцирующий в красном свете. Еще несколько аналогичных флюоресцирующих белков было выделено в самое последнее время учеными Российской академии наук из различных коралловых полипов порядка Anthozoa. Он может быть денатурирован очень высокой температурой, крайними значениями рН или сильными восстановителями типа Na2SO4. При возвращении к физиологическим условиям GFP в значительной степени восстанавливает способность к флюоресценции. В составе химерных белков, созданных генноинженерными методами, GFP обычно сохраняет свою функциональную активность. В живых клетках белок GFP также очень стабилен.
CAT – гены отвечают за синтез хлорамфениколацетилтрансферазы (выделены из Escherihia coli). Этот фермент катализирует реакцию переноса ацетильной группы от ацетил-КоА к хлорамфениколу. Определяется гистохимически, по изменению окраски ткани при добавлении соответствующего субстрата.
LUC – ген кодирует фермент люциферазу (клонирована из бактерий и светлячка). Она вызывает свечение трансформированных клеток. Бактериальный фермент состоит из двух субъединиц. Для определения активности ферментов необходимо специальное оборудование - флуориметр и цифровая видеокамера с амплификатором светового сигнала. Фермент теряет активность при действии детергентов и повышенной температуры. Замена селективных генов на репортерные при отборе трансгенных растений часто весьма желательна, так как возможность потенциального риска для окружающей среды и здоровья человека при использовании репортерных генов практически исключена. Однако область применения репортерных генов шире, чем просто контроль трансгеноза. Другое, и, очевидно, более важное назначение репортерных генов состоит в том, чтобы выявлять (по возможности количественно) временные и пространственные особенности экспрессии данного конкретного гена, будь то собственного или чужеродного. Присоединение репортерного гена к одной лишь промоторной области позволяет исследовать в "чистом виде" ее роль в регуляции экспрессии изучаемого гена на уровне транскрипции.
Замена белок-кодирующей области гена на репортерную при сохранении участка, кодирующего 5'-концевую не транслируемую последовательность мРНК, позволяет оценить роль этой последовательности в процессах транспорта мРНК из ядра в цитоплазму и инициации трансляции.
Одно из самых важных свойств гена - способность к экспрессии. За это свойство отвечают различные генетические элементы, которые мы должны встроить в векторную молекулу, несущую ген.
2.1 Регуляция экспрессии гена у прокариот
Многие бактериальные гены устроены таким образом, что они способны функционировать с существенно разной эффективностью. У E. coli, например, относительное содержание различных белков варьирует в очень широких пределах (от менее чем 0.1% до 2%) в зависимости от их функций; при этом каждый белок в хромосоме E. coli кодируется единственным геном. Такие вариации обусловлены действием системы контроля генной экспрессии, которая осуществляется главным образом на уровне транскрипции ДНК. Таким образом, чаще всего уровень активности гена связан с количеством синтезируемой на нем мРНК, то есть с активностью фермента РНК-полимеразы.
Последовательности ДНК, расположенные перед началом структурного гена и определяющие степень активности РНК-полимеразы, называются регуляторными последовательностями. Одна из таких последовательностей представляет собой участок ДНК, с которым связывается РНК-полимераза. Этот участок называется промотором.
Последовательность оснований промотора определяет частоту инициации синтеза иРНК, причем замена одного основания в этой последовательности может привести к уменьшению частоты инициации в 1000 раз.
Промотор может быть сильным и слабым. Сильный промотор инициирует синтез иРНК часто, слабый - гораздо реже. С другой стороны, промотор может быть регулируемым и нерегулируемым. Например, промотор β-лактамазы нерегулируемый, но сильный. Использование таких промоторов не всегда удобно. Дело в том, что большое количество белка может блокировать рост бактерий. Кроме того, интенсивная транскрипция рекомбинантной ДНК может помешать репликации плазмиды, и она будет утрачена. Поэтому удобнее использовать регулируемые сильные промоторы (индуцибельные), включение которых, а значит и синтез чужеродного белка можно осуществить, когда получена большая бактериальная масса.
Некоторые плазмидные векторы содержат промотор, работа которого регулируется температурочувствительным белковым продуктом гена-репрессора. Белок-репрессор активен при определенных температурах и блокирует действие промотора. Повысив температуру до 42 оС, можно "включить" промотор и быстро получить большое количество требуемого белка.
В качестве индуцибельных промоторов используют также Trp-промотор триптофанового оперона, который регулируется триптофановым голоданием, lac-промотор лактазного оперона, который индуцируется субстратом (лактозой) и другие.
Интенсивность транскрипции определенных структурных генов может зависеть от эффективности ее терминации, в частности, от того, как часто РНК-полимераза прекращает синтез РНК, не дойдя до этих генов. Сравнительно недавно обнаружено, что во многих оперонах Е.coli, контролирующих биосинтез аминокислот, между промотором и первым структурным геном имеется терминирующая последовательность. В определенных условиях происходит образование терминирующего сигнала, ослабляющего интенсивность транскрипции.
Это явление получило название аттенуации, а участок ДНК - аттенуатор (ослабитель). Как и репрессия, аттенуация зависит от присутствия в среде соответствующих аминокислот. Например, избыток триптофана в клетках триптофанзависимых мутантов, дефектных по репрессору, только 1 из 10 молекул РНК-полимеразы, начавших транскрипцию, преодолевает аттенуатор и считывает структуру генов. Удаление триптофана втрое повышает эффективность транскрипции генов. В отличие от репрессии, антенуация зависит не от самой аминокислоты, а от триптофанил - тРНК (аминокилоты, присоединенной к соответствующей тРНК).
На эффективность продуктивности рекомбинантной ДНК в существенной степени влияет количество копий этой ДНК в расчете на клетку. Суммарная активность экспрессируемого гена растет с ростом копийности плазмиды. Таким образом, используя многокопийные плазмиды, можно достичь сверхсинтеза нужных белковых продуктов. Обычно используемые плазмидные векторы (pBR 322 и др.) поддерживаются в клетке в количестве 20-50 копий. Сейчас исследователи имеют в своем распоряжении температурно-чувствительные мутантные плазмиды, способные накопить до одной-двух тысяч копий на клетку, не нарушая ее жизненно-важных функций. Таким образом можно достичь сверхпродукции плазмидных генов бактериальными штаммами-сверхпродуцентами.
Регуляция экспрессии у E. coli происходит также и на уровне трансляции. Последовательность оснований длиной 6-8 нуклеотидов, расположенная непосредственно перед инициирующим кодоном АУГ, определяет эффективность трансляции. Эта последовательность представляет собой участок связывания мРНК с рибосомой. Как правило, он отстоит на 8 нуклеотидов от инициирующего кодона, и его сдвиг в ту или иную сторону может резко снижать эффективность трансляции соответствующей мРНК. Описанный участок называется последовательностью Шайна-Дальгарно, по имени исследователей, впервые его идентифицировавших.
В состав вектора кроме всего прочего должен входить маркерный ген, позволяющий селектировать измененные клетки. Часто в качестве селективных используют широко распространенные в природе гены ферментов, модифицирующих антибиотики и инактивирующие их действие.
Особенности организации генома эукариот
У эукариотических организмов механизм регуляции транскрипции гораздо более сложен. В результате клонирования и секвенирования генов эукариот обнаружены специфические последовательности, принимающие участие в транскрипции и трансляции.
Для эукариотической клетки характерно:
1. Наличие интронов и экзонов в молекуле ДНК.
2. Созревание и-РНК - вырезание интронов и сшивка экзонов.
3. Наличие регуляторных элементов, регулирующих транскрипцию, таких как: а) промоторы - 3 вида, на каждый из которых садится специфическая полимераза. Pol I реплицирует рибосомные гены, Pol II - структурные гены белков, Pol III - гены, кодирующие небольшие РНК. Промотор Pol I и Pol II находятся перед участком инициации транскрипции, промотор Pol III - в рамках структурного гена; б) модуляторы - последовательности ДНК, усиливающие уровень транскрипции; в) усилители - последовательности, усиливающие уровень транскрипции и действующие независимо от своего положения относительно кодирующей части гена и состояния начальной точки синтеза РНК; г) терминаторы - специфические последовательности, прекращающие и трансляцию, и транскрипцию.
Эти последовательности по своей первичной структуре и расположению относительно инициирующего кодона отличаются от прокариотических, и бактериальная РНК-полимераза их не "узнает". Таким образом, для экспрессии эукариотических генов в клетках прокариот нужно, чтобы гены находились под контролем прокариотических регуляторных элементов. Это обстоятельство необходимо учитывать при конструировании векторов для экспрессии.
2.2 Способы прямого введения гена в клетку
Прямое введение гена в клетку осуществляют несколькими способами:
1. Трансфекция
2. Микроинъекция
3. Электропорация
4. Метод «мини-клеток»
5. Упаковка в липосомы
6. Электронная пушка
При трансфекции ДНК адсорбируется на кристаллах фосфата кальция (Грэхем Ван дер Эб, 1973). Образуются частицы кальциевого преципитата. Они поглощаются клеткой путем фагоцитоза.
Для повышения эффективности трансформации к специфической ДНК, содержащей ген, по которому будет производится селекция, добавляется неспецифическая ДНК-носитель. Обычно для этой цели берут ДНК из тимуса теленка или спермы лосося. Часть ДНК связывается с мембраной и не попадает в клетки. ДНК акцептируют от 15 до 90% клеток. Через несколько суток после введения небольшая доля клеток способны экспрессировать чужеродные гены, но затем уровень экспрессии падает и более или менее стабильную трансформацию претерпевает 10-3 - 10-5 клеток.
Для трансфекции используется и ДЭАЭ-декстран, полимер, адсорбирующий ДНК. Эффект вхождения в клетки и время экспрессии высоки, но частота стабильной трансформации ниже, чем при использовании преципитата кальция. Частоту трансфекции увеличивает глицериновый шок (4 минуты в 15% растворе глицерина в НEPES-буфере).
В клетки можно вводить любой ген, если заранее лигировать его с клонированным селективным маркером. Однако дальнейшие исследования показали, что лигирование вне клетки не обязательно. Клетки, поглощающие селективный ген, вместе с ним поглощают и другую ДНК, имеющуюся в кальциевом преципитате. Таким образом, пользуясь методом котрансформации, практически любой клонированный сегмент ДНК можно ввести в культивируемые клетки эукариот, если включить эту ДНК вместе с селективным маркером в состав смеси для образования кальциевого преципитата.
Для трансфекции можно использовать хромосомы или фрагменты хромосом. Клетки-доноры блокируются на стадии митоза. Митотические хромосомы высвобождаются под воздействием осмотического шока и гомогенизации. Их очищают путем дифференциального центрифугирования. Хромосомы осаждают на поверхности клеток хлористым кальцием, а через несколько часов обрабатывают реагентом, способным перфорировать мембраны (например, глицерином).
Для обработки клеток-рецепиентов используются грубо очищенные препараты хромосом, так как хромосомы при этом разрушаются меньше всего. Количество хромосом для обработки 1 клетки ограничено. Лучше использовать не более 20 хромосом на 1 клетку-рецепиент, так как при высоких концентрациях хромосом в суспензии они агглютинируют. Рецепиентная клетка содержит фрагменты донорных хромосом, которые могут встраиваться в геном, могут реплицироваться самостоятельно. Во введенных фрагментах часто наблюдаются делеции.
Не все клетки способны к трансформации геномной ДНК с высокой частотой. Человеческие фибробласты эффективно включают плазмидную ДНК и почти не включают геномную.
Микроинъекция ДНК в клетки млекопитающих стала возможной с появлением прибора для изготовления микропипеток диаметром 0.1-0.5 микрона и микроманипулятора (рис. 45). Так, плазмиды, содержащие фрагмент вируса герпеса с геном тимидинкиназы (ТК) и плазмиду рВR322, были инъецированы в ТК--клетки и было показано, что ТК-ген проник в ядра и нормально в них реплицировался. Метод введения ДНК с помощью микроинъекций был разработан в начале 70-х годов Андерсоном и Диакумакосом. В принципе, при наличии хорошего оборудования можно за 1 час инъецировать 500-1000 клеток, причем в лучших экспериментах в 50% клеток наблюдается стабильная интеграция и экспрессия инъецированных генов. Преимущество описываемого метода заключается также в том, что он позволяет вводить любую ДНК в любые клетки, и для сохранения в клетках введенного гена не требуется никакого селективного давления.
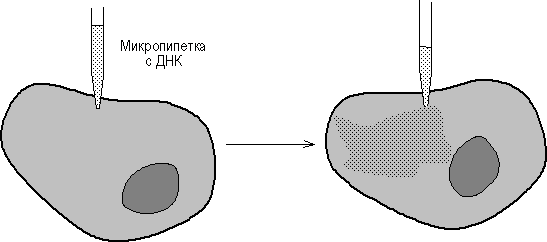
Рис. 4. Введение ДНК путем микроинъекции
Электропорация основана на том, что импульсы высокого напряжения обратимо увеличивают проницаемость биомембран. В среду для электропорации добавляют клетки и фрагменты ДНК, которые необходимо ввести в клетки (рис. 46). Через среду пропускают высоковольтные импульсы (напряжение 200 - 350 В, длительность импульса 54 мс), приводящие к образованию пор (электропробой) в цитоплазматической мембране, время существования и размер которых достаточны, чтобы такие макромолекулы, как ДНК, могли из внешней среды войти в клетку в результате действия осмотических сил. При этом объем клетки увеличивается.
Напряженность электрического поля и продолжительность его действия, концентрации трансформирующей ДНК и реципиентных клеток для каждой системы клеток подбирают экспериментально, с тем чтобы достичь высокого процента поглощения ДНК выжившими клетками. Показано, что в оптимальных условиях электропорации количество трансформантов может достигать 80% выживших клеток.
Электропорация — физический, а не биохимический метод, и это, по-видимому, обусловливает его широкое применение. Многочисленные исследования продемонстрировали, что электропорация может успешно использоваться для введения молекул ДНК в разные типы клеток, такие как культивируемые клетки животных, простейшие, дрожжи, бактерии и протопласты растений. Электропорирующий эффект высоковольтного разряда на бислойную липидную мембрану, по-видимому, зависит от радиуса ее кривизны. Поэтому мелкие бактериальные клетки эффективно поглощают ДНК при значительно большей напряженности (10 кВ/см и более), чем крупные животные и растительные клетки, эффективно поглощающие ДНК при напряженности поля 1—2 кВ/см.
Электропорация — наиболее простой, эффективный и воспроизводимый метод введения молекул ДНК в клетки. Однако до недавнего времени этот метод использовался в ограниченном числе лабораторий в связи с отсутствием серийных приборов — электропораторов. Появление и совершенствование таких приборов в ближайшие годы приведет к широкому применению данного подхода в генетической инженерии самых разных типов клеток.

Рис. 5. Метод электропорации
«Мини-клетки» получают путем блокирования донорных клеток митозе колцемидом. При продолжительной обработке клеток колцемидом в них вокруг каждой хромосомы формируется новая ядерная мембрана. Обработка цитохалазином В и центрифугирование приводит к образованию мини-клеток, представляющих микроядра, инкапсулированные в цитоплазматическую мембрану.
Полученные мини-клетки очень чувствительны к разного рода воздействиям, поэтому для слияния подбирают специальные мягкие условия. Метод трудный, капризный, эффективность низкая – 10-6 – 10-7.
Упаковка в липосомы используется для защиты экзогенного генетического материала от разрушающего действия рестриктаз.
Липосомы - сферические оболочки, состоящие из фосфолипидов. Получают их путем резкого встряхивания смеси водного раствора и липидов, либо обрабатывая ультразвуком водные эмульсии фосфолипидов. Липосомы, состоящие из фосфатидилсерина и холестерина наиболее пригодны для введения ДНК в клетки животных и растений. Системы переноса с помощью липосом низкотоксичны по отношению к клеткам.
Метод биологической баллистики (биолистики) является одним из самых эффективных на сегодняшний день методов трансформации растений, особенно однодольных.
Суть метода заключается в том, что на мельчайшие частички вольфрама, диаметром 0,6—1,2 мкм, напыляется ДНК вектора, содержащего необходимую для трансформирования генную конструкцию. Вольфрамовые частички, несущие ДНК, наносятся на целлофановую подложку и помещаются внутрь биолистической пушки. Каллус или суспензия клеток наносится в чашку Петри с агаризированной средой и помещается под биолистическую пушку на расстоянии 10—15 см. В пушке вакуумным насосом уменьшается давление до 0,1 атм. В момент сбрасывания давления вольфрамовые частички с огромной скоростью выбрасываются из биолистической пушки и, разрывая клеточные стенки, входят в цитоплазму и ядро клеток.
Обычно клетки, располагающиеся непосредственно по центру, погибают из-за огромного количества и давления вольфрамовых частиц, в то время как в зоне 0,6—1 см от центра находятся наиболее удачно протрансформированные клетки. Далее клетки осторожно переносят на среду для дальнейшего культивирования и регенерации.
С помощью биолистической пушки были протрансформированы однодольные растения, такие, как кукуруза, рис, пшеница, ячмень. При этом были получены стабильные растения-трансформанты. Кроме успехов в получении трансгенных однодольных, биолистическая трансформация применяется для прямого переноса ДНК в эмбриогенную пыльцу и дальнейшего быстрого получения трансгенных дигаплоидных растений, которые являются важным этапом в селекционной работе. В настоящее время этим методом была проведена трансформация растений табака и после регенерации гаплоидных растений получены стабильные трансформанты.
2.3 Способы прямого введения гена в клетку
В настоящее время бактерия E. coli является самой изученной клеткой из всех существующих. У большинства наиболее полно изученных фагов клеткой - хозяином является также E. coli.
Протопласт E. coli одет в муреиновый мешок, прилегающий к внешней мембране. E. coli относится к микроорганизмам, не обладающим физиологической компетентностью к поглощению экзогенной ДНК. Поэтому необходимо создать условия, позволяющие преодолеть барьер клеточной стенки. Сначала получают сферопласты путем обработки клеток лизоцимом в изотоническом растворе.
Липополисахаридный слой внешней мембраны грамотрицательной бактерии стабилизирован двухвалентными катионами, поэтому для разрыхления внешней мембраны E. coli используется комплексообразователь этилендиаминтетрауксусная кислота (EDTA), которая связывает двухвалентные катионы. При обработке EDTA часть липополисахаридов высвобождается из внешней мембраны клетки, и лизоцим может достигнуть муреинового мешка и гидролизовать его. Это ведет к повышению проницаемости клеточной оболочки. Усовершенствование методов получения сферопластов E. coli и их трансфекции позволили достичь достаточно высокой эффективности трансформации молекулами ДНК различных фагов.
Обнаружено, что на инфекционность существенное влияние оказывает форма молекул фаговых ДНК, которую они принимают in vivo. Фаги с кольцевой или линейной, но быстро замыкающейся ДНК (лямбоидные фаги) характеризуются наибольшей эффективностью трансфекции.
Успешное проведение экспериментов на кишечной палочке стало стимулом для проведения аналогичных исследований с другими прокариотическими организмами. Наибольших успехов удалось достичь с клетками Bacillus subtilis. B. subtilis - непатогенный почвенным микроорганизм, растущий в строго аэробных условиях. Бациллы не образуют токсинов и непатогенны ни для животных, ни для человека, тогда как клеточная стенка E. coli содержит эндотоксин, который довольно трудно отделить от продуктов генной инженерии. Кроме того, клеточная стенка бацилл имеет простую структуру и бактерии могут секретировать многие белки в культуральную жидкость. 20 различных видов бацил секретируют в культуральную жидкость более 40 ферментов с внеклеточной локализацией. E. coli секретирует в среду относительно мало белков, а выделение и очистка их затруднены. В бациллах также обнаружены плазмиды и фаги, которые к настоящему моменту уже хорошо изучены.
Чужеродные гены клонируют в так называемых челночных векторах. Эти вектора с одинаковым успехом реплицируются в клетках нескольких хозяев, в данном случае, в клетках E. coli и B.subtilis. Векторы были получены комбинацией in vitro фрагментов этих плазмид.
Гены E. coli со своими регуляторными районами не функционируют в B.subtilis, поэтому были использованы собственные гомологичные районы B.subtilis.
Для конструирования рекомбинантной ДНК, содержащей в своем составе ген, который должен экспрессироваться, придерживаются следующей стратегии. Синтезируют кДНК или из клонотеки выделяют клетки, несущие фрагмент генома с нужным геном, и клонируют их в соответствующем векторе. Фрагменты геномной ДНК подвергают модификации - удаляют из них некодирующие области и участки соседних генов. Часто для проведения этой операции необходимо секвенирование данного фрагмента ДНК. Затем конструируются промежуточные рекомбинантные ДНК, в которых ген помещается под контроль бактериальных регуляторных элементов (промотор, оператор, точка связывания с рибосомами). Эти регуляторные элементы выделяют из гибридных плазмид, сконструированных специально как источники регуляторных элементов. Полученная конструкция встраивается в подходящий вектор, например, pBR 322, и ген экспрессируется в бактериальной клетке.
Однако удобнее встраивать ген в специальный вектор для экспрессии, который уже содержит регуляторные элементы, обеспечивающие активную экспрессию после введения рекомбинантной плазмиды в бактериальную клетку. К таким эффективным регуляторным участкам относится, например, сильный промотор гена бэта-лактамазы (ген устойчивости к пенициллину, входящий в состав плазмиды pBR 322). Ряд генов, в том числе и ген инсулина, встраивали в сайт рестрикции Pst I, который расположен в структурной части гена. Промотор этого гена обеспечивает эффективную транскрипцию, которая продолжается до тех пор, пока РНК-полимераза не дойдет до сигнала терминации встроенного гена.
В качестве примера маркирования вектора могут служит первые эксперименты с E. coli, а точнее с одной из ее плазмид рBR322, проведенные Гилбертом для получения инсулина. Плазмида pBR322 содержит 2 гена, которые определяют устойчивость к ампициллину и тетрациклину. Рестриктаза PstI расщепляет плазмиду в средней части гена, кодирующего фермент устойчивости к апициллину. После расщепления плазмиды на ее концы с помощью концевой трансферазы надстраивали последовательность из четырех нуклеотидов с остатками гуанина. Затем, как обычно, с помощью лигаз "вшивали" ген проинсулина, получая рекомбинантную ДНК. Встроенный в плазмиду фрагмент ДНК нарушал синтез фермента, разрушающего ампициллин, но ген, обеспечивающий устойчивость к тетрациклину, оставался активным. Трансформированные таким образом клетки E. coli синтезировали гибридный белок, содержащий последовательности пенициллазы и проинсулина, поэтому биологически активный инсулин получали путем отщепления пенициллазы и средний сегмент проинсулина.
С другой стороны, если фрагмент чужеродной ДНК встраивается в один из генов устойчивости, то последний инактивируется. Следовательно, успешное встраивание фрагмента чужеродной ДНК в один из этих генов легко детектировать по исчезновению у бактерий устойчивости к данному антибиотику.
2.4 Введение генов в клетки млекопитающих
Манипуляции с клетками млекопитающих можно разделить на 2 большие группы: эксперименты с соматическим клетками и эксперименты по трансформации половых клеток. В последнем случае конечный результат – получение трансгенных организмов.
Характеристика векторов для переноса генов в животные клетки
Одними из лучших носителей для введения чужеродной информации в животную клетку являются вектора на основе ретровирусов, например, на основе вируса лейкоза мышей. Они обеспечивают высокоэффективный перенос генов и их стабильное встраивание в хромосому клеток-мишеней. В основном трансформации животных клеток осуществляют либо с помощью ретровирусов (около 40% от всех трансформаций), либо путем упаковки ДНК в липосомы (25%), реже используют аденовирусы, так как они могут вызывать сильный иммунный ответ, кроме того, невозможно их повторное введение.
Если проблема доставки чужеродной ДНК in vitro практически решена, а ее доставка в клетки-мишени разных тканей in vivo успешно решается (главным образом путем создания конструкций, несущих рецепторные белки, в том числе и антигены, специфичные для тех или иных тканей), то другие характеристики существующих векторных систем - стабильность интеграции, регулируемая экспрессия, безопасность — все еще нуждаются в серьезных доработках.
Прежде всего это касается стабильности интеграции. До настоящего времени интеграция в геном достигалась только при использовании ретровирусных либо аденоассоциированных векторов. Повысить эффективность стабильной интеграции можно путем совершенствования генных конструкций типа рецептор-опосредованных систем, либо путем создания достаточно стабильных эписомных векторов (то есть ДНК-структур, способных к длительной персистенции внутри ядер).
В последнее время особое внимание уделяется созданию векторов на базе искусственных хромосом млекопитающих (MAC - mammalian artificial chromosomes). Благодаря наличию основных структурных элементов обычных хромосом такие мини-хромосомы длительно удерживаются в клетках и способны нести полноразмерные (геномные) гены и их естественные регуляторные элементы, которые необходимы для правильной работы гена, в нужной ткани и в должное время. Такие искусственные хромосомы уже созданы для дрожжей (YAK), так как геном дрожжей полностью картирован.
Для идентификации модифицированных клеток, необходимы маркеры. Если трансформируют соматические клетки, то применяют обычно селективные маркеры. Аксель с коллегами из колледжа терапии и хирургии Колумбийского университета исправили таким образом генетический дефект клеток мыши. Они взяли фрагмент ДНК, содержащий ген тимидинкиназы (ТК), который получен из вируса герпеса, смешали эту ДНК с несколькими миллиграммами ДНК-носителя из спермы лосося и осадили ДНК на культуру L-клеток мыши, в которых ген ТК отсутствовал (ТК-). С частотой 1 на 100000 клетки приобретали ген ТК, поэтому на селективной среде, которая не позволяла расти ТК- - клеткам, росли и нормально размножались ТК+ - клетки.
Другой селективный маркер - ген, кодирующий дигидрофолатредуктазу (ДГФР), можно использовать при трансформации немутантных линий клетки. Благодаря экспрессии многих копий этого гена животная клетка вместе с плазмидой приобретает устойчивость к высоким концентрациям ингибитора фермента, и таким образом трансформантов можно отбирать при высоких концентрациях ингибитора.
Разработано еще два универсальных вектора, содержащих генные маркеры, работающие в нормальных клетках. Они построены по одному и тому же принципу: прокариотические гены, определяющие фенотип трансгенных клеток, соединены с эукариотическими регуляторными сигналами.
Один из векторов состоит из прокариотического гена устойчивости к антибиотику неомицину, встроенного в раннюю область генома SV-40. Эукариотические клетки чувствительны к аналогу неомицина G 418, который инактивируется продуктом гена. Таким образом клетки, прошедшие трасфекцию приобретают способность расти на среде, содержащей G 418.
2.5 Генетическая транформация соматических клеток млекопитающих
Культуры трансформированных клеток млекопитающих используют для получения различных веществ. Хотя культуры клеток животных, особенно при массовом выращивании, гораздо менее экономичны, чем бактериальные дрожжевые культуры, они обладают существенным преимуществом - способностью осуществлять мелкие, но весьма важные модификации белков - продуктов гена млекопитающих. Например, для эффективного функционирования ряда белков необходимо присоединение к ним цепочек из молекул углеводов или липидов. Образование и присоединение таких цепочек - обычный процесс для клеток млекопитающих, тогда как бактериальная клетка не способна производить подобные модификации.
Помимо создания клеток-продуцентов, трансформация соматических клеток млекопитающих позволяет изучать тонкие механизмы регуляции экспрессии генов и целенаправленно модифицировать генетический аппарат клетки животных, а при необходимости и человека, что имеет огромное значение для медицинской генетики.
Культуры клеток млекопитающих могут оказаться эффективным источником выделения некоторых вирусных антигенов с целью получения вакцин для животных и человека. Получение таких вакцинных культур клеток осуществимо при помощи техники рекомбинантных ДНК и эффективных векторов экспрессии для клеток млекопитающих и человека. При использовании ДНК-вакцин в организм вводится не антиген, а ген, кодирующий синтез этого антигена. Ген встраивается в плазмиду, а плазмида вводится организм путем обыкновенной инъекции.
ДНК-вакцины имеют хорошие перспективы в животноводстве. Фибер – белок вирусной оболочки. Эпитоп фибера кодирует синтез протективных антител. Одно из заболеваний птиц – синдром снижения яйценоскости (ССЯ) вызывается вирусом. После анализа ДНК этого вируса был выделен ген, кодирующий фибер, проклонирован и встроен в плазмиду. Рекомбинантная вакцина при введении ее в организм принесет ДНК фибера в клетку, выработка вирусного белка спровоцирует синтез специфических антител, т. е. вызовет иммунный ответ.
Достоинством таких вакцин является очень маленький объем – для иммунизации одной мыши достаточно 10-50 мкг плазмиды, одной коровы – 200-300 мкг. Плазмида сохраняется в организме до 1 года. В стадии клинических испытаний в настоящее время находятся ДНК-вакцины против микоплазм, возбудителя туберкулеза, сальмонеллеза, лейшманиоза.
Развитие злокачественной опухоли в организме обычно подавляет иммунитет. Проблема в том, чтобы подхлестнуть иммунную систему в целом и направить ее действие против раковых клеток. Исследователи из Медицинской школы в Энн-Арборе (Мичиган) придумали метод борьбы с раком. В опухолевые клетки толстой кишки подопытных мышей ввели гены, кодирующие белки другой линии мышей. Это можно осуществить с помощью липосом или вируса. После появления на внешней стороне клеточной мембраны этих белков иммунная система атаковала такие клетки. 20% больных мышей выздоровели, у 70% опухоль уменьшилась, в контрольной группе все умерли. Лимфоциты боролись не только с «меченными» клетками опухоли, но и клетками метастаз, следовательно, иммунная система «проснулась». В настоящее время ведутся эксперименты на людях с раком кожи.
2.6 Генотерапия
Лечение заболеваний с помощью генов получило название генотерапии. Сейчас в мире насчитывается порядка 400 проектов, посвященных лечению с помощью генотеропии.
Разработке программы генной терапии предшествуют тщательный анализ тканеспецифической экспрессии соответствующего гена, идентификация первичного биохимического дефекта, исследование структуры, функции и внутриклеточного распределения его белкового продукта, а также биохимический анализ патологического процесса. Все эти данные учитываются при составлении соответствующего медицинского протокола.
Апробацию процедуры генокоррекции наследственного заболевания проводят на первичных культурах клеток больного, в которых в норме функционально активен данный ген. На этих клеточных моделях оценивают эффективность выбранной системы переноса экзогенной ДНК, определяют экспрессию вводимой генетической конструкции, анализируют ее взаимодействие с геномом клетки, отрабатывают способы коррекции на биохимическом уровне. Используя культуры клеток, можно разработать систему адресной доставки рекомбинантных ДНК, однако проверка надежности работы этой системы может быть осуществлена только на уровне целого организма. Поэтому такое внимание в программах по генной терапии уделяется экспериментам in vivo на естественных или искусственно полученных моделях соответствующих наследственных болезней у животных.
Успешная коррекция генетических дефектов у таких животных и отсутствие нежелательных побочных эффектов генной терапии являются важнейшей предпосылкой для разрешения клинических испытаний. Таким образом, стандартная схема генокоррекции наследственного дефекта включает серию последовательных этапов. Она начинается созданием полноценно работающей (экспрессирующейся) генетической конструкции, содержащей смысловую (кодирующую белок) и регуляторную части гена. На следующем этапе решается проблема вектора, обеспечивающего эффективную, а по возможности и адресную доставку гена в клетки-мишени. Затем проводится трансфекция (перенос полученной конструкции) в клетки-мишени, оценивается эффективность трансфекции, степень коррегируемости первичного биохимического дефекта в условиях клеточных культур (in vitro) и, что особенно важно, in vivo на животных - биологических моделях. Только после этого можно приступать к программе клинических испытаний.
Существует два типа генотерапии: заместительная и корректирующая.
Заместительная генотерапия заключается во вводе в клетку неповрежденного гена. Внесенная копия заменит по функциям сохранившийся в геноме больного дефектный ген. Все проводимые сегодня клинические испытания используют внесение в клетку дополнительных количеств ДНК.
При корректирующей терапии предполагается замена дефектного гена нормальным в результате рекомбинации. Пока этот метод на стадии лабораторных испытаний, так как эффективность его еще очень низка.
В зависимости от способа введения экзогенных ДНК в геном пациента генная терапия может проводиться либо в культуре клеток (ex vivo), либо непосредственно в организме (in vivo). Клеточная генная терапия или терапия ex vivo предполагает выделение и культивирование специфических типов клеток пациента, введение в них чужеродных генов, отбор трансфецированных клеток и реинфузию их тому же пациенту.
Примером может служить лечение комбинированного иммунодефииицита. Комбинированный иммунодефицит может быть результатом дефекта гена аденозиндезаминазы. Впервые попытка лечения такого больного методами генотерапии была предпринята в США в 1990 г. У больного ребенка извлекли Т-лимфоциты, трансформировали ретровирусным вектором, введя нормальный ген аденозиндезаминазы и вернули клетки в организм. Введение приходится повторять. Более эффективна аналогичная трансформация стволовых клеток костного мозга.
Генная терапия in vivo основана на прямом введении клонированных и определенным образом упакованных последовательностей ДНК в специфические ткани больного. В настоящее время не существует общедоступного метода культивирования клеток легких, поэтому при легочных заболеваниях единственный способ доставить чужеродный ген - это ввести его прямо в организм. Муковисцидоз - весьма распространенное среди людей белой расы тяжелое наследственное заболевание легких, которое поражает, например, в семьях из Центральной Европы одного новорожденного из 2500 и для которого установлен дефектный ген, кодирующий белок-регулятор трансмембранной проводимости. Основное проявление дефектного гена – пневмония. Поражаются все эпителиальные клетки. Основная проблема – как доставить ген в клетки, покрытые слизью, которая препятствует трансформации. Неповрежденную копию "гена заболевания", включенную в аденовирусный вектор или липосому, вводят в форме аэрозоля в дыхательные пути больного.
Для коррекции нарушения при прогрессирующей мышечной дистрофии Дюшенна (заболевании мальчиков, связанном с дефектами Х-хромосомы) нормальный ген, кодирующий белок дистрофии, пытались прямо вкалывать в мышечные волокна, используя либо "голую" ДНК, либо аденовирусный вектор. Другие исследователи трансплантировали больному миобласты после генетической коррекции. Ранее неподвижный ребенок приобретал способность двигаться! К сожалению, во всех этих опытах удается получить только временный терапевтический эффект, и процедура введения гена должна неоднократно повторяться.
Список наследственных заболеваний, которые пытаются или планируют лечить генами, велик. Это и ревматоидный артрит, и фенилкетонурия, и заболевания, связанные с недостатком гормонов (инсулина, эритропоэтина, гормона роста). В случае хронической анемии, связанной с дефицитом эритропоэтина, на основании опытов на животных предлагается принципиально новый подход к лечению. Так как каждая из наших клеток содержит один и тот же геном, можно заставить фибробласты кожи, которые в норме не производят эритропоэтина, синтезировать этот гормон. Для этого нужно ввести в геном новую контролирующую область и тем самым снять запрет со считывания (экспрессии) гена эритропоэтина, присутствующего, но "молчащего" в фибробластах.
Практически в любой области медицины либо начаты клинические испытания лечения наследственных заболеваний с помощью генотерапии, либо в опытах на животных разрабатываются подходы к такому лечению. По мере усовершенствования методов доставки генов и контроля их экспрессии список заболеваний, к которым можно применять генотерапию, будет безусловно расширяться.
Генотерапия применима не только к наследственным заболеваниям. Предстоит решить проблему лечения генами "чумы XX века" — синдрома приобретенного иммунодефицита (СПИД), возникающего при заражении вирусом иммунодефицита человека (ВИЧ). ВИЧ представляет собой ретровирус, поражающий Т-лимфоциты и макрофаги. Болезнь удалось бы победить, если бы были найдены новые гены, введение которых в зараженные ВИЧ лимфоциты останавливало бы дальнейшее размножение вируса. Предложено множество хитроумных способов борьбы со СПИДом с помощью привнесенных генов. Все они основаны на новейших данных о строении и функционировании генома ретровируса. Например, вводя прямо в мышцы больного ретровирусные векторы, несущие отдельные гены ВИЧ, ученые рассчитывали на то, что гены ВИЧ после внедрения в ДНК хромосом хозяина смогут дать информацию для синтеза вирусных белков и произойдет "противоСПИДная" иммунизация больного этими белками. Однако еще не получено ощутимых результатов, которые сулили бы успех в борьбе с вирусом дикого типа, коварство которого заключается в его изменчивости.
Огромные перспективы открывает использование генотерапии для лечения онкологических заболеваний. Многолетние усилия ученых привели к пониманию того, что рак — это генетическое заболевание и его развитие происходит многостадийно, в результате серии генетических нарушений, накапливающихся в клетке. Следовательно, каждый из таких отдельных генетических эффектов может стать точкой приложения генотерапевтического подхода.
2.7 Получение трансгенных животных
Если вводить ДНК в клетки многоклеточного организма, то результатом трансформации будет изменение свойств лишь небольшого числа клеток, которые приобрели новый ген или гены. Следовательно, для изменения свойств всего организма следует изменять геном половых клеток, которые перенесут новые свойства потомкам. У растений и животных целесообразно изменять такие свойства, как скорость роста, устойчивость к заболеваниям, способность адаптироваться к новым внешним условиям. В качестве маркеров в этом случае можно использовать полиморфизм длины рестрикционных фрагментов (AFLP), анализ мини-сателлитов, анализ микросателлитной ДНК (SSR), гибридизацию и т.д.
Разработаны способы введения генов в эмбриональные клетки млекопитающих, мух и некоторых растений. От работы с довольно крупными яйцами амфибий перешли к изучению яйцеклеток и эмбрионов мыши, которая представляет наиболее изученное в генетическом отношении млекопитающее.
Микроинъекцию клонированных генов производят в один или оба пронуклеуса только что оплодотворенной яйцеклетки мыши. Чаще выбирают мужской пронуклеус, привнесенный сперматозоидом, так как его размеры больше. После инъекции яйцеклетку немедленно имплантируют в яйцевод приемной матери, или дают возможность развиваться в культуре до стадии бластоцисты, после чего имплантируют в матку.
Можно вводить ген в сперматозоиды и затем проводить ими оплодотворение. Таким образом были инъецированы гены интерферона и инсулина человека, ген β-глобина кролика, ген тимидинкиназы вируса простого герпеса и кДНК вируса лейкемии мышей. Число молекул, вводимое за одну инъекцию, колеблется от 100 до 300 000, а их размер - от 5 до 50 кб. Выживает обычно 10 - 30% яйцеклеток, а доля мышей, родившихся из трансформированных яйцеклеток варьирует от нескольких до 40%. Таким образом, реальная эффективность составляет около 10%.
Интеграция чужеродных генов неспецифична по отношению к хромосомам, а число копий чужеродного гена может различаться от нескольких штук до 100 и более. Эти гены образуют группу тандемных повторов, объединенных по типу "голова к хвосту". Чужеродная ДНК после инъекции была обнаружена как в соматических, так и в половых клетках. Это означает, что интеграция проходит на самых ранних стадиях развития зиготы.
В нескольких случаях гетерологичная ДНК наследовалась в трех поколениях мышей, что свидетельствует о стабильной интеграции. Интегрировавшая в половые клетки ДНК передается как менделевский ген. Установлено, что уровень экспрессии чужеродного гена зависит от места интеграции ДНК с хромосомами и от степени ее метилирования, а также от дифференцировки тканей. В некоторых случаях удалось получить тканеспецифическую экспрессию. Важно отметить что специфические чужеродные гены можно встраивать в геном клетки таким образом, что они подчиняются нормальным регуляторным сигналам.
В 1981 году Константини и Лэси (Оксфорд) провели инъекцию в яйцеклетки мыши фрагменты хромосомной ДНК кролика длиной 19 килобаз. Эти фрагменты содержали ген β-глобина кролика. Яйцеклетки культивировали до стадии бластоцисты и имплантировали в матку. У 24 мышей, родившихся в результате развития имплантированных яйцеклеток, проведены частичная гепатоэктомия. Анализ ДНК из клеток печени показал, что у 9 мышей встречается от 1 до 20 копий на клетку гена β-глобина. После спаривания 4 трансформированных самцов с нормальными самками получили потомство из 18 животных. 6 из них также имели ген β-глобина. Установлено, что интеграция гена в клетки млекопитающих происходит случайным образом и не связана с конкретными областями хромосомы. Ген нестабилен, может быть утрачен или стать неактивным. Вместе с геном необходимо вводить регуляторные последовательности.
Метод введения генов в эмбриональные клетки имеет ограничения. Не всегда удается встроить чужеродную ДНК в заданный участок хромосомы. Разработанные методические примы пока не позволяют заменить имеющийся в геноме ген, вытесняя его, не всегда удается подчинить новый ген системе регуляции организма.
При трансгенозе могут возникать неожиданные проблемы. Например, одни из первых работ по генетической транформации животных проводились путем встраивания генов гормона роста. Перенос гена гормона роста крысы мышам увеличивал рост мышей в 2 раза. Эксперименты по трансгенозу генов гормона роста быка кроликам также увенчались успехом. А вот аналогичные эксперименты по модификации крупного рогатого скота привели к увеличению прироста всего на 10-20%. Очевидно, это связанно с тем, что у мышей сохраняется широкая норма реакции, и встраивание генов, увеличивающих количество гормона, заставляет генотип реализоваться максимально полно. У домашнего скота в результате направленной селекции организмы работают на верхнем пределе нормы реакции, отсюда ожидаемый эффект не проявился.
В нашей стране получены свиньи, несущие ген соматотропина. Они не отличались по темпам роста от нормальных животных, но изменение обмена веществ сказалось на содержании жира. У таких животных ингибировались процессы липогенеза и активировался синтез белка. К изменению обмена веществ приводило и встраивание генов инсулиноподобного фактора. Такие трансгенные свиньи были созданы для изучения цепочки биохимических превращений гормона, а побочным эффектом явилось укрепление иммунной системы.
Самая мощная белоксинтезирующая система находится в клетках молочной железы. Если поставить гены чужих белков под контроль казеинового промотора, то экспрессия этих генов будет мощной и стабильной, а белок будет накапливаться в молоке (животное-ферментер). Уже получены трансгенные коровы, в молоке которых содержится человеческий белок лактоферрин. Этот белок планируют применять для профилактики гастроэнтерологических заболеваний у людей с низкой иммунорезистентностью. Это больные СПИДом, недоношенные младенцы, больные раком, прошедшие радиотерапию. Ведутся клинические испытания такого молока. Уже сейчас корпорация Genzyme Transgenics планирует исследования с целью создания трансгенного крупного рогатого скота, содержащего в молоке человеческий альбумин. Был куплен патент на получение эмбрионов, содержащих геном клеток соединительной ткани (фибробластов), включающий ген, ответственный за синтез человеческого белка. Подобная технология позволяет увеличить эффективность создания трансгенных молочных животных, так как при обычном впрыскивании генов в оплодотворенную яйцеклетку рождается от только 5 - 10% трансформированных животных, из них - несколько самцов, не дающих молока.
Использование новой технологии клонирования позволяет получать животных только женского пола, дающих трансгенный протеин. Альбумин используется в терапии для поддержания осмотического давления в крови. Ежегодно в мире требуется около 440 тысяч литров плазмы крови для выделения этого белка (стоимость около 1,5 млрд. $). Каждая молочная корова может произвести 80 кг рекомбинантного человеческого альбумина ежегодно. Genzyme Transgenics занимается разработкой аналогичных методов получения человеческого гормона роста и β-интерферона.
В Англии созданы трансгенные овцы, молоко которых содержит фактор свертывания крови.
В нашей стране были попытки создать овец, продуцирующих химозин (фермент для сыроварения). Было получено 2 овцы, у одной – ген не экспрессировался, у второй содержание химозина достигало 300 мг/л. Однако потомство этой овцы давало низкие удои – порядка 50 кг за период лактации. Причина заключалась в том, что химозин вырабатывается в виде предшественника – прохимозина, который превращается в активный фермент при рН=5. Было запланировано получать именно прохимозин, но в каких-то участках вымени происходило снижение рН, что приводило к активации химозина непосредственно в организме. Активный химозин свертывал молоко, а оно закупоривало протоки вымени. Сейчас пытаются решить эту проблему.
В Подмосковье получены кролики, выделяющие γ-интерферон, эритропоэтин, но кролики не являются традиционными продуцентами молока. Эксперименты же по трансформации сельскохозяйственных животных очень дорогостоящи – одно трансгенное животное стоит десятки и сотни тысяч долларов.
Трансгенных животных получают и для целей ксенотрансплантации. Одним из излюбленных доноров органов являются свиньи, так как имеется анатомическое сходство органов и сходство иммунологических свойств. Реакции отторжения при трансплантации имеют сложный механизм. Одним из сигналов для атаки организма на чужой орган являются белки, локализованные на внешней поверхности мембраны. У трансгенных свиней эти белки заменены на человеческие.
Еще одно направление трансгеноза – получение устойчивых к болезням животных. Животноводство держится на вакцинах, так как селекция ведется преимущественно на хозяйственно ценные признаки – шерстистость, молочность и т. д. Повышение устойчивости – дело генных инженеров. К защитным белкам относятся интерфероны, поэтому ген интерферона встраивали различным животным. Трансгенные мыши получили устойчивость, они не болели или болели мало, а вот у свиней такого эффекта не обнаружено.
Другое направление – введение генов, кодирующих антисмысловую РНК. Для животноводства острой проблемой являются лейкозы, вызываемые РНК-вирусами. Трансгенные кролики, несущие гены, отвечающие за присутствие в клетке антисмысловой РНК, были устойчивы к лейкозам.
Трансгенных животных можно использовать для изучения наследственных заболеваний мозга и нервной системы. Гены болезни Альцгеймера (отложение белка β-амилоида приводит к образованию характерных бляшек) и гены, отвечающие за развитие эпилепсии, болезней мозга вводятся в геном нормальных животных; при этом получают трансгенных животных-моделей, на которых можно испытывать различные терапевтические приемы.
Трансгенных животных стали использовать для исследования воспалительных и иммунологических заболеваний человека, например, ревматоидного артрита. Моделируются болезни, связанные с липидным обменом.
Заключение
Хотя генетика и генная инженерия уже играют огромную роль в медицине и сельском хозяйстве, основные результаты ещё впереди. Нам ещё очень многое предстоит узнать о том, как работает сложная генетическая система в нашем организме и у других видов живых существ.
Необходимо определить функции и назначение каждого гена, определить, каковы условия его активации, в какие периоды жизни, в каких частях тела и при каких обстоятельствах он включается и приводит к синтезу соответствующего белка. Далее, необходимо понять, какую роль играет в организме этот белок, выходит ли он за пределы клетки, какие сообщения несёт, какие реакции катализирует, как влияет на запуск биологических процессов в других частях организма, какие гены активирует. Отдельной сложной задачей является решение проблемы сворачивания белков - как, зная последовательность аминокислот, составляющих белок, определить его пространственную структуру и функции. Эта проблема требует новых теоретических знаний и более мощных суперкомпьютеров.
Но учёные не пасуют перед масштабом этой задачи. Расшифровка генома человека потребовала более десяти лет, решение проблемы сворачивания белков может занять чуть дольше, но когда она будет решена, человек сможет полностью контролировать жизненные процессы в любых организмах на всех уровнях.
Список литературы
1. Албертс Б., Брей Д., Льюис Дж. и др. Молекулярная биология клетки. Т. 1 - 3. М.: Мир, 1994.
2. Анализ генома. Методы / Под ред. К. Дейвиса. М.: Мир, 1990. 246 с.
3. Атанасов А. Биотехнология в растениеводстве. Новосибирск: ИЦиГСО РАН, 1993. – 241 с.
4. Барановов В.С. Генная терапия – медицина XXI века // Соросовский образовательный журнал. № 3. 1999. С. 3 – 68.
5. Бекер М.Е., Лиепиньш Г.К., Райпулис Е.П. Биотехнология. М.: Агропромиздат, 1990. 334 с.
6. Борисюк Н.В. Молекулярно - генетическая конституция соматических гибридов // Биотехнология. Итоги науки и техники ВИНИТИ АН СССР. М., 1988. Т. 9. С. 73 -113.
7. Валиханова Г.Ж. Биотехнология растений. Алматы: Конжык, 1996. 272 с.
8. Глеба Ю.Ю. Биотехнология растений // Соросовский образовательный журнал. № 6. 1998. С. 3 – 8.
9. Глебов О.К. Генетическая трансформация соматических клеток // Методы культивирования клеток. Л.: Наука, 1988.
10. Гольдман И.Л., Разин С.В., Эрнст Л.К., Кадулин С.Г., Гращук М.А. Молекулярно-биологические аспекты проблемы позиционно-независимой экспрессии чужеродных генов в клетках трансгенных животных // Биотехнология. 1994. № 2.
11. Дыбан А.П., Городецкий С.И. Интродукция в геном млекопитающих чужеродных генов: пути и перспективы // Молекулярные и клеточные аспекты биотехнологии. Л.: Наука, 1986. С. 82 - 97.
12. Егоров Н.С., Самуилов В.Д. Современные методы создания промышленных штаммов микроорганизмов // Биотехнология. Кн. 2. М.: Высшая школа, 1988. 208 с.
13. Зверева С.Д., Романов Г.А. Репортерные гены для генетической инженерии растений: хара¬ктеристика и методы тестирования // Физиология растений. 2000. Т. 47, № 3. С. 479-488.
14. Лещинская И.Б. Генетическая инженерия // Соросовский образовательный журнал. 1996. №1. С. 33 - 39.
15. Ли А., Тинланд Б. Интеграция т-ДНК в геном растений: прототип и реальность // Физиология растений. 2000, том 47, № 3. С. 354-359
16. Лутова Л.А., Проворов Н.А., Тиходеев О.Н. и др. Генетика развития растений. СПб.: Наука, 200. 539 с.
17. Льюин Б. Гены. М.: Мир, 1987. 544 с.
18. Пирузян Э.С., Андрианов В.М. Плазмиды агробактерий и генная инженерия растений.М.: Наука, 1985. 280 с.
19. Пирузян Э.С. Генетическая инженерия растений.М.: Знание, 1988. 64 с.
20. Пирузян Э.С. Основы генетической инженерии растений.М.: Наука, 1988. 304 с.
21. Пирузян Э.С. Проблемы экспрессии чужеродных генов в растениях // Итоги науки и техники ВИНИТИ. Сер. Биотехнология. 1990. Т. 23. 176 с.
22. Попов Л.С., Языков А.А. Трансгенные животные как модели для изучения репродукции эмбрионального развития и заболеваний человека // Успехи современной биологии.1999. Т 119, № 1. С. 30-41.
23. Романов Г.А. Генетическая инженерия растении и пути решения проблемы биобезопасности // Физиология растений, 2000. Том 47, № 3. С. 343-353
24. Сельскохозяйственная биотехнология: Учеб. / В.С. Шевелуха, Е.А. Калашникова, С.В. Дегтярев и др.: Под ред. В.С. Шевелухи. М.: Высш. школа, 1998. 416 с.
25. Сингер М., Берг П. Гены и геномы. Т. 1-2. М.: Мир, 1998.
26. Томилин Н.В., Глебов О.К. Генетическая трансформация клеток млекопитающих // Молекулярные и клеточные аспекты биотехнологии. Л.: Наука, 1986. С. 62 - 82.
27. Фаворова О.О. Лечение генами – фантастика или реальность? // Соросовский образовательный журнал. № 2. 1997. С. 21 – 27.
28. Щелкунов С.Н. Генетическая инженерия. Ч. 1. Новосибирск: Изд-во Новосибирского ун-та, 1994. 304 с.