| Скачать .docx | Скачать .pdf |
Реферат: Психрофільні мікроорганізми та їх використання
Реферат на тему:
«Психроф і льн і мікроорганізми та їх використання »
Вступ
В даному рефераті мова піде про психрофільні мікроорганізми, що широко представлені в тваринному світі. До них належать представники про – та еукаріотів, а також багато видів грибів, дріжджів та навіть комах і хребетних. В останні роки вплив даних мікроорганізмів широко вивчається оскільки область їх застосування досить різноманітна, від харчової промисловості до генної інженерії. Дослідженням даного питання займаються по всьому світу: в Індії, Росії, Японії, США, Канаді, Норвегії, Україні та інших країнах. Це пов’язано не тільки з науковою цікавістю, а насамперед, через можливість використання психрофільних організмів в умовах холодного, та помірного клімату, де температура опускається нижче 0 °С. Останні дослідження у галузі генетики виявили амінокислотні послідовності білків, що знаходяться і являються структурною складовою психрофілів та їх ферментів, що дозволяє проаналізувавши дану інформацію, виявити які конкретні гени відповідають за стійкість до досить низьких температур. Це в свою чергу призводить до використання таких особливостей в багатьох різних сферах, зокрема і в сільському господарстві, яке кожний рік зазнає втрат через морози. Також можливе використання можуть набути антибіотики виділені з психрофілів, так як вони ще не використовувались людьми, оскільки патогенні мікроорганізми не мали можливості пристосуватися оскільки вони раніше не були відомі, і виділяли їх вкрай рідкі види що зустрічаються тільки в Антарктиці.
Не дивлячись на важливість психрофільних організмів, вони вивчені вкрай недостатньо, оскільки сповільнене зростання робить їх незручними лабораторними об'єктами. Саме тому у даній роботі я намагатимусь дати уявлення про те якими особливостями вони володіють, і як ці особливості впливають на їх подальше застосування у сферах науки та техніки.
1. Основні чинники довкілля, що впливають на мікроорганізми. Вплив гідростатичного тиску
Вплив довкілля на живі організми включає ряд чинників, які обумовлюють фенотипічні особливості кожного виду і мікроорганізмів зокрема. До таких чинників відносяться:
· Гідростатичний тиск
· Сонячна активність
· Значення рН
· Кількість кисню
· Температура
1.1 Вплив гідростатичного тиску
Бактерії відносно мало чутливі до зміни гідростатичного тиску. Підвищення тиску до деякої межі не позначається на швидкості росту звичайних наземних бактерій, але врешті-решт починає перешкоджати нормальному зростанню і діленню. Життєдіяльність деяких бактерій пригнічується вже при 100 атм, але підвищення тиску до 200 атм навіть стимулює зростання бактерій таких як Е. coli. Слід зазначити, що різні процеси клітинного метаболізму різною мірою чутливі до підвищення тиску. Проте при тиску в 400 атм Е.coli починає утворювати нитчасті клітини, зростання яких сповільнене. При тиску в 1000 атм за 48 ч гинуло 90% клітин, а через 5 – 7 діб спостерігали їх повну загибель.
Барофільні бактерії розвиваються краще при тиску вищому, ніж тиск на земній поверхні, причому оптимальні значення тиску для різних штамів неоднакові. Зниження тиску наводить до уповільнення зростання таких штамів і порушення процесів ділення. При цьому утворюються ниткоподібні клітини.
Баротолерантні та барофільні бактерії зазвичай психрофільні, оскільки температура в глибинах океану низька. Підвищення тиску, так само як пониження температури, знижує текучість ліпідного біошару мембран. У глибоководних бактерій збереження оптимальної текучості мембран забезпечується своєрідним жирнокислотним складом їх ліпідів. Ці ліпіди характеризуються значним вмістом (до 20%) довголанцюгових поліненасичених жирних кислот, причому при підвищенні тиску вміст цих кислот зростає.
Поліенові кислоти мають відносно низьку температуру плавлення і відповідно не замерзають при значному підвищенні тиску. У складі мембран більшості вивчених бактерій подібні жирні кислоти не зустрічаються. При тиску, який значно нижче атмосферного, життєздатність бактерій зазвичай не порушується, проте при падінні тиску неминуче зменшується вміст в середовищі О2 , Н2 , С02 і інших газоподібних речовин, що може значно впливати на зростання бактерій.
В умовах глибокого вакууму субстрат висихає і життя неможливе[1].
1.2 Вплив сонячної активності і освітлення
Істотним чинником, що впливає на фізичні умови проживання організмів на Землі, є сонячна активність. Її зміна впливає на інтенсивність електромагнітного випромінювання Сонця. Короткохвильове випромінювання майже повністю поглинається у верхніх шарах атмосфери, впливаючи на стан іоносфери Землі і на фонові електромагнітні поля на поверхні Землі. Встановлений непрямий вплив сонячної активності на взаємодію патогенних мікроорганізмів з їх господарями. Наприклад, прослідивши зв'язок між проходженням сонячних плям через центральний меридіан Сонця, посиленням південно-західних вітрів і частотою виявлення збудника чуми в Ілі-Карательському межиріччі виявлено, що викликані змінами сонячної активності північно-західні вітри приносять значне пониження температури повітря; це, у свою чергу, призводить до посилення процесів обміну речовин в організмі звірів і підвищує їх сприйнятливість до збудника чуми, поширеність якого відповідно збільшується. [1]
Сонце впливає не лише побічно, змінюючи клімат, але і прямо. Під дією потоків заряджених часток, що викидаються з Сонця під час сонячних бурь, змінюється магнітне поле Землі. Його зміна впливає безпосередньо на клітини рослин. Проникність клітинних мембран збільшується, а ефективність змінних процесів із зовнішнім середовищем зростає. Значить, в цей час є можливість інтенсивніше «забирати» потрібні живильні речовини. Залежно від сонячної активності змінюється і чисельність різних мікроорганізмів, таких як амоніфікуючі і нітрифікуючі бактерії. Інакше кажучи, сонячна активність сама «здобрює» грунт. [2]
1.3 Вплив значення рН
Кислотність середовища є важливим чинником, що визначає існування в ній прокаріотів. Концентрація іонів водню в довкіллі діє на організм безпосередньо або побічно, через вплив на іонний стан і доступність багатьох іонів і метаболітів. Так, наприклад, при низьких значеннях рН знижується розчинність вуглекислоти – джерела вуглецю для автотрофних прокаріот, а розчинність таких катіонів, як Cu2 +, Mo2 +, Mg2 +, Ar+, зростає і досягає токсичних рівнів. Навпаки, при високих значеннях рН розчинність багатьох катіонів, необхідних клітині (Fe2 +, Са2 +, Mg2 +, Mn2 +), різко знижується, і вони стають недоступними для організму. Від значення рН залежить стан речовин в довкіллі. Багато органічних кислот в кислому середовищі знаходяться в недисоційованій формі і легко проникають в клітини, стаючи токсичними для неї.
Граничні розміри значень рН, оптимальні для зростання різних представників прокаріот, знаходяться в межах від 1 до 11. Залежно від відношення до кислотності середовища прокаріоти можуть бути розділені на декілька груп. Для зростання переважної більшості прокаріот оптимальним є середовище, близьке до нейтральної. Такі організми називають нейтрофілами. Проте зростання багатьох нейтрофілів можливе в середовищах, значення рН яких лежить в діапазоні від 4 до 9. Типовими нейтрофілами є штами Е.coli, Стsubtilis , Streptococcus faecalis . Багато нейтрофілів здатні зростати або виживати при значеннях рН, що лежать за межами вказаного діапазону. Такі прокаріоти вважаються кислото – або лужно-толерантними. До кислототривких відносяться багато бактерій, що продукують органічні кислоти, наприклад оцтовокислі, молочнокислі та ін. Лужнотолерантні, тобто стійкі до значень рН близьким до 9–10, багато з ентеробактерій.
У деяких видів бактерій адаптація до певних значень рН середовища призвела до того, що оптимальним рН для зростання перемістився в кислу (рН 4 і нижче) або лужну (рН від 9 і вище) області. Такі прокаріоти названі ацидо – або алкалофільними (кислото – або луго любивими) відповідно.
Здібність до зростання при низьких або високих значеннях рН забезпечує організму певні переваги, оскільки в цих умовах мала конкуренція з боку більшості інших організмів. Проте деякі бактерії – облігатні форми – не просто переносять високі концентрації Н+ або ОН-, але і потребують цих іонів для зростання і стабільності[1].
1.4 Вплив кількості кисню
Кисень широко поширений в природі, знаходячись як в зв'язаному, так і вільному стані. У першому випадку він входить до складу молекул води, органічних і неорганічних сполук. У другому – присутній в атмосфері у вигляді молекулярного кисню (О2 ), об'ємна доля якого складає 21%.
Кисень є обов'язковим хімічним компонентом будь-якої клітини. Переважна більшість організмів задовольняють свої потреби в цьому елементі, використовує обидві форми кисню. При вирощуванні Pseudomonas у присутності 18О2 і 18Н2О джерелом приблизно 10% кисню, що входить до складу клітинного матеріалу, служив газоподібний кисень, 50–60% клітинного кисню походило з води. Останній кисень в клітини поставляли органічні і неорганічні компоненти живильного середовища (глюкоза, фосфати, нітрати, сульфати і ін.).
Серед прокаріотів існують значні відмінності у відношенні до молекулярного кисню. За цією ознакою вони можуть бути розділені на декілька груп (рис. 1).
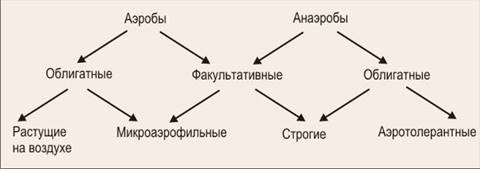
Рис. 1. Класифікація за споживанням кисню
Прокаріоти, для зростання яких О2 необхідний, називають облігатними аеробами. До них відносяться більшість прокаріотних організмів. Серед облігатних аеробів виявлені істотні відмінності у відношенні до рівня молекулярного кисню в середовищі. Деякі представники цієї групи не здатні до зростання при концентрації О2 , рівній атмосферній, але можуть зростати, якщо вміст О2 в довкіллі буде значно нижчий (приблизно 2%). Такі облігатні й аеробні прокаріоти отримали назву мікроаерофілів.
Потреба прокаріотів у низькій концентрації О2 в довкіллі пов'язана з їх метаболічними особливостями. Багато аеробних азотфіксуючих бактерій можуть зростати в середовищі з молекулярним азотом лише при концентрації О2 нижче 2%, тобто як мікроаерофіли, а у присутності зв'язаного азоту, наприклад амонійного, – на повітрі. Це пояснюється дією молекулярного кисню, що інгібує, на активність нітрогенази – ферментного комплексу, відповідального за фіксацію N2.
Хоча облігатні анаеробні бактерії в цілому дуже чутливі до О2 , вони можуть в природі знаходитися в аеробних зонах. Широке поширення представників роду Clostridium в місцях з високим парціальним тиском О2 пояснюється наявністю у них эндоспор, не чутливих до молекулярного кисню. Проте строго анаеробні прокаріоти виявлені в природі в місцях, де спостерігається активний розвиток облігатних аеробів. Ймовірно, спільний розвиток з облігатними аеробами, активно споживаючими молекулярний кисень, що призводить до утворення зон з низькою концентрацією О2 , створює можливості і для розвитку строго анаеробних видів.
Існують прокариотні організми, які можуть зростати як в аеробних, так і в анаеробних умовах. Вивчення цього явища показало, що природа його різна. Бактерії, що не потребують О2 (останній не бере участь в здійснюваних ними метаболічних реакціях), але здатні зростати в його присутності, є за типом здійснюваного ними метаболізму облігатними анаеробами, стійкими до О2 зовнішнього середовища. Прикладом таких організмів служать молочнокислі бактерії. Багато прокаріотів, що відносяться до цієї ж групи, пристосувалися залежно від наявності або відсутності О2 в середовищі перемикатися з одного метаболічного шляху на інший, наприклад з дихання на бродіння, і навпаки. Такі організми отримали назву факультативних анаеробів, або факультативних аеробів. Представниками цієї фізіологічної групи прокариот є ентеробактерії. У аеробних умовах вони отримують енергію в процесі дихання. У анаеробних умовах джерелом енергії для них служать процеси бродіння або анаеробного дихання.
Потреба в О2 в аеробів визначається його участю в енергетичних і конструктивних процесах. У першому випадку О2 служить обов'язковим кінцевим акцептором електронів, в другому – бере участь в реакціях на шляху багатоступінчастого перетворення клітинних метаболітів або екзогенних субстратів. В облігатних аеробів велика частина О2 використовується як кінцевий акцептор електронів в реакціях, цитохромоксидазами, що каталізують. Менша частина включається в молекули за допомогою ферментів, що отримали загальну назву оксигеназ. У клітинах факультативних анаеробів також містяться цитохромоксидази. В облігатних анаеробів немає ферментів, що каталізують взаємодію з О2 . [3]
1.5 Вплив температури
Всі фізико-хімічні процеси, що забезпечують функціональну активність клітини, а також стан її макромолекул, більшою чи меншою мірою залежать від температури.
З підвищенням температури швидкість хімічних реакцій зростає, отже, швидкість росту мікроорганізму збільшується. Проте при високих значеннях температури білки, нуклеїнові кислоти і інші компоненти клітин можуть безповоротно інактивуватися, що призводить до її загибелі. При дуже низькій температурі також порушуються процеси біосинтезу, і зростання припиняється.
Для кожного організму існує мінімальна температура, нижче за яку зростання не спостерігається, як би довго не тривав період інкубації. Оптимальна температура, при якій організм зростає з найбільшою швидкістю, і максимальна температура, вище за яку зростання неможливе. Оптимальна температура завжди ближче до максимальної, ніж до мінімальної. Ці три температурні позначки, названі кардинальними, характерні для кожного виду або навіть штаму.
Значення кардинальних температур можуть декілька змінюватися під впливом різних чинників середовища, наприклад, при вирощуванні мікроорганізмів на різних живильних середовищах. В більшості випадків зростання бактерії відбувається в температурному інтервалі порядку 40 °С, але інколи ці рамки значно вужчі. Температурні інтервали, в яких можливе зростання бактерій в природі, широкі – від негативних до значень вище 100 °С. Значення кардинальних температур у різних бактерій вельми істотно розрізняються. По відношенню до температури бактерії ділять на мезофіли, термофіли і психрофіли.
Мезофіли достатньо поширені в природі, їх можна виявити в грунті, у воді помірних і тропічних широт, в організмі різних тварин і людини. Діапазон температур, в якому можливе зростання тих або інших форм мезофільних бактерій, різний, так само як всілякі значення їх кардинальних температур.
До термофільних відносять організми, які зростають при температурі вище 45–50 °С. Залежно від значень кардинальних температур серед термофільних бактерій розрізняють: облігатні термофіли, що мають температурний оптимум 65–70 °С і мінімум вище 40–42 °С, факультативні термофіли з температурним максимумом 50–65 і мінімумом менше 20 °С і термотолерантні бактерії з температурним максимумом 45–50 °С. Бактерії з температурним оптимумом вище 70 °С визначають як екстремально термофільні. Термотолерантні бактерії здатні переносити досить високу температуру без порушення нормального зростання. Вони відрізняються від термофільних, що віддають перевагу високим значенням температури, по характеру зміни швидкості росту при підвищенні температури культивування. [1] Психрофіли ж вимагають окремого розгляду.
2. Характеристика психрофілів
Область температур розмноження психрофілів лежить в межах від -10 до +20о С і вище. У свою чергу, психрофіли поділяються на облігатні та факультативні.
Основна відмінність між підгрупами полягає в тому, що облігатні психрофіли не здатні до зростання при температурі вище 20о С, а верхня температурна межа зростання факультативних форм набагато вища. Таким чином, факультативні психрофіли характеризуються ширшим температурним діапазоном, при якому можливе їх зростання. Якщо в області низьких температур вони схожі з облігатними формами, то в області підвищених температур володіють здатністю розмножуватися в значно вищих температурних межах. Розрізняються вони також і оптимальними температурними зонами зростання, що знаходяться в облігатних психрофілів значно нижче, ніж у факультативних.
Існування двох типів психрофілів пояснюється особливостями їх середовища існування. Облігатні психрофіли пристосувалися до стійких холодних умов (глибини морів та океанів, крижані печери). Психрофіли іншого типу пристосувалися до існування в нестійких холодних умовах. У природі більшість психрофілів представлена факультативними формами. Здатність психрофілів зростати в умовах низьких температур пов'язують в першу чергу з особливостями їх ферментних білків і мембранних ліпідів. Збільшення в останніх вмісту ненасичених жирних кислот дозволяє мембранам знаходитися у функціонально активному рідинно-кристалічному стані при низьких температурах. Обов'язкова умова можливості зростання психрофілів при мінусових температурах – знаходження води в рідкому стані.
Зростати при низькій температурі можуть також організми, що одержали назву психроактивних («психротрофних»). По суті це евритермні організми, що мають достатню активність при низькій температурі. Екофізіологічна відмінність між психрофілами і психроактивними організмами досить велика. Психрофіли існують в постійно холодних умовах, наприклад, в глибинах океану або глибоких водоймищах, де коливання температури дуже незначні. Психроактівні пристосовані до сезонних змін клімату, в теплий період вони накопичують біомасу, але продовжують зростати і в той час, коли активність інших пригнічена. Пристосування до пониження температури зв’язують із зміною складу мембран і із здатністю до створення криопротекторів. При знижених температурах знижується не лише швидкість росту, але і швидкість відмирання, і, відповідно, збільшується виживання. Інший механізм пов'язаний з підвищеним синтезом ключових ферментів що мають вищий температурний оптимум, але лише завдячуючи накопиченню тих, що зберігають достатню для функціонування клітин активність. Серед психрофілів багато протистів і багатоклітинних [4].
Психрофілія не є властивістю якийсь конкретною систематичної групи. Важливими первинними продуцентами в таких умовах, мабуть, служать еукаритичні водорості. Так «снігова» зелена водорость Chamydomonas nivalis , вегетативні клітини якої забарвлені в зелений колір, а спори в червоний колір утворює рожеві нальоти на поверхні снігу і льоду. У покритих льодом водах Антарктики виявлено діатомові і зелені водорості.
Багато психрофілів є грамнегативними бактеріями і їх представники виявлені серед родів Vibriо, Рseudomonas Асhrоmоbасtеr, Flavobacterium, Cytophaga . Серед грамнегативних бактерій психрофіли описані у видів роду Clostridium .
Психрофіли широко поширені в природі, оскільки вони формують постійну мікрофлору регіонів вічного холоду, полярних регіонів і океанів, які займають14 і 71% поверхонь Землі відповідно.
В даний час психрофіли знайдені в південній частині Антарктиди і гірських льодовиках Арктики. Океани також є природним середовищем існування психрофільних бактерій, де вони знаходяться у воді у вільному стані або в симбіозі з морськими рослинами або тваринами
Аналіз проб морської води на 16S рРНК показав присутність у ній різних архей. По числу і різноманітності представників бактерії зазвичай домінують над археями, хоча на великих глибинах морів і океанів знайдені еквівалентні кількості архей, які відносять частіше до родів Меthanogenium і Меthanococcus [13].
Психротрофи також широко поширені в природі. Вони описані серед представників родів Васillus, Мiсrососсus Clostridium, Рseudomonas, Chrоmоbасtеr, Acinetobacter і ін. Багато механізмів адаптації до холоду вивчені в таких модельних психротрофних видів, як Васillus psychrophilus і Мiсrососсus сryophilus . Пристосування до зниженої температури виявляється у зміні складу мембран (у ній підвищується вміст ненасичених жирних кислот) і синтезі криопротекторів (наприклад глицерола). Інший механізм пов'язаний з накопиченням в клітках більших кількостей найбільш важливих ферментів, так що навіть при неоптимальній температурі їх функціонування дозволяє клітині підтримувати достатню активність [5].
У зоні холодного і помірного клімату значення психрофільних і психротрофних
мікроорганизмів в природних процесах дуже велике. При дослідженні метаногенних колоній мікроорганізмів тундри групою російських вчених під керівництвом Г.А. Заварзіна вперше було виявлено перемикання трофічного маршруту асоціацій мікроорганизмів в залежності від температури, обумовлене зміною домінуючих груп мікроорганизмів. Так, при температурі вище 15 °С основним кінечним процесом в колоніях мікроорганізмів був метаногенез, а нижче 15 °С – створення ацетату [6].
Присихрофільними властивостями характеризуються не тільки бактерії а й дріжджі (наприклад: Candida, Criptococus, Rhodotorula ), гриби (наприклад: Aureobasidium, Dematium, Cladosporium, Hormodendron ) та бактеріофаги (наприклад: фаги для Pseudomonas та Micrococus cryophilus ) [12]. Згідно з літературними даними велика кількість психрофілів виділено з різних об'єктів по всьому світу.
Дж. Боуменом із співавторами описані різноманітність і асоціації психрофільних мікроорганізмів льодів Антарктичного морів [7]. З прибережної частини Охотського моря виділено факультативний психрофіл Psychrobacter okhotskensis, що продукує ліпазу і нова галофільна факультативна психрофільна бактерія Psychromonas marina [8].
Підчас 7-ої антарктичної української експедиції було виділено бактерію Methylobacterium що знижує температуру кристалізації води до -13 °С [9].
Результати отриманих зразків ґрунту в Гімалаях (Індія) показали що більш ніж 60% мікроорганізмів що знаходилися у зразках були психрофілами[10].
А також проведені досліди з бактерією штамму Colwellia 34H, показали можливість існування бактерій які проявляють активність навіть при температурі -193,5 °С [11].
2.1 Вплив температури на ріст та розмноження
Ріст – основна функція мікроорганізмів, на яку впливає температура. Ріст підпорядковується термодинамічним законам, хоча має і свої специфічні особливості.
Основною характеристикою темпу розмноження мікроорганізмів є питома швидкість росту. Середня питома швидкість росту за проміжок часу (t1 -t2 ) розраховується за формулою 1:
![]() де: m1
таm0
– початкова і кінцева вага клітин.
де: m1
таm0
– початкова і кінцева вага клітин.
Формула 1 Швидкість росту
Температура, при якій швидкість росту максимальна називається оптимальною.
Головною особливістю психрофілів, на відміну від інших мікроорганізмів, є здатність інтенсивно рости при температурах 0–10 °С. Прикладом можуть послугувати такі вибіркові данні:
Табл. 1. Час генерації різних штамів Pseudomonas при температурах від 26–0 °С
Температура в °С |
Час генерації в хв. | |
Pseudomonas 1–36 (психрофіл) |
Pseudomonas aeruginosa (мезофіл) | |
| 26 | 59 | - |
| 25 | - | 56 |
| 20 | 95 | 120 |
| 15 | - | 160 |
| 14 | 140 | - |
| 10 | 160 | 470 |
| 8 | 210 | 1400 |
| 0 | 620 | - |
Можна також привести данні про інтенсивність росту у факультативних та облігатних психрофілів, при температурах які прямують до граничних для їх росту. Таким приладом є представлений графік Арреніуса граф. 1.

Графік 1. Крива Арреніуса для швидкості росту психрофілів
Графік дає уявлення, що швидкість росту для облігатного психрофілу залишається лінійною до -2 °С, а для факультативного відхилення починається за декілька градусів від нуля. Це вкрай важливе спостереження, оскільки воно може являтися фактором, що приводить до екологічного домінування в місце існуваннях з низькою температурою. Ці данні були підтвердженні в експериментах з морськими психрофілами в модельних системах, а також в умовах безперервного культивування. В цих дослідах облігатні психрофіли виявилися більш конкурентно спроможними, при температурах нижче 15 °С.
Іншим показником є температурний коефіцієнт росту (Q10 ) він визначається як відношення швидкостей при одній температурі та при іншій на 10 °С нижчій.
Q10 =Kt-/Kt
Формула 2. Температурний коефіцієнт росту
Для функції росту коефіцієнт може бути визначений в будь-якому інтервалі температур ΔТ з допомогою наступної формули:
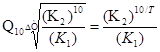
Формула 3. Температурний коефіцієнт Q10 для функції росту при температурі ΔТ
К1 та К2 –температурні коефіцієнту росту з різницею в 10 °С
Залежність швидкості біологічних реакцій від температури робить цей показник досить зручним чинником для порівняння мікроорганізмів по ступеню стійкості швидкості росту до зниження температури[12].
2.2 Фактори, що обумовлюють низьку максимальну температуру росту
Вважається, що верхня температурна межа росту мікроорганізмів залежить від термолабільних одного або кількох важливих у хімічному або структурному відношенні компонентів. Очевидно, якщо клітинний компонент (або будь-яка активність) відрізняється чутливістю до температури, близької до максимальної температури зростання, то між цією чутливістю і верхньою межею зростання повинен існувати причинний взаємозв'язок. Крім того, іноді вдається простежити позитивний взаємозв'язок між зменшенням термолабільності одного або декількох клітинних компонентів (або активності) і збільшенням максимальної температури зростання мікроорганізмів. Деякі дослідники культивували мікроорганізми при температурах як вище, так і нижче дійсної максимальної температури їх зростання. Дані, отримані в результаті таких експериментів, сприяють кращому розумінню механізмів, які визначають верхню межу зростання психрофільних і психротрофних мікроорганізмів. Синтез білків інтактними клітинами деяких психрофільних мікроорганізмів при відносно помірних температурах припиняється. Наприклад, синтез білків облігатним психрофілом Pseudomonas зупиняється при 22,5 ° С. Аналогічно в умовах безперервного культивування в хемостаті швидкість синтезу білків цим же організмом швидко падала при температурі близько 20 ° С. У цих умовах переставав синтезувати білки і одночасно втрачав життєздатність А4 cryophilus .
Слід зазначити, що температури, при яких відбувається денатурація рибосом психрофільних мікроорганізмів, набагато більша, ніж верхня межа зростання цих організмів. Разом з тим існує позитивна кореляція між температурою денатурації рибосом або рРНК і максимальною температурою зростання різних психрофілів. Таким чином, важко сказати чи має такий взаємозв'язок пряме відношення до верхньої температурної межі росту психрофільних мікроорганізмів. Характер фізичних змін рибосом під дією нагрівання не завжди знаходиться в прямій залежності з максимальною температурою зростання психрофільних мікроорганізмів. На відміну від дії підвищених температур на структуру рибосом на їх функціонування у психрофільних мікроорганізмів впливають температури набагато більш близькі до температур верхньої межі зростання. Рибосоми психрофілів можуть втратити активність при температурах, які мало відрізняються від максимальної температури росту мікроорганізму, хоча зростання при цьому ще можливе.
Вплив відносно невисоких температур може призводити до модифікації структурних компонентів мікроорганізмів, що ростуть при низьких температурах. Іноді відбуваються істотні зміни тонкої структури мікроорганізмів. В інших випадках спостерігаються фізіологічні порушення, викликані структурними змінами. Температура може також викликати ультраструктурні зміни у В.psychrophilus . Клітинні стінки цього психрофіла швидко руйнувалися при 40 ° С, що супроводжувалося злипання клітин та їх загибеллю. Модифікація структури клітин психрофілів під дією температури може позначатися і на їх зовнішньому вигляді. Крім безпосередньо спостережених або вимірюваних змін клітинної структури при дії відносно помірних температур з виділення внутрішньоклітинних речовин або справжньому повного лізису можна виявити модифікацію таких структур, як структури, відповідальні за проникність. Облігатний психрофіл Candida nivalis виділяє в навколишнє середовище різні внутрішньоклітинні розчинні сполуки: амінокислоти або короткі поліпептиди, неорганічний фосфат і нуклеотідмонофосфати. Виділення цих сполук не може бути викликано лізисом клітин, так як загальна кількість клітин, визначена прямим рахунком або за мутності, при цьому не зменшується. Далі, оскільки виділення з клітин білка і нуклеїнових кислот не виявлено, можна припускати, що клітинна мембрана піддається специфічній модифікації. Виділення названих вище сполук корелює із втратою клітинами життєздатності при 25 ° С; не виключено, що низька максимальна температура зростання цього мікроорганізму принаймні частково пояснюється термолабільністю клітинної мембрани.
Під дією відносно помірних температур може відбуватися лізис клітин психрофілів, а не тільки виділення з них з'єднань. Так, наприклад, цілі клітини В.psychrophilus зазнають лізис в калій-фосфатному буфері, рН 6,5 при температурах вище 28 °С. Чутливість ферментів деяких психрофільних мікроорганізмів до відносно помірних температур може бути причиною (чи однією з причин) того, що ці організми відрізняються низькою максимальною температурою росту. Зважаючи на це пояснення, слід, однак, враховувати ту обставину, що фермент може інактивувати під дією температури тільки у виділеному стані, але не в інтактних клітинах. Іноді цілісність клітини забезпечує повний або частковий захист активності термолабільних ферментів. Наприклад, малатдегідрогеназа в екстрактах клітин облігатного психрофіла V.marinus MP-1 легко інактивується при нагріванні. Активність цього ферменту знижувалася навіть при 0 ° С і майже повністю зникала в результаті інкубації протягом 10 хв при 30 ° С, тобто при температурі, що перевищує максимальну температуру росту. Аналогічну інактивацію зазнавав очищений в 20 разів препарат цього ферменту. Разом з тим, при температурах нижче оптимальної температури росту активність малатдегідрогенази в інтактних клітинах не змінювалася; отже, в цих температурних межах її захист забезпечується цілісністю клітин. Проте при температурах вищих оптимальної (15 °С) активність цієї дегідрогенази значно знижується навіть у інтактних клітинах даного психрофільного організму. Таким чином, чутливість цього ферменту до відносно низьких температур може служити причиною низького верхньої температурної межі зростання бактерій.
У інших психрофільних мікроорганізмів термолабільні ферменти виявляються зовсім не захищеними навіть у інтактних клітинах. Швидкості окислення глюкози і ендогенних субстратів зруйнованими клітинами В.psychrophilus змінювалися при зростанні температури від 5 до 45 ° С так само, як і в інтактних клітинах. Крім того, як показало порівняльне дослідження безклітинних препаратів інтактних клітин цього психрофільного організму і мезофільні штаму Bacillus , у психрофіла активність окислення глюкози пригнічувалась більшою мірою при низькій температурі.
Психрофільні гриби також містять термолабільні ферменти, від яких може залежати верхня межа зростання цих організмів. Швидкість ендогенного метаболізму сніжної цвілі Typhula idahoensis знижується при 20 °С – температурі, що пригнічує зростання.
У деяких психрофільних мікроорганізмів загальна активність і зростання пригнічується при порівняно низьких температурах через наявність тільки одного термолабільного ферменту[13].
3. Використання психрофільних мікроорганізмів в промисловості
Новітні досягнення в області науки і техніки дозволили використовувати, особливості психрофільних мікроорганізмів. Спектр такого використання досить широкий, від отримання специфічних термолабільних ферментів, до проектування водо – та грунтоочисних споруд. Хоча потенціал використання психрофільних організмів досить значний, не дивлячись на їх температурні особливості, що ускладнюють процес їх культивування і розмноження, психрофільні мікроорганізми призводять до багатьох проблем. Прикладом таких негативних впливів є прояв редуцентних властивостей в харчовій промисловості, та підгнивання пшениці в сільськогосподарських угіддях. Таким чином, в представленій роботі буде більш детально розглянуто промислові особливості і можливості даних організмів.
3.1 Засоби для охорони навколишнього середовища
Серед основних чинників очищення морської води та ґрунту, важливу роль відіграє біологічний фактор. Для прискорення процесів очищення морських акваторій та ґрунту необхідне застосування технологій, основаних на останніх науково-технічних розробках, що забезпечують результат при відносно невеликих фінансових витратах.
Застосування біологічних препаратів прискорює процес деструкції нафти в природних умовах за допомогою нанесення на поверхню забруднених земель культур мікроорганізмів, здатних розщеплювати вуглеводні. Розроблений препарат «Біоойл-СН» застосовується у виробничих масштабах. Препарат складається з асоціації мікроорганізмів на основі мезо- та псіхрофільних штамів – деструкторів нафти, що працюють при температурі від 1 °С в широкому діапазоні кислотності середовища по деструкції різноманітних фракцій нафти в ґрунтовому і водному профілях. Всі 6 штамів, що входять до складу препарату, відносяться до родів Saccharomyces sp, Enterobacter sp., Bacillus sp. (2 види), Acinetobacter sp. (2 види) депоновані, є висновок про їх безпеку для людини і навколишнього середовища. Консорціум псіхрофільних бактерій: Acinetobacter calcoaceticus Baumann et al. штам 279 МКПМ-В-7179, Psеudomonas fluorescens Stanier et al . штам 325 МКПМ-В-7152, Alcaligenes faecalis Castellani et Chalmers штам 404 МКПМ-В-7180-призначений для очищення грунту, води, ємностей і стічних вод промислових підприємств від забруднень нафтопродуктами в північних регіонах країни за 3 – 15 °C. Селекціоновані псіхрофільні штами бактерій характеризуються високою окисної здатністю, стійкістю в тривалому безперервному та періодичному процесі, активно ростуть в прісній і морській воді. Винахід відноситься до засобів боротьби із забрудненнями грунту, води і ємностей нафтою і нафтопродуктами за допомогою спеціально підібраних асоціації псіхрофільних культур бактерій. Процес самоочищення в природних умовах Півночі, Західного Сибіру, районах, де недовгий літній період, що пов'язано з бідністю флори нафтоокислюючих бактерій, дефіцитом органіки, аерації, низькою температурою середовища, що значно ускладнює використання біологічних агентів[14].
Для очищення стічних вод від важких металів та радіонуклідів, були розроблені біологічні препарати МБК та ЗМУ. Діючою основою препаратів є гранули, що складаються із живих мікроорганізмів і необхідних для них джерел живлення. До складу препаратів входять мікробні асоціації з широким «метаболічним діапазоном». Внесення в препарати різноманітних субстратів дозволяє регулювати метаболізм мікробних асоціацій і досягати максимального ефекту очищення стічних вод від ксенобіотиків. Видове різноманіття мікроорганізмів та їх висока концентрація забезпечують одночасну реалізацію декількох незалежних механізмів детоксикації ксенобіотиків. Тому препарати МБК та ЗМУ із високою швидкістю та ефективністю очищують змішані стоки, що містять суміш важких металів, радіонуклідів та органічних сполук. Препарати МБК та ЗМУ ефективно очищують розчини від металів, радіонуклідів та органічних забруднень. МБК вилучає на 98,5–99,9% важкі метали (Cu2+, Hg2+, Cd2+, Pb2+, Zn2+, CrО42– і т.д.) у діапазоні 0,006 – 10000 мг/л. МБК та ЗМУ знижують активність РРВ по сполукам 51Cr, 90Sr, 83Rb, 54Mn, 140La, 60Co – на 5–6 порядків; вилучають уран, плутоній і америцій на 99,6 – 99,9%. ЗМУ вилучає з дуже розведених розчинів рідкісноземельні та дорогоцінні метали та забезпечує глибоку доочистку стоків від зникаюче малих концентрацій важких металів та органічних забруднень. Очистка РРВ на прикладі Спецкомбінату «РАДОН». До складу РРВ Спецкомбінату «Радон» входять 4 радіонукліди з активністю (Бк/кг): 137Cs (10135,7), 60Co (30,7), 22Na (79,6), 90Sr (50192). Біотехнологія передбачала очистку РРВ в періодичних та проточних умовах в касетній установці, що представлена десятьма послідовно з‘єднаними циліндрами з гранульованим МБК. Схема даної установки зображена на рисунку 2.
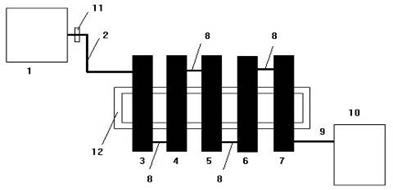
Рисунок 2. Схема проточної установки для очистки РРВ за допомогою МБК
1 – ємність з РРВ; 2 – шланг для подачі РРВ; 3–7 – колонки з МБК; 8 – шланги сполучення колонок; 9 – шланг для подачі очищених РРВ; 10 – ємність для очищених РРВ; 11 – вентиль подачі РРВ в установку; 12 – рама для закріплення колонок[15].
3.2 Застосування і проблеми у сільському господарстві
В останні роки значно зросло застосування біологічних препаратів у сучасному сільському господарстві. Одним з перспективних напрямків застосування мікробіологічних препаратів є їх використання при закладці плодоовочевої продукції на зимове зберігання. Застосування штучного холоду – незамінна умова для продовження термінів зберігання плодово-овочевої продукції. Однак, холодильне зберігання при температурах, близьких до кріоскопічної, не виключає ураження продукції псіхрофільними мікроорганізмами. В процесі пошуку альтернативних методів захисту рослин вагому перспективу відкривають біологічні засоби захисту, які зможуть успішно конкурувати з хімічними, не маючи в той же час їх недоліків: «звикання» збудника до фунгіцидів, токсичність, накопичення в екологічних харчових ланцюгах.
Особливо перспективним є використання бактерій-антагоністів фітопатогенів при зберіганні рослинної продукції, Також було встановлено, що бактерії-антагоністи і продукти їх життєдіяльності здатні активізувати захисні реакції бульб картоплі, такі як накопичення суберіна і процеси ранової репарації У результаті багаторічних досліджень був розроблений біопрепарат комплексної дії Екстрасол. Застосування Екстрасола для обробки продовольчої картоплі призначеного для закладання на зберігання, дозволяє значно знизити втрати, що виникають в результаті розвитку фітопатогенної мікрофлори.
А що в край важливо, так це те що були проведені досліди, в результаті яких отримані данні про те що, бактерії не впливають на організм людини оскільки вони інактивуються, при потраплянні до людського організму через свою термолабільність. [17]
3.2 Застосування у харчовій промисловості
Для виробництва м'ясних виробів одним із перспективних напрямків є використання біологічно активних речовин на основі продуктів життєдіяльності мікроорганізмів. Встановлено, що мікроорганізмі, внесені з заквасками, за рахунок ферментів змінюють структуру ковбас, створюючи нові речовини, що покращують якість показників продукту. Область застосування психрофільного, галотолерантного, денітріфікуючого штаму Paracoccus dentrificans була визначена як стартова культура для посолу свинячих та яловичих копченостей. Різниця в температурних оптимумах розвитку цих мікроорганізмів дає можливість штаму ефективно редуціювати нітрат натрію до активізації молочнокислих мікроорганізмів. Це є важливим у зв'язку з тим, що молочнокіслі мікроорганізми володіють вираженим антагонізмом не тільки по відношенню до штаму Paracoccus sp ., але і по відношенню до самого процесу денітріфікації за рахунок зниження рН. При цьому у сировині накопичується окис азоту, яка в умовах подальшого зниження рН забезпечує найбільш сприятливі умови для реакції кольоростворення.
Розробка Датського м'ясного інституту – стартова культура Moraxella penylpyruvica цікава з точки зору ароматостворення. Це психрофільна культура – факультативний анаероб, що дозволяє їй активно розвиватися в товщі продукту і як показали дослідження, продукувати попередника аромату. Американські технологи, поряд з традиційними бактеріями, такими як Lactobacillus та Pediococcus, в склад стартових культур включають Micrococcus, який володіє здатністю відновлювати нітрати та нітрити, при цьому покращують смак і колір готових ковбасних виробів.
Японські дослідники виділили з психрофільного бактеріального штаму, ізольованого з проб води в Антарктиці, термолабільну фосфатазу, а з галофільного штаму Pseudomonas sp. – протеазу, яка проявляє максимальну активність при 18% концентрації NaCl в середовищі. Цей фермент використовується для приготування рибного соусу.
4. Обладнання для культивування психрофілів
Обладнання яке потрібне для культивування психрофільних мікроорганізмів, значно не відрізняється від типових вже розроблених біореакторів, оскільки процес регулювання температури, не є складним з точки зору апаратурного забезпечення. Тому більшу увагу треба приділити таким аспектам як аерація та перемішування.
Однорідність перемішування повинна забезпечити відсутність градієнтів в середовищі по температурі, рН, концентрації субстрата та продукту. В той же час необхідно враховувати небезпеку травмування в процесі перемішування. Типовий спосіб перемішування здійснюється за рахунок застосування гвинтового перемішуючого пристрою з великим діаметром та невеликою швидкістю обертання. Пневматичне перемішування в аерліфтних реакторах або гідравлічне перемішування за допомогою зовнішнього насоса також дозволяє вирішити цю проблему при роботі з культурами клітин.
Для запобігання утворенню піни і зниження ризику травмування клітин бульбашками повітря використовують менший обсяг повітря, що подається, площі аерації або застосуванням системи подачі повітря «вільного від бульбашок». При скороченні обсягу повітря, що подається, необхідно збільшити об'ємну долю кисню в газовій суміші, що подається. Для цього застосовують системи змішування газів (повітря, азот, кисень, вуглекислий газ) для забезпечення оптимальних умов.
Культивування клітин може здійснюватися в режимах періодичному (batch), з підживленням (fed-batch) і безперервному. Періодичне культивування характеризується зростанням клітин без подачі додаткових порцій субстрату після посіву культури. Ліміт субстрату або утворення токсичних компонентів можуть призвести до зниження продуктивності процесу. Для запобігання негативним наслідкам ліміту субстрату застосовується техніка культивування з підживленням, при цьому субстрат або інші необхідні компоненти додаються або безперервно, або по сигналу від датчика. При інгібуванні зростання продуктами метаболізму (лактат, амоній) використовується безперервне культивування, де в ході процесу відбувається відбирання середовища з доливанням компенсуючого об'єму свіжого середовища. Оскільки низька швидкість росту є одним з лімітуючих чинників при культивуванні психрофілів безперервний процес необхідно вести із затриманням біомаси і недопущенням її віднесення з вихідними об'ємами культуральної рідини.
Висока щільність культури забезпечить високу швидкість процесу при безперервному культивуванні. Затримання клітин можливо здійснити за рахунок вживання адекватної системи фільтрації.
Багато клітин можуть зростати лише прикріпленими до поверхні. Такі культури вимагають прикріплення до мікроносіїв таких як: скло, целюлоза, декстрин, колаген, желатин або пластик. Якщо носій має пори, клітини можуть зростати всередині носія тим самим, знижуючи негативний вплив перемішування і повітряних бульбашок.
Висновки
Отже виходячи з вище зазначеного можна впевнитися, що психрофільні організми мають поширене використання. А також мають перспективи в застосуванні в місцях з холодним кліматом, або нестабільними погодними умовами, в генній інженерії, та біохімії. Процеси культивування психрофілів, менш матеріально та енерговитратні, оскільки не потребують підтримування високої температури. Також вони є альтернативним джерелом специфічних ферментів та білків, а також антибіотиків. Все це говорить про доцільність розглянутого питання. Роботу виконано згідно ДСТУ 3008–95.
Перелік використаних джерел літератури
1. «Экология бактерий» Б.В. Громов, Г.В. Павленко Санкт-Питербург 1989
2. «Возмутители спокойствия» Д. Назаров Москва 2001
3. http://medbiol.ru/medbiol/microbiol/000bfa27.htm
4. «Введние в природоведческую микробиологию» Г.А. Заврзин Н.Н. Колотилова Москва 2001
5. «Молекулярные аспекты адаптации прокариот» Е.В. Ермилова
Санкт-Питербург 2007
6. «Лекции по природоведческой биологии» Г.А. Заварзин Москва 2003
7. «Methods for Psychrophilic Bacteria» JP Bowman London 2001
8. «Psychrobacter okhotskensis, a lipase-producing facultative psychrophile isolated from the coast of the Okhotsk Sea» Isao Yumoto 2003
9. « PSYCHROPHILIC METHYLOTROPHIC BACTERIA IN ANTARCTIC REGION BIOTOP E»V.A. Romanovskaya, P.V. Rokitko O.B. Tashirev, S.O. Shilin, N.A. Chernaya Kiev 2006
10. «Bacterial diversiry of soil samples from the western of Himalayas»Pooja Ganwar, Syed Inteyaz Alam, Sunita Bansod, Lokendra Singh 2008
11. «Sub-zero survivors» Fox, Douglas 2006
12. «Адаптация микроорганизмов к низким температурам» С.П. Лях Москва 1976
13. «Cold adapted Archaea» Cavicchioli, R. 2006
14. «КОНСОРЦИУМ ШТАММОВ БАКТЕРИЙ ACINETOBACTER CALCOACETICUS , PSEUDOMONAS FLUORESCENS , ALCALIGENES FAECALIS ДЛЯ ДЕСТРУКЦИИ НЕФТИ И НЕФТЕПРОДУКТОВ» Биттеева М.Б., Бирюков В.В., Щеблыкин И.Н.,
15. «Очищення промислових стічних вод на основі термодинамічного прогнозування взаємодії мікроорганізмів з металами та радіонуклідами» О.Б. Таширев Киев 2005
16. Косогорова Э.А. защита полевых и овощных культур от болезней. М.: Росагропромиздат. 1991
17. Коршунова А.Ф. и др. Корневые гнили зерновых в Сибири. – Л.: Колос, 1974
18. http://www.fermenter.ru/content/page_175_0.html