| Скачать .docx | Скачать .pdf |
Реферат: Темпы развития парциальных кустов брусники обыкновенной Vaccinium vitis-idaea L
МИНИСТЕРСТВО ОБРАЗОВАНИЯ РОССИЙСКОЙ ФЕДЕРАЦИИ
МАРИЙСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
Кафедра ботаники, экологии
и физиологии растений
Дипломная работа
студентки Истоминой Анастасии Николаевны
группы Б-53 биолого-химического факультета
Тема: Темпы развития парциальных кустов брусники обыкновенной Vaccinium vitis - idaea L .
Заведующий кафедрой ботаники,
экологии и физиологии растений МарГУ
_______ д.б.н., проф. Н.В. Глотов
Научные руководители:
_______ д.б.н., проф. Н.В. Глотов,
_______ преподаватель кафедры ботаники,
экологии и физиологии растений Л.В. Прокопьева
Рецензент:
_______ к.б.н., старший преподаватель кафедры
зоологии и прикладной экологии Н.В. Ившин
Студент:
_______ А.Н. Истомина
Йошкар-Ола
2003
СОДЕРЖАНИЕ
ВВЕДЕНИЕ 3
1. Обзор литературы 4
1.1. Темпы развития растений 4
1.1.1. Представление о поливариантности онтогенеза 4
1.1.2. Влияние экологических факторов на
темпы развития 10
1.2. Распространение и фитоценотическая приуроченность
брусники 22
2. Материалы и методика 25
2.1. Объект исследований 25
2.2. Место исследований 27
2.3. Методы исследований 30
3. Результаты и обсуждение 34
3.1. Экологическая характеристика местообитаний 34
3.2. Темпы развития брусники обыкновенной 39
3.2.1. Выживаемость парциальных кустов 39
3.2.2. Пропуск возрастных состояний 42
3.2.3. Продолжительность возрастных состояний 44
3.2.4. Итоговые возрастные спектры 50
ВЫВОДЫ 54
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ 55
БЛАГОДАРНОСТИ 63
ПРИЛОЖЕНИЕ 64
ВВЕДЕНИЕ
Темпы развития особей представляют собой один из существенных признаков популяций растений. От этой стороны организации зависят способность популяционной системы к самоподдержанию и её устойчивость, поскольку различие особей по темпам развития является одним из механизмов регулирования численности и возрастного состава ценопопуляций.
Для разработки режима рационального использования ценопопуляций (ЦП) брусники на современном научном уровне необходимы не только данные по запасам сырья и плотности запаса сырьевых частей, но также и знание темпов развития её ценопопуляций.
Изучение темпов развития как одно из направлений популяционных исследований является необходимым условием при решении практических задач охраны хозяйственно ценного вида, а также рационального использования и восстановления естественных и создания искусственных сообществ.
Целью данной работы является изучение темпов развития парциальных кустов брусники обыкновенной Vaccinium vitis - idaea L. в Государственном природном заповеднике «Большая Кокшага» Республики Марий Эл.
В работе решались следующие задачи:
1. Дать эколого-ценотическую характеристику местообитаний брусники;
2. Определить темпы развития парциальных кустов брусники в различных местообитаниях.
1. ОБЗОР ЛИТЕРАТУРЫ
1.1. ТЕМПЫ РАЗВИТИЯ РАСТЕНИЙ
1.1.1. Представление о поливариантности онтогенеза
Темпы развития, по определению Л.И.Воронцовой (1967), представляют собой относительную скорость прохождения растениями большого жизненного цикла. Т.А.Работнов (1950) отмечал, что показателем высокой жизненности вида в фитоценозе являются быстрые темпы перехода ювенильных особей в генеративные. В возрастном спектре популяций это выражается небольшой долей переходных особей.
А.А.Уранов (1960), рассматривая вопрос о темпах развития вида в растительном сообществе, различает у многолетних растений при нормальном состоянии их вегетативных органов два случая ускорения развития: временное, быстро проходящее, связанное с наличием в фитоценозе «свободной жизненной ниши», и длительно сохраняющееся. В первом случае увеличение численности взрослых особей приводит к замедлению их развития, во втором — при накоплении взрослых особей быстрые темпы развития сохраняются как на восходящей, так и на нисходящей части жизненного цикла растения, при этом старые особи относительно быстро отмирают и освобождают место молодым растениям.
Анализируя основные закономерности развития растений, Д.А.Сабинин (1963) отмечал две наиболее общие черты, свойственные всем растительным организмам: расчленённость онтогенеза и многовариантность развития. Первая предполагает существование качественных различий разных этапов онтогенеза при новообразовании органов и структур, вторая означает возможность реализации разнообразных путей онтогенеза при воздействии экзогенных и эндогенных факторов.
При популяционных исследованиях были обнаружены различные модификации онтогенетических состояний у луговых трав: тимофеевки луговой — Phleum pratense L., люцерны серповидной — Medicago falcata L., лерхенфельдии извилистой — Lerchenfeldia flexuosa (L.) Schur. Эти явления получили название многовариантности, мультивариантности, поливариантности развития. Одновременно они были описаны также у некоторых степных растений: подмаренника русского (Galium ruthenicum Willd.) и полыни Artemisia lerchiana Web. exStechn. (Жукова, 1995).
В дальнейшем Л.И.Воронцовой и Л.Б.Заугольновой (1978) было выделено три типа поливаринтности развития растений:
1) по уровням жизненности;
2) темпам развития;
3) морфологической структуре.
Накопление материалов показало, что поливариантность развития — явление чрезвычайно широко распространённое в растительном мире, и оно может рассматриваться как одно из проявлений модификационной изменчивости (Ценопопуляции растений, 1988; Жукова, 1995). В настоящее время различают пять типов поливариантности: размерную, морфологическую, способов размножения и воспроизведения, ритмологическую и временную (Ценопопуляции растений, 1988).
Л.А. Жукова и А.С. Комаров (1991) предлагают выделять в общем надтипе динамической поливариантности два типа: ритмологический и собственно динамический, или временной (по длительности возрастных состояний и темпам развития).
Поливариантность по темпам развития или временная поливариантность выражается в различной скорости индивидуального развития (Воронцова, Заугольнова, 1978). Это наименее изученный к настоящему времени тип поливариантности онтогенеза. Одним из важнейших его параметров является продолжительность онтогенетических состояний, или онтохрон. Определить его можно после многолетних наблюдений за маркированными растениями. Так, подобными наблюдениями за маркированными луговыми растениями модельных видов показано, что длительность пребывания особи в одном онтогенетическом состоянии (онтохрон) может варьировать в зависимости от темпов развития: то более быстрых, то более медленных, вплоть до временного прекращения развития (вторичный, или временный покой) (Ценопопуляции растений, 1988; Жукова, 1995).
Существование внутрипопуляционных групп, отличающихся по темпам развития, впервые было описано в лесоводческой литературе, когда в одновозрастных насаждениях различных древесных пород были обнаружены растения с пониженной жизненностью и замедленной скоростью роста (Завадский, 1968; Ценопопуляции растений, 1988).
В результате многолетних наблюдений в естественных и искусственных луговых фитоценозах (Ермакова, Жукова, 1985; Жукова, Шейпак, 1985) были описаны пять классов временной поливариантности: ускоренное развитие, нормальное развитие, замедленное развитие, вторичный покой и реверсии в более молодые возрастные состояния вследствие омоложения.
1 класс — ускоренное развитие. В луговедческой литературе неоднократно отмечалось ускоренное развитие луговых трав в посевах (Смелов, 1966; Работнов, 1984), прежде всего за счёт уменьшения прегенеративного периода. Так, некоторые особи ежи сборной Dactylis glomerata L. и овсяницы луговой Festuca pratensis L. завершают полный онтогенез за 4-5 лет (Ермакова, Жукова, 1985). Особи луговика дернистого на вырубках и залежах иногда зацветали на 3-й год (вместо 5-7-го).
Однако возможен и другой путь ускорения развития — пропуск одного или нескольких возрастных состояний либо целого периода. Такие явления были описаны у ежи сборной, овсяницы луговой, подорожника большого (Plantago major L.), лютика ползучего (Ranunculus repens L.) в посевах, где зарегистрированы пропуски имматурного, виргинильного, молодого, средневозрастного и старого генеративного и субсенильного состояний. Максимальный интервал — 5 возрастных состояний: от имматурного до сенильного (Ермакова, Жукова, 1985).
2 класс — нормальные темпы развития. Анализ временной поливариантности требует выработки критериев нормального развития. Вопросы о границах между нормой и отклонением для любых биологических объектов достаточно сложны. Нормальными темпами развития особей были признаны такие, при которых продолжительность каждого возрастного состояния является модой для данной ценопопуляции. Как показали наблюдения, в луговых агроценозах доля растений, развивающихся нормальными темпами, составляет 50-60%.
3 класс — замедленное развитие — может проявляться в виде задержки растений в тех или иных возрастных состояниях, прежде всего в прегенеративном периоде. Так, у луговика дернистого на пойменных лугах Северной Двины встречаются 7-8-летние ювенильные растения, хотя обычно в этом состоянии растение находится не более одного вегетативного периода (Ценопопуляции растений, 1988). У борщевика сибирского прегенеративный период может продолжаться до 20 лет (Работнов, 1978). У луговых трав задержка развития может приводить к пропуску очередного возрастного состояния.
Темпы развития растений связаны с их жизненным состоянием: у многолетников отмечается тенденция к увеличению продолжительности жизни при низком уровне жизненности (Ценопопуляции растений, 1988). А.А.Чистякова (1978, 1982) при изучении темпов развития основных пород в широколиственных лесах обнаружила, что у деревьев и кустарников в экологически неблагоприятных условиях неопределённо долгая задержка в прегенеративном периоде приводит к образованию стланниковых и полустланниковых форм, практически никогда не достигающих генеративного периода и существующих в омоложенном состоянии.
4 класс — вторичный или временный покой. Является крайним выражением задержки развития. Это явление было детально описано Т.А.Работновым у ветреницы (Anemonoides sp .) на субальпийских лугах Кавказа. Дальнейшие наблюдения показали, что вторичный покой гораздо более распространённое явление среди растений, чем предполагалось ранее. В луговых агроценозах в каждый срок наблюдений в течение вегетационного периода от 2,3 до 25,0% особей ежи сборной и от 2,0 до 17,5% особей овсяницы луговой находятся в состоянии вторичного покоя. Указывается, что при воздействии высоких доз удобрений, как и высокой плотности посевов, доля растений, относящихся к этой форме временной поливаринтности, значительно возрастает. Это позволяет оценить вторичный покой как одно из важнейших приспособлений к крайне неблагоприятным условиям существования (Куркин, 1976; Работнов, 1978, 1984; Ценопопуляции растений, 1988).
5 класс — реверсии в более ранние состояния или омоложение. Степень омоложения различна у видов, принадлежащих различным типам онтогенеза и разным биоморфам: явнополицентрическим, неявнополицентрическим и моноцентрическим. Поливариантность омоложения заключается в возможности существования в пределах одной ценопопуляции разных реверсий: например, у липы сердцелистной Tilia cordata Mill. (Чистякова, 1978), у Deschampsia flexuosa на вырубках (Ценопопуляции растений, 1988).
Длительные наблюдения за маркированными особями ряда луговых трав различных биоморф (ежа сборная, овсяница луговая, подорожник ланцетовидный Plantago lanceolata L., лютик ползучий, тмин обыкновенный Carum carvi L., одуванчик лекарственный Taraxacum officinalis L., кровохлёбка лекарственная SanguisorbaofficinalisL.) показали, что на протяжении индивидуальной жизни одного растения эти формы динамических изменений могут сменяться одна другой и неоднократно повторяться (Ценопопуляции растений, 1988). Задержка на любом из этапов, как на протяжении от одного состояния до другого, так и при переходе во временный покой увеличивает общую продолжительность онтогенеза.
Систематизация вариантов динамической поливариантности, правила перехода из одного состояния в последующие могут быть представлены в виде таблиц запретов и разрешений переходов на протяжении полного онтогенеза (Жукова, 1995). Так, такие таблицы для некоторых луговых трав показывают, что 29 переходов (35,8%) у ежи сборной и овсяницы луговой не могут быть осуществлены из-за биологических свойств этих видов, и ни разу не были зарегистрированы. Например, переход из ювенильного состояния в средневозрастное генеративное или реверсии из субсенильного в молодое или средневозрастное генеративное состояние. Такие таблицы могут служить своеобразной видовой характеристикой, характеристикой биоморфы и являться путём выявления особенностей экологической и фитоценотической обстановки, позволяя выявить специфичность воздействия различных факторов на данную ценопопуляцию. Длительными наблюдениями за маркированными особями ряда луговых трав установлено, что на протяжении полного онтогенеза одной особи темпы её развития могут изменяться на разных этапах, особенно при большой его длительности. Возможность разнообразных сочетаний нормального, ускоренного, замедленного развития, вторичного покоя, а также нарушений последовательности состояний создаёт теоретическую предпосылку существования множества путей индивидуального развития. По этим данным с помощью IBM было рассчитано максимально возможное число путей онтогенеза для ряда модельных видов в различных экологических ситуациях. Дальнейшие расчёты позволили определить в каждом конкретном варианте максимально возможное число реализованных путей развития и число фактически реализуемых путей. Отмечено, что неблагоприятные условия вызывают массовую гибель, а немногие выжившие экземпляры могут воплотить лишь малое число возможных путей. В то же время наиболее благоприятные условия существования ускоряют развитие особей, сокращают продолжительность онтогенетических состояний, что также уменьшает разнообразие возможных путей онтогенеза.
Один из главных аспектов дальнейшего углубления концепции поливариантности онтогенеза — выявление генетической компоненты изменчивости признаков элементов популяции (Жукова, 1995). По мнению И.И.Шмальгаузена (1968), в основе поливариантности развития лежит генетический полиморфизм и модификационная изменчивость в пределах генотипа. Адаптивные модификации могут быть выражены на любом этапе онтогенеза.
Результаты наблюдений М.М.Палёновой (1991) в посемейных посадках клевера ползучего Trifolium repens L. показали статистически значимые межсемейные различия темпов развития особей. Характер изменчивости этого признака статистически интерпретируется как «в значительной мере генетически обусловленный». Дальнейшее сравнение отдельных семей по темпам развития доказало, что в искусственно созданных популяционных локусах Trifolium repens чётко выявляются семьи с задержкой перехода в генеративное состояние. Автор подчёркивает чрезвычайную важность того, что семьи сохраняют не только проявление признака, но и самобытность тенденций изменения этого признака, его динамики. Следовательно, существуют реальные доказательства того, что динамическая поливариантность у клевера ползучего в значительной мере обусловлена генетически. Можно предположить, что это справедливо и для других видов растений.
1.1.2. Влияние экологических факторов на темпы развития
Ценопопуляция, подобно любой биологической системе, характеризуется определённой структурой, включающей помимо некоторого набора элементов пространственное их размещение, а также характер связей между ними и с окружающей средой. Современное понимание структуры охватывает не только взаимное размещение частей в пространстве, но и динамику их во времени, поскольку каждая система существует одновременно во времени и пространстве.
Для дальнейшей разработки механизмов устойчивости и динамичности видов в ценозе необходима количественная оценка основных процессов в ценопопуляциях и выявление их связей с экотопическими и ценотическими факторами. Такими процессами можно считать характер и степень изменения численности и возрастного состава во времени, интенсивность отмирания особей, темпы их онтогенетического развития (Ценопопуляции растений, 1977).
Экологические факторы различаются по природе (абиотические, биотические, антропогенные) и по характеру воздействия на живые системы (организмы, популяции, ценозы). Среди абиотических факторов различают следующие типы: вещественно-энергетический (трофический), летальный (и повреждающий) и информативный (сигнальный). Воздействие трофических факторов на численность и возрастной состав ценопопуляций может носить менее выраженный характер, так как оно опосредовано через изменение интенсивности размножения, темпы развития и отмирания растений (Ценопопуляции растений, 1988). В качестве примеров трофических воздействий на ценопопуляции можно привести изменение темпов развития в более аридных условиях.
В литературе нет сведений об изменениях темпов развития растений в разных географических зонах. Имеются лишь общие указания, что при ухудшении условий произрастания продолжительность жизни увеличивается. Отмечается, что на аридных и субаридных территориях с усилением аридности старение растений ускоряется.
Л.И.Воронцовой (1967) впервые сделана попытка показать влияние зональных условий на возрастные спектры популяций. Так, на примере типчака Festuca sulcata L. доказано ускорение темпов развития в благоприятных условиях. Накопление стареющих и сенильных растений в данном случае не препятствует ускоренному развитию вегетативных особей, так как сенильные экземпляры быстро отмирают. В ксероморфных ассоциациях, приуроченных к верхним частям западин и микроплакорным участкам, темпы развития типчака замедляются, в связи с этим в популяциях накапливаются переходные особи. Тенденция к ускорению жизненного цикла осуществляется у данного вида при более низкой численности и некотором угнетении его особей. Автор указывает, что ускорение темпов развития типчака следует расценивать как приспособление к суровым условиям, в данном случае к усилению аридности климата, когда конкуренция между растениями за воду становится особенно напряжённой.
На возможность быстрого развития при некоторой подавленности особей в неблагоприятных условиях указывает А.А.Уранов (1960), рассматривая это явление как биологическое приспособление, выгодное не для особи, а для популяции в целом. Ускорение темпов развития типчака гарантирует ему большую устойчивость в ценозе и позволяет быстро захватывать свободное жизненное пространство.
Изменчивость возрастного состава популяций в зависимости от внешних и ценотических условий отражена в работах многих авторов.
Так, Н.И.Шориной (1967) установлено, что в зависимости от условий может несколько изменяться онтогенез безвременника. Е.В.Тюрина (1975) отмечает изменение ритма роста и развития Peucedanum batealense (Redow.) C. Koch. в зависимости от экологических условий и при интродукции. Л.Е.Гатцук (1967) установлено, что чрезвычайно высокий темп развития характерен для копеечника кустарникового Hedysarum fruticosum Pall., что сказывается на длительности его возрастных состояний, на скорости перехода из одного состояния в другое и на пропуске в ряде случаев отдельных фаз. Высказывается предположение, что скорость развития копеечника зависит от относительно благоприятных экологических условий на песках степной и лесостепной зоны, в частности, увлажнения, аэрации субстрата, освещения, летнего тепла, отсутствия конкуренции. Столь быстрое развитие, несомненно, полезно в условиях подвижного субстрата. Это полезное свойство могло закрепиться под действием естественного отбора.
В.А.Черёмушкина (1998) при описании хода онтогенеза лука сизого Allium glaucum Schrad. в разных эколого-фитоценотических условиях установила его поливариантность, играющую существенную роль в формировании структуры популяций. Отмечается, что у лука сизого можно встретить все типы поливариантности, причём онтогенез части особей идёт по сокращённому пути: минуя генеративный период, виргинильные растения переходят в субсенильное или сенильное состояние. Определяющими факторами, влияющими на ход онтогенеза, оказались общее проективное покрытие фитоценоза и задернованность почвы.
М.Р.Трубиной (1998) приводятся факты задержки темпов онтогенетического развития особей скерды кровельной в условиях загрязнения. Кроме того, сообщается, что выращивание растений на чистой золе вызывает значительную задержку темпов развития у многолетних видов.
Л.К.Давидюк (1987) при изучении динамики численности и возрастного состава ценопопуляций представителей травяного покрова пойменных дубрав (подмаренник волжский Galium sp ., ястребинка зонтичная Hieracium umbellatum L.) обнаружил ускорение темпов онтогенетического развития в первый год рубки, совпавший с засухой. При этом наблюдался переход большинства особей в следующее возрастное состояние и увеличение в составе ценопопуляций относительной доли растений, перешедших в состояние вторичного покоя и вторичной вегетации. Отмечается, что для изученных видов характерно выпадение отдельных фаз онтогенетического развития, отмирание на любой стадии большого жизненного цикла, переход при неблагоприятных условиях в состояние вторичного покоя и вторичной вегетации. Таким образом, резкое изменение условий произрастания в результате рубки древостоя вызывает ускорение темпов онтогенетического развития и элиминацию ценопопуляций вида.
Е.С.Закамская и Г.М.Панова (1998) при изучении возрастной структуры ценопопуляций костяники Rubus saxatilis L. в различных экологических условиях установили, что при неблагоприятных условиях внешней среды, каковым считается избыточное освещение, происходит раннее старение, и особи из прегенеративного онтогенетического периода сразу переходят в постгенеративный период.
Л.А.Жуковой (1995) обнаружена поливариантность онтогенеза по темпам развития у многих луговых растений. Так, динамическая поливариантность отмечена, несмотря на стабильность жизненной формы, у плотнодерновинных злаков. Указывается, что она является ответной реакцией на воздействие неблагоприятных условий, поэтому только часть особей проходит полный онтогенез. При ухудшении жизненного состояния особей может произойти пропуск одного или нескольких онтогенетических состояний.
В условиях сухих и богатых почв отмечено максимальное количество побегов ковыля перистого Stipa pennata L. в дерновине, а на популяционном уровне та же совокупность факторов приводит к уменьшению численности подроста в результате его высокой смертности и как следствие, к общей низкой плотности ценопопуляций. В этих условиях темпы развития особей оказываются высокими, и растения отмирают в генеративном состоянии, поэтому в ценопопуляциях отсутствуют субсенильные и сенильные растения. Зависимость между плотностью ценопопуляции и темпами развития особей можно рассматривать как проявление отрицательных обратных связей, обеспечивающих регуляцию ценопопуляций.
На примере ковыля перистого показано, что высокая фитомасса ценопопуляции обеспечивается за счёт быстрых темпов развития при относительно невысокой плотности или за счёт высокой плотности при сочетании внешних факторов, ограничивающих мощность растений, но не их выживание. Это обычно сочетается с замедленными темпами развития, что приводит к накоплению старых растений в ценопопуляции (Ценопопуляции растений, 1988).
Влияние плотности на темпы развития особей синюхи голубой Polemonium coeruleum L. и валерианы лекарственной Valeriana officinalis L. в природных и искусственных ценопопуляциях изучалось Н.В.Илюшечкиной (1998). Наблюдения за маркированными особями позволили изучить динамическую поливариантность онтогенеза и выделить классы развития. Для синюхи голубой обнаружено пять классов развития: нормальное, ускоренное, замедленное развитие, вторичный покой и омоложение (или реверсия); для валерианы лекарственной только четыре класса развития, отсутствовал вторичный покой. Распределение классов развития синюхи голубой показало, что при изменении плотности посадки наблюдается перераспределение частот разных классов развития. Можно видеть, что с одной и той же частотой при разной плотности растения уходят во вторичный покой, но резко возрастает число растений с ускоренным развитием. На основании проведённых наблюдений переходов одного онтогенетического состояния в другое составлены схемы путей онтогенеза валерианы лекарственной и синюхи голубой.
Об изменчивости общей продолжительности полного онтогенеза и отдельных его периодов можно судить по данным, полученным в посадках многолетних трав. Так, у ежи сборной и овсяницы луговой средняя длительность полного онтогенеза, генеративного и постгенеративного периодов вдвое длиннее в контрольных вариантах с фосфорно-калийными удобрениями, чем в опытах с добавками азотных удобрений. Минимальная продолжительность онтогенеза для обоих видов отмечена в смешанном посеве с полным удобрением, что подчёркивает дополнительное влияние межвидовой конкуренции. Протяжённость каждого возрастного периода, в свою очередь, определяет онтохроны возрастных состояний. Масштабы их изменчивости ещё больше, причём максимальная и минимальная продолжительность у большинства онтогенетических состояний этих растений не совпадает в разных вариантах опытов. Только для субсенильного состояния зарегистрирована минимальная длительность в чистых, загущенных посевах с полным удобрением (Жукова, 1995).
При изучении воздействия гербицидов на растения Л.А.Жуковой и О.А.Шейпак (1985) обнаружено необычайное разнообразие путей онтогенеза у подорожника большого. Анализ данных, полученных в искусственных и естественных ценопопуляциях подорожника большого, показал, что генеративные особи наиболее стойки и способны сохранить характер онтогенеза. Однократная и двукратная обработка 2,4-D в более сильной концентрации вызывает в ряде случаев у генеративных растений задержку развития или ускоренное развитие с пропуском одного или нескольких возрастных состояний и лишь иногда последующее отмирание. Уникальный случай мгновенного старения был зарегистрирован при двукратной обработке аминной солью ювенильных растений, которые через два месяца перешли в субсенильное состояние. Отмечается, что биологической причиной старения при воздействии 2,4-D, вероятно, следует считать более быструю реализацию потенциальных возможностей растений. Кроме того, проведённые опыты позволили экспериментально установить возможность перехода некоторых растений подорожника в состояние вторичного покоя после двукратной обработки 2,4-D. Установлено, что у старых генеративных и особенно у субсенильных растений под воздействием гербицидов начинаются процессы омоложения.
Таким образом, наблюдаемое многообразие путей онтогенетического развития представляет собой сложные адаптации организменного уровня к экстремальным ситуациям.
В литературе практически нет данных об исследованиях темпов развития брусники. Результаты исследований онтогенеза, возрастной структуры ценопопуляций и темпов развития растения сходной жизненной формы черники Vaccinium myrtillus L. отражены в работах некоторых авторов.
Так, Е.В.Шавриной (1991) установлено, что в экологическом ряду сосняков с увеличением влажности почвы наблюдается тенденция к некоторому увеличению длительности жизненного цикла черники.
И.Г.Соколова (1991) при изучении пространственно-временной структуры ценопопуляций черники установила, что темпы развития парциальных кустов черники в березняке и осиннике мало отличимы и составляют почти 7 лет. В ельнике темп развития парциальных кустов больше на 1-2 года.
Кроме того, в каждой ценопопуляции наблюдалась поливариантность развития парциальных кустов черники, которая осуществляется по следующей схеме:
![]()
![]()
![]()
![]()
![]()
![]()
![]() j v g1 g2 g3 ss s
j v g1 g2 g3 ss s
![]()
![]() sss
sss
К.А.Миронов (1987) изучал восстановление черники после пожаров в лесном Среднем Заволжье. Анализ возрастной структуры пирогенных и естественных ценопопуляций показал, что низовые пожары умеренной интенсивности способствуют удлинению жизненного цикла парциальных кустов. Так, если на гари 12-летней давности средний возраст кустов составляет 8,3 года, то в естественных ценопопуляциях он равен лишь 6,6 годам. Это обусловлено, по-видимому, стерилизующим влиянием огня на среду обитания ягодника, что приводит к снижению повреждаемости последнего вредителями и болезнями.
Г.И.Бумар и П.Н.Таргонский (1989) отмечают, что исследованию ценопопуляций и парциальных кустов брусники посвящён ряд работ, выполненных главным образом в слабо затронутых хозяйственной деятельностью лесах Западной Сибири. В 1986-1988 гг. ими были проведены исследования по определению возрастного состава популяций брусники в Украинском Полесье, на южной границе её сплошного распространения, в следующих фитоценозах: сосняке молиниевом, сосняке бруснично-зеленомошном и гари 1986 г., естественно возобновившейся к 1988 г. берёзой повислой Betula pendula Roth. и сосной обыкновенной Pinus sylvestris L. В условиях Украинского полесья при высокой освещённости травяного яруса (на гарях и вырубках) брусника интенсивно разрастается, а, следовательно, её проективное покрытие в данных условиях местопроизрастания выше, чем под пологом насаждений. Исследованные популяции независимо от типа леса характеризуются стабильностью численности и возрастных параметров. Они также не зависят от инвазии семян с других сообществ и от колебаний экологических факторов. В спектре возрастного состава первых двух популяций преобладают зрелые генеративные особи (25,4-25,6%), в спектре третьей — молодые генеративные (28,8%).
Е.А.Мазная (2001) провела оценку жизненного состояния и возрастной структуры ценопопуляций Vaccinium myrtillus L.иVaccinium vitis- idaea L., совместно произрастающих в сосняках кустарничково-лишайниковых и находящихся под постоянным воздействием атмосферного загрязнения — выбросов комбината «Североникель», с целью определения возрастной и виталитетной структуры ценопопуляций. Выяснилось, что независимо от возраста сосны и степени повреждения лесных экосистем возрастной спектр ценопопуляций сохраняет полночленность, что указывает на довольно высокую приспособленность видов к экстремальным условиям, вызванным интенсивным атмосферным загрязнением. Однако при этом увеличивается доля субсенильных и сенильных особей. Абсолютный максимум в возрастном спектре независимо от степени загрязнения приходится на группу молодых виргинильных парциальных кустов (25,9-43,3%).
Е.Е.Тимошок и Н.В.Паршина (1992) впервые описали основные признаки возрастных состояний парциальных кустов брусники, количественные и качественные. К количественным признакам относятся высота растения, диаметр кроны, возраст растения, длина годичного прироста, число цветков; к качественным — тип ветвления, степень цветения и степень отмирания.
Для описания возрастных состояний парциальных кустов также были использованы общие характеристики качественных и количественных признаков возрастных состояний, разработанные Т.А.Работновым (1950), с дополнениями. Было выделено семь возрастных состояний парциальных кустов: виргинильные, молодые генеративные, зрелые генеративные, старые генеративные, потенциально генеративные, субсенильные и сенильные. Отмечается, что в исследованных ценопопуляциях брусники в Западной Сибири, как и в её ценопопуляциях в европейской части России, она проходит весь цикл развития до образования семян и диссеминации, но самоподдержание ценопопуляций осуществляется путём интенсивного вегетативного размножения начиная с молодого виргинильного состояния. Во всех изученных ценопопуляциях не было обнаружено проростков и молодых особей брусники семенного происхождения, что объясняется низкой численностью генеративных парциальных кустов и в связи с этим низким плодоношением, а также некоторыми дополнительными факторами. В связи с этим изученные ценопопуляции названы неполночленными. При этом указывается, что полученные данные согласуются с данными других авторов, изучавших размножение брусники в западной Европе и на российском Дальнем Востоке.
Е.Е.Тимошок (1998) отмечает, что полный онтогенез брусники, так же как и черники, складывается из последовательной смены поколений сменяющих друг друга парциальных кустов. Возрастная структура её ценопопуляций определяется длительностью жизни и особенностями развития парциальных кустов. Проведённые исследования показали, что в онтогенетических спектрах ценопопуляций преобладали виргинильные кусты, постоянно высоким было и участие потенциально генеративных кустов. В кедрово-сосновом бруснично-зеленомошном лесу, сосняке бруснично-зеленомошном и сосняке бруснично-лишайниково-зеленомошном, в горных лиственничном и лиственнично-берёзовом бруснично-зеленомошных лесах, в кедровом бруснично-мохово-лишайниковом редколесье и в бруснично-лишайниковой тундре отмечено значительное участие генеративных кустов. При ухудшении условий местообитания, в восстанавливающихся после рубок лесах и при достаточно низкой рекреационной нагрузке участие генеративных парциальных кустов снижается. При интенсивной рекреационной нагрузке генеративные парциальные кусты отсутствуют. В большинстве исследованных ценопопуляций в равнинных и горных лесах, подгольцовых редколесьях и высокогорных тундрах отмечено низкое участие субсенильных и сенильных парциальных кустов, что связано с отмиранием парциальных кустов на всех предшествующих этапах онтогенеза. Высокое участие субсенильных парциальных кустов характерно для ценопопуляций брусники в условиях высокой рекреационной нагрузки, что объясняется тем, что большая часть парциальных кустов проходит ускоренный онтогенез (переходя из потенциально генеративного и виргинильного в субсенильное состояние), вследствие чего и происходит их накопление в этом возрастном состоянии.
Высокое участие виргинильных парциальных кустов в исследованных ценопопуляциях связано с интенсивным и регулярным вегетативным самоподдержанием ценопопуляций во всех исследованных местообитаниях.
Таким образом, быстрые темпы развития не всегда служат показателем оптимальных условий. Разнообразие климатических, экологических условий и способов антропогенного воздействия способствует разным темпам развития особей во времени. В различных экологических ситуациях меняется не только общая продолжительность полного онтогенеза, отдельных состояний и периодов, но и их соотношения. Все эти проявления временной поливариантности могут вызвать серьёзные изменения возрастной структуры ценопопуляций и их динамики (Жукова, 1995). Темпы развития являются одним из признаков, характеризующих тип стратегии на организменном уровне (Смирнова, 1987). На популяционном уровне поливариантность онтогенеза является важнейшим адаптационным механизмом, определяющим динамическую гетерогенность ценопопуляций и способствующим их устойчивости, что является существенным резервом наследственной изменчивости (Ценопопуляции растений, 1988).
1.2. РАСПРОСТРАНЕНИЕ И ФИТОЦЕНОТИЧЕСКАЯ
ПРИУРОЧЕННОСТЬ БРУСНИКИ
Брусника имеет обширный голарктический ареал, который охватывает большую часть Европы — от Скандинавии (71°7' с.ш.) до Балкан и от Пиреней до Урала. Встречается брусника также в Японии, Северной Америке (от Аляски до Лабрадора и Гренландии) (Пояркова, 1952). В Европе растение встречается на высоте от 200 до 1500 м над уровнем моря (Ritchie, 1955).
Брусникараспространена почти по всей территории бывшего СССР, кроме южных районов европейской части, всей Средней Азии, подавляющей части Казахстана и Закавказья. Северная граница ареала проходит по морскому побережью, а также брусника встречается на о. Колгуев, южном острове Новой Земли. Восточная граница идёт по побережью Тихого океана, захватывая Командорские о-ва, Камчатку, о-ва Курильской гряды, всё побережье Охотского моря, о. Сахалин и Приморье. Изолированные участки ареала брусники обнаружены в высокогорьях Кавказа (41-44° с. ш.) и близ устья Днепра (Атлас ареалов…, 1976).
Брусника, по А.Г.Крылову (1984), относится к боровому типу ценоэлемента, к таёжно-боровому подтипу, к ксеромезофильным, олиготрофным вечнозелёным кустарничкам.
А.И.Толмачёв (1954) определяет бруснику как несомненно лесной вид. Большинство близких видов рода Vaccinium произрастает в горных лесах запада Северной Америки. «Тропические» черты облика этого вечнозелёного кустарничка находятся в полном соответствии с распространением большей части его близких сородичей в тропических горных районах.
Брусника произрастает, главным образом, в хвойных, часто преобладая в травяно-кустарничковом ярусе (Кормовые растения, 1956), лесах: сосновых, кедровых, пихтовых, лиственничных, а также в смешанных, лиственных лесах и на сфагновых болотах (Ильинская, 1963; Тимошок, 1996). В горных и вересковых местообитаниях брусника занимает господствующее положение (Ritchie, 1955). Отмечено, что брусника занимает значительное место среди кустарничков мезотрофных болот (Елина и др., 1984).
В условиях подзоны средней тайги Кировской области брусника находит свой экологический оптимум в сосняках брусничных, здесь древостой достигает возраста 12-80 лет, высота 4-25 м, сомкнутость крон 0,3-0,6, на песчаных и супесчаных почвах разной степени оподзоленности (Егошина, 1986).
Популяции брусники в Республике Марий Эл имели 100% встречаемость, обилие сор-2 в сосняке брусничном — на слабоподзолистых, крупнопесчаных почвах, в сосняке черничном — на пониженных местах с близким залеганием грунтовых вод и влажными сильноподзолистыми почвами (Иванова, Ефремова, 1998).
В условиях верховых болот (сфагновые сосняки, заболоченные сфагновые березняки) брусника является компонентом микроассоциаций на буграх и микроповышениях, так как плохо переносит анаэробность субстрата и конкуренцию со сфагновым покровом (Солоневич, 1956; Ильинская, 1963). Брусника встречается на торфяных, торфяно-минеральных и болотно-минеральных почвах (Bozenna, 1986).
Наиболее обильнабрусникана дренированных, большей частью кислых и бедных питательными веществами подзолистых почвах. В некоторых районах (Центральная Якутия) она хорошо произрастает на относительно богатых почвах, с близким залеганием карбонатных горизонтов (различные варианты буро-серых неоподзоленных или осолодевших почв) (Кормовые растения, 1956). Однако брусника может произрастать и на почвах, где содержание оксида кальция не превышает 0,8% (Богданова, Муратов, 1978). Следовательно, кустарничек может произрастать на сухих, очень бедных почвах или бедных как минеральными, так и органическими компонентами почвах (Bozenna, 1986).
К влажности почвы брусника также нетребовательна — встречается как на сухих, так и на влажных почвах (Булгаков и др., 1987). Уровень стояния вод –12…-45 см (это наиболее сухие экотопы) (Боч, Смагин, 1993). Большей частью с увеличением сухости почв обилие брусники в травяно-кустарничковом покрове сильно вырастает, особенно на открытых, хорошо освещённых участках (Солоневич, 1956).
V. vitis- idaea относительно мало теневынослива и поэтому наиболее обильна в светлохвойных или слабо сомкнутых темнохвойных лесах (Кормовые растения, 1956). Брусника светолюбива и лучше плодоносит на открытых местах или низкоплотных древостоях (Булгаков и др., 1987).
Брусника отличается достаточно широкой амплитудой по отношению к кислотности почв. По данным разных авторов рН колеблется от 2 до 6 (Зворыкина, 1973; Сокол, 1980; Ипатов, Кирикова, 1997; Викторов, Ремезова, 1988).
Приспособленность к жизни в холодных, умеренно и средне увлажнённых местообитаниях с кислыми и очень бедными почвами послужила основанием для выделения некоторых кустарничков, в том числе и брусники, в экологическую группу психрофитов (Шенников, 1950). Ксероморфное строение психрофитов связано с низкой температурой среды, физиологической сухостью повышенно влажных почв и с замедлением темпов всех физиологических процессов (Солоневич, 1956).
2. МАТЕРИАЛЫ И МЕТОДИКА
2.1. Объект исследования
Брусника обыкновенная Vaccinium vitis - idaea L. — длиннокорневищный вечнозелёный кустарничек из семейства вересковых — Ericaceae.
Удлинённые побеги брусники полициклические, поликарпические, укореняющиеся и приподнимающиеся. Листья кожистые, многолетние, очередные, эллиптические или обратно яйцевидные, тупые или слегка выемчатые, слегка зазубренные или цельнокрайние с завороченным краем, 5-27 мм длиной, 3-12 мм шириной, на коротких опушённых черешках 0,5-3,0 мм длиной, сверху тёмно-зелёные, снизу бледные с тёмно-бурыми рассеянными желёзками.
Основание побега переходит в гипогеогенное корневище, залегающее на глубине 2-10 см, обычно очень длинное — до 18 м длиной, шнуровидное, зеленовато-коричневого цвета. От корневища отходят тонкие, короткие и редко расположенные придаточные корни.
Соцветие — короткая, густая, поникающая кисть с 2-8 (редко 18) цветками, формируется в верхней части прошлогоднего годичного побега. Цветки на коротких, опушённых, красноватых цветоножках, обладают слабым, но приятным запахом. Чашечка спайнолистная 4-зубчатая, с короткими округлыми красноватыми зубцами, венчик спайнолепестный колокольчатый, бледно-розовый, с четырьмя лопастями. Тычинок 8, с волосистыми тычиночными нитями, пыльники без придатков. Столбик выдаётся из венчика, завязь 4-гнёздная, гинецей синкарпный, из 4 плодолистиков, плацентация центрально-угловая (Баландина, Вахрамеева, 1978). Диплоидный набор хромосом равен 24. В природных условиях найдено триплоидное растение с 36 хромосомами (Богданова, Муратов, 1978).
Брусника имеет обширный голарктический ареал с преимущественным распространением в северной части Евразии. Встречается по всей территории СНГ, кроме южных районов европейской части, всей Средней Азии, подавляющей части Казахстана и Закавказья (Атлас ареалов…, 1976).
Брусника растёт в лесной и арктической зонах, поднимается до гольцового пояса, распространена в хвойных и смешанных лесах, на вырубках, в горных и равнинных тундрах. Чаще всего она встречается в травяно-кустарничковом ярусе сосновых и лиственничных лесов, являясь доминантом этого яруса (Брусника…, 1986).
Брусника — медоносное, дубильное, пищевое, лекарственное растение, кормовое для диких животных.
2.2. Место исследования
Исследования проводились в 1999-2002 годах на территории государственного природного заповедника «Большая Кокшага» Республики Марий Эл.
Заповедник расположен в среднем течении реки Большая Кокшага на территории Медведевского и Килемарского административных районов РМЭ на расстоянии около 40 км от г. Йошкар-Олы. Координаты крайних точек территории заповедника: северной — 56º64' с.ш., западной — 49º09' в.д., восточной — 47º26' в.д. (Летопись природы, 1998).
Территория заповедника, согласно физико-географическому районированию СССР и Среднего Поволжья, расположена в пределах лесной зоны Русской равнины подзоны хвойно-широколиственных лесов Ветлужско-Унженской географической провинции Ветлужско-Кокшагского полесского района Оршано-Кокшагской флювиогляциальной равнины. По ботанико-географическому районированию Марий Эл (Абрамов, 1989) она входит в Ветлужско-Юшутский район.
Согласно климатическому районированию СССР заповедник находится в умеренном климатическом поясе атлантико-континентальной области. В пределах Марий Эл он расположен в центральном агроклиматическом районе (Агроклиматические ресурсы Марийской АССР, 1972).
Климат умеренно-континентальный, характеризующийся морозной зимой и умеренно-жарким летом. Приход солнечной радиации составляет около 90 ккал/см². Основная часть тепла при этом расходуется на испарение воды с поверхности почвы и на транспирацию растением.
Средняя продолжительность тёплого периода года (с температурой воздуха выше 0ºС) составляет 201 день. Устойчивый переход через 0ºC, являющийся признаком начала и конца зимы, наблюдается в среднем 26 октября (осень) — 7 апреля (весна). Зима продолжается 164 дня, а наиболее морозная её часть с температурой ниже -10ºС — 89 дней.
Активная вегетация растений начинается в среднем 9 мая и заканчивается 15 сентября, продолжаясь 128 дней. Наиболее тёплая часть лета продолжается всего 77 дней (с 6 июня по 22 августа).
За год в среднем выпадает 538 мм осадков, из которых подавляющая часть (387 мм) приходится на тёплый период.
На территории заповедника сформировались дерново-подзолистые почвы различного механического состава. Преобладающими являются слабоподзолистые песчаные почвы.
Основными ландшафтами заповедника являются дюнно-бугристые и пологоволнистые зандровые равнины, покрытые в основном сосняками и березняками, а также пойменные урочища, занятые вдоль р. Большая Кокшага дубово-липовыми насаждениями, а вдоль малых рек и ручьёв — берёзовыми или черноольховыми с небольшой примесью ели (Летопись природы, 1998).
Для исследования были заложены 4 постоянные пробные площади; проведены геоботанические описания данных местообитаний.
Пробная площадь №1 — сосняк. Квартал 65, выдел 33. В 1995 году здесь прошёл низовой пожар. Расположен на первой надпойменной террасе реки Большая Кокшага. Возобновление отсутствует. Подлесок отсутствует. В травяно-кустарничковом ярусе преобладает брусника (4,2%).
Пробная площадь №2 — сосняк брусничный. Находится в охранной зоне заповедника. В 1972 г. здесь прошёл пожар, который полностью уничтожил травяно-кустарничковый ярус. Возобновление отсутствует. В подлеске — рябина обыкновенная Sorbus aucuparia L., ель обыкновенная Picea abies L. Сейчас в травяно-кустарничковом ярусе преобладает брусника (34,8 %), лишайники (различные виды кладоний — лесная, альпийская, оленья и др., 9,9%), марьянник луговой Melampyrum pratensis L. (8,5%), зелёные мхи (плевроциум Шребера, кукушкин лён обыкновенный, дикранум многоножковый, 6,6%).
Пробная площадь №3 — сосняк зеленомошно-брусничный. Квартал 90, выдел 24. Расположен на первой надпойменной террасе реки Большая Кокшага. В данном фитоценозе был пожар в 1921 году. Возобновление отсутствует. В подлеске — можжевельник обыкновенный Juniperus communis L. В травяно-кустарничковом ярусе преобладают зелёные мхи (плевроциум Шребера, кукушкин лён обыкновенный, дикранум многоножковый, 59,7%), брусника (16,9%).
Пробная площадь №4 — сосняк зеленомошно-брусничный. Квартал 65, выдел 33. Расположен на первой надпойменной террасе реки Большая Кокшага. Представляет собой посадки начала XX века после пожара. Возобновление отсутствует. В подлеске — ель обыкновенная. В травяно-кустарничковом ярусе преобладают зелёные мхи (плевроциум Шребера, кукушкин лён обыкновенный, дикранум многоножковый, 51,3%), брусника (18,1%).
Таксационные описания изученных пробных площадей представлены в таблице 1.
Таблица 1
Таксационные описания пробных площадей
| № пробной площади | Состояние древостоя | Средний возраст | Класс возраста | Бонитет | Полнота | Сомкнутость |
| 1 | 10С+Ед.Б | 85 | 5 | 2 | 0,8 | 0,7 |
| 2 | 9С1Б | 66 | - | 1 | 0,5 | - |
| 3 | 10С+Б | 45 | 3 | 3 | 0,8 | 0,7 |
| 4 | 10С+Ед.Б | 85 | 5 | 2 | 0,8 | 0,7 |
2.3. Методы исследований
Изучение ценопопуляций брусники проводили в 1999-2002 годах в Государственном природном заповеднике «Большая Кокшага» Республики Марий Эл.
Исследования проводили в четырёх различных ассоциациях. Геоботанические описания обработаны по экологическим шкалам Д.Н.Цыганова (1983) с помощью компьютерной программы «Ecoscale». Исследованные местообитания сравнили по флористическому составу с помощью коэффициента Жаккара:
![]()
где a — число видов в первой ценопопуляции,
b — число видов во второй ценопопуляции,
с — число общих видов (Лакин, 1990).
В каждой ассоциации было заложено по 30 учётных площадок размером 1 м², в пределах которых в 1999 году были замаркированы парциальные кусты брусники различных возрастных состояний (имматурного, виргинильного, молодого генеративного, средневозрастного генеративного, старого генеративного, субсенильного, сенильного и отмирающего).
Возрастные состояния парциальных кустов брусники выделяли согласно методике, предложенной А.А.Урановым с дополнениями. Диагнозы возрастных состояний для брусники обыкновенной были приведены Л.В.Прокопьевой с соавторами (2000).
ПРОРОСТКИ брусники однопобеговые, с двумя несколько удлинёнными семядолями и 2-8 ассимилирующими листьями. Семядольные листья сверху тёмно-зелёные, блестящие, снизу более светлые, длиной 2-2,5 мм и шириной 1 мм. Ассимилирующие листья 1,5-2,0 мм длиной и около 1,0 мм шириной, средняя жилка у них не выражена совсем или очень слабо. Корневая система представлена главным и немногочисленными боковыми корнями II порядка.
ИММАТУРНЫЕ растения представлены одним моноподиально нарастающим побегом или побегом I порядка с 1-2 побегами II порядка. Стебель коричневатый, опушённый. Длительность состояния—1-3 года. Корневая система представлена многочисленными придаточными корнями рыжего цвета.
У ВИРГИНИЛЬНЫХ растений число побегов II порядка 1-4. Эти растения формируют побеги II и IIIпорядков, образуя парциальный куст. Отмирающие побеги отсутствуют. Стебель опушён, коричневого или тёмно-зелёного цвета. Корневая система густая, представлена придаточными корнями рыжего цвета.
МОЛОДЫЕ ГЕНЕРАТИВНЫЕ парциальные кусты формируют побеги II, III и редко IV порядков. Побег I порядка одревесневший. Отмерших побегов не более 10%. Парциальные кусты начинают цвести и плодоносить, после чего побеги нарастают симподиально. Возраст листьев обычно 3 (2-4) года, они обычной формы, длина их достигает 1,8-2,0 (редко до 2,9) см, ширина 0,8-1,1 см. Корневая система приобретает бурый оттенок.
![]() v
-
парциальные кусты имеют все вышеперечисленные признаки, но не цветут и не плодоносят. От виргинильных растений они будут отличаться одревесневшим побегом I порядка. Возраст парциальных кустов 2-5 лет.
v
-
парциальные кусты имеют все вышеперечисленные признаки, но не цветут и не плодоносят. От виргинильных растений они будут отличаться одревесневшим побегом I порядка. Возраст парциальных кустов 2-5 лет.
СРЕДНЕВОЗРАСТНЫЕ ГЕНЕРАТИВНЫЕ парциальные кусты имеют побеги II, III, IV и редко V порядков. Побеги I порядка, нижние части побегов II порядка одревесневшие. Появляются побеги замещения. Отмирание побегов не более 10-40 %. Растение обильно цветёт и плодоносит. Листья двух-, трёхлетние, их длина 2,0-2,5 см, ширина 0,8-1,3 см. Корневая система бурого цвета. Возраст парциальных кустов 4-8 лет.
![]() v
-растения по всем признакам сходны со средневозрастными генеративными парциальными кустами, но не цветут и не плодоносят.
v
-растения по всем признакам сходны со средневозрастными генеративными парциальными кустами, но не цветут и не плодоносят.
СТАРЫЕ ГЕНЕРАТИВНЫЕ парциальные кусты формируют побеги VI и более порядков, цветут и плодоносят уже менее интенсивно, чем средневозрастные генеративные растения. Побеги I и II порядков одревесневшие, чёрного цвета. Отмершие побеги составляют 40-60%. Корневая система начинает отмирать, она бурого или тёмно-коричневого цвета, в незначительном объёме.
![]() v
-
растения не цветут и не плодоносят, но имеют особенности старых генеративных парциальных кустов по степени отмирания и новообразования и др. Возраст парциальных кустов 4-8 лет.
v
-
растения не цветут и не плодоносят, но имеют особенности старых генеративных парциальных кустов по степени отмирания и новообразования и др. Возраст парциальных кустов 4-8 лет.
![]() v
-,
v
-,
![]() v
-
и
v
-
и![]() v
-
парциальные кусты брусники составляют довольно значительную группу, они во много раз превосходят по численности генеративные растения и играют значительную роль в жизни ценопопуляций этого вида, поскольку позволяют удерживать занимаемую видом территорию, а также способствуют вегетативному разрастанию особи и обеспечивают самоподдержание ценопопуляций.
v
-
парциальные кусты брусники составляют довольно значительную группу, они во много раз превосходят по численности генеративные растения и играют значительную роль в жизни ценопопуляций этого вида, поскольку позволяют удерживать занимаемую видом территорию, а также способствуют вегетативному разрастанию особи и обеспечивают самоподдержание ценопопуляций.
СУБСЕНИЛЬНЫЕ ПК имеют побеги II-VI и более высоких порядков. Не цветут и не плодоносят. Степень отмирания более 60%. Корневая система слабая, чёрного цвета. Число листьев не более 10-14, сохраняются не более 2 лет. Побеги I-III порядков одревесневают. Растениям 5 и более лет.
СЕНИЛЬНЫЕ ПК имеют всего один побег с 1-5 листьями. Приростов нынешнего года нет. Доля отмерших побегов составляет 80-99%. Продолжительность состояния 5 и более лет.
ОТМИРАЮЩИЕ растения представлены системой скелетных осей тёмного цвета. Листьев обычно нет или 1-2 засохших. Корневая система отмирает или она чёрного цвета.
Наблюдения проводили за примерно 2300 парциальными кустами брусники.
В течение четырёх лет определялись возрастные состояния каждого замаркированного парциального куста брусники. Были составлены схемы возможных путей развития парциальных кустов отдельно для каждого исходного возрастного состояния (Приложение).
Далее проводился анализ схем возможных путей развития по нескольким направлениям:
1) динамика выживания парциальных кустов брусники;
2) доля парциальных кустов брусники с пропусками возрастных состояний;
3) возрастные спектры последнего года исследования для особей разных возрастных состояний на старте;
4) продолжительность возрастных состояний.
Полученные данные были проанализированы с помощью критерияχ² (таблицы сопряжённости r × c ) (Глотов и др., 1982).
3. РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
3.1. Экологическая характеристика местообитаний
Характеристики изученных местообитаний по шести экологическим шкалам Д.Н.Цыганова (1983) — увлажнение (Hd), богатство почв (Tr), богатство азотом (Nt), кислотность (Rc), освещённость (Lc) и переменность увлажнения (fH) — представлены в таблице 2.
Рассматриваемые ценопопуляции находятся в довольно сходных экологических условиях. Для всех ценопопуляций характерны бедные азотом почвы (шкала Nt).По шкале освещённости (Lc) все ценопопуляции можно классифицировать как светлые леса. Однако наблюдаются и некоторые различия. Так, для сосняка (пробная площадь 1) и сосняка брусничного (пробная площадь 3) характерны низкие показатели по шкале увлажнения почв (Hd), в связи с чем они классифицируются как влажнолесолуговые, в то время как два других местообитания — переходные от влажнолесолуговых к сыролесолуговым. Изученные местообитания отличаются слабокислыми, переходными от небогатых к довольно богатым почвами. Для пробной площади 1 характерно увлажнение переходное от слабо переменного к умеренно переменному. Остальные местообитания характеризуются слабо переменным увлажнением.
Таким образом, исследованные местообитания являются достаточно сходными в отношении экологических условий с небольшими отличиями.
Таблица 2
Характеристика местообитаний
по шкалам Д.Н. Цыганова (1983)
№ пробной площади |
Hd Увлажне-ние |
Tr Богатство почв |
Nt Богатство почв азотом |
Rc Кислотность |
Lc Освещён-ность |
fH Перемен- ность увлажне- ния |
| 1 | 12,68 влажноле- солуговое |
5,29 небогатые/ довольно богатые |
4,95 бедные азотом |
6,16 слабо кислые |
4,29 светлые леса |
5,21 слабо переменное/умеренно переменное |
| 2 | 13,13 влажноле- солуговое/ сыролесо- луговое |
4,88 небогатые |
4,27 бедные азотом |
5,32 кислые/ слабокислые |
4,55 светлые леса |
4,65 слабо переменное |
| 3 | 12,82 влажноле- солуговое |
5,02 небогатые/ довольно богатые |
4,82 бедные азотом |
6,06 слабокислые |
4,45 светлые леса |
4,59 слабо переменное |
| 4 | 13,24 влажноле- солуговое/ сыролесо- луговое |
5,35 небогатые/ довольно богатые |
5,00 бедные азотом |
6,06 слабокислые |
4,53 светлые леса |
4,80 слабо переменное |
По данным таблицы 2 можно построить экологическое пространство для исследованных ценопопуляций брусники, а также оценить долю экологического пространства и построить полиграмму экологического ареала и экологического пространства (таблица 3, рис. 1).
Таблица 3
Экологические характеристики ЦП брусники
в ГПЗ «Большая Кокшага»
Экологические шкалы |
Диапазон экологического ареала брусники по Д.Н.Цыганову |
Доля диапазона экологического ареала брусники, % | Экологическое пространство изучаемых ЦП |
Доля экологического пространства изучаемых ЦП, % |
| Hd (1-23) | 10-17 | 34,8 | 12,7-13,2 | 7,0 |
| Tr (1-19) | 1-7 | 36,8 | 4,9-5,5 | 6,7 |
| Nt (1-11) | 1-5 | 45,5 | 4,3-5,0 | 14,6 |
| Rc (1-13) | 1-6 | 46,2 | 5,3-6,2 | 14,0 |
| Lc (1-9) | 1-8 | 88,9 | 4,3-4,6 | 3,3 |
| fH (1-11) | 1-5 | 45,5 | 4,7-5,2 | 11,2 |
Как свидетельствуют полученные данные, брусника имеет достаточно широкий экологический ареал по фактору освещённости — 88,9%. Самые низкие значения брусника имеет по шкалам увлажнения и солевого режима почв, где экологический ареал вида занимает всего около 35% от возможного диапазона шкалы.
Изученные ценопопуляции отличаются сравнительно узким диапазоном экологического пространства по всем экологическим шкалам (3,3-14,6%).
Кроме того, следует отметить, что по таким факторам, как кислотность и переменность увлажнения, изученные ценопопуляции несколько выходят за границы ареала брусники, предложенного Д.Н.Цыгановым (1983) (рис. 1).
Диапазон экологических шкал
Экологический ареал брусники по Д.Н.Цыганову (1983)
Экологическое пространство изученных ценопопуляций брусники
Рис. 1. Полиграмма экологического ареала и экологического пространства ценопопуляций брусники в ГПЗ «Большая Кокшага»
Исследованные местообитания сравнили по флористическому составу при помощи коэффициента общности Жаккара (Лакин, 1990). Полученные данные представлены в таблице 4.
Таблица 4
Флористическое сходство пробных площадей
(коэффициент общности Жаккара, %)
| № пробной площади | 1 | 2 | 3 |
| 2 | 23,3 | __ | __ |
| 3 | 25,8 | 37,1 | __ |
| 4 | 33,3 | 28,6 | 31,0 |
Все пробные площади достаточно сильно отличаются друг от друга по видовому составу, коэффициент общности не превышает 40% для всех сравниваемых пар.
Таким образом, рассматриваемые местообитания характеризуются близкими значениями по экологическим шкалам, но довольно различны по видовому составу, что, по-видимому, связано с какими-то неучтёнными факторами (влияние пожаров в разное время, разное антропогенное воздействие и т.п.).
3.2. Темпы развития парциальных кустов брусники
3.2.1. Выживаемость парциальных кустов
В исследованных ценопопуляциях проводили изучение темпов развития парциальных кустов брусники путём длительных наблюдений за маркированными парциальными кустами и анализа полученных данных.
На рисунке 2 показаны частоты выживаемости парциальных кустов за весь период наблюдения.
Полученные данные были обработаны с помощью критерия хи-квадрат (χ²). Установлено, что ценопопуляции по выживаемости парциальных кустов всех возрастных состояний имеют значимые различия (P<0,001). При этом максимальная смертность во всех случаях отмечена в ЦП 2 (15-45% выживших ПК от исходного количества).
В ряде случаев (для парциальных кустов имматурного, средневозрастного генеративного и старого генеративного онтогенетических состояний) низкая выживаемость наблюдалась и в ЦП 4 (36-63% выживших ПК от исходного количества). По-видимому, в начале своего онтогенеза (im ) и в период максимального развития (g 2 и g 3 ) парциальные кусты брусники данного местообитания испытывают влияние каких-то резких факторов среды, что сказывается на их выживании.
Минимальная смертность парциальных кустов во всех возрастных группах наблюдалась в ценопопуляциях 1 и 3, между ними нет статистически значимых различий (P>0,1).
Высокую смертность парциальных кустов брусники в ценопопуляциях 2 и 4 можно объяснить тем, что данные ценопопуляции характеризуются наибольшей плотностью парциальных кустов — 169,5 и 102,0 экз./м² соответственно. Ценопопуляции 1 и 3 отличаются низкой плотностью—48,8 и 46,6 экз./м² (Алметева, 2003).
а) для парциальных кустов имматурного состояния
![]()
б) для парциальных кустов виргинильного состояния
![]()
в) для парциальных кустов молодого генеративного состояния
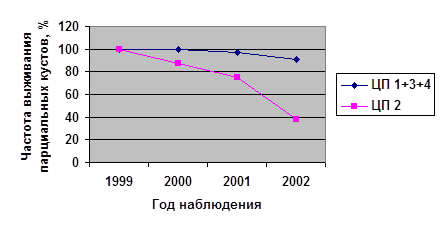
Рис. 2. Динамика выживания парциальных кустов брусники
г) для парциальных кустов средневозрастного генеративного состояния
![]()
д) для парциальных кустов старого генеративного состояния
![]()
Рис. 2 (продолжение). Динамика выживания парциальных кустов брусники
3.2.2. Пропуск возрастных состояний
В исследованных ценопопуляциях определялась доля парциальных кустов, в ходе своего онтогенеза перешедших из одного возрастного состояния в другое с пропуском одного или нескольких или целого периода. Зарегистрированы случаи пропусков виргинильного, молодого, средневозрастного и старого генеративного, субсенильного, сенильного и отмирающего состояний. Максимальное число пропущенных возрастных состояний — 6 (переходы im — sc ). Результаты представлены в таблице 5.
Таблица 5
Доля парциальных кустов брусники, первоначально зарегистрированных в определённом возрастном состоянии и пропустивших хотя бы одно возрастное состояние, %
№ ЦП |
Возрастное состояние | Всего |
|||||||
| im | v | g1 | g2 | g3 | ss | s | sc | ||
| 1 | 89,4 | 90,8 | 28,4 | 34,2 | 63,2 | 80,0 | - | 0 | 64,6 |
| 2 | 98,2 | 93,4 | 82,2 | 82,9 | 93,5 | 84,6 | 50,0 | 0 | 86,3 |
| 3 | 88,2 | 70,3 | 38,4 | 40,2 | 44,3 | 54,5 | - | 0 | 52,9 |
| 4 | 95,9 | 81,5 | 38,0 | 60,5 | 70,9 | 78,4 | 14,3 | 0 | 62,1 |
Пути развития парциальных кустов брусники с пропуском одного или нескольких возрастных состояний, обнаружены во всех исследованных ценопопуляциях, однако доля таких парциальных кустов в разных ценопопуляциях различна. Максимальная частота обнаружена в сосняке брусничном (ЦП 1) — 64,6%, минимальная — в сосняке зеленомошно-брусничном (ЦП 3) — 52,9%. Таким образом, путь онтогенеза парциальных кустов брусники в более чем половине случаев происходит с пропусками одного или нескольких возрастных состояний. Это может быть объяснено двумя причинами. Во-первых, наблюдения за маркированными кустами велись лишь один раз в сезон, поэтому возможно, что мы не заметили «правильные» (или поочерёдные) переходы. И во-вторых, данные пропуски могут быть связаны с тем, что для этого кустарничка нормальным ходом онтогенеза будут именно пропуски онтогенетических состояний.
Доля парциальных кустов брусники, характеризующихся такими темпами онтогенетического развития парциальных кустов, при которых развитие происходит последовательно без пропусков возрастных состояний, приведена в таблице 6.
Таблица 6
Доля парциальных кустов брусники без пропусков онтогенетических состояний, % от общего числа ПК данного возрастного состояния
| Путь онтогенеза | № ценопопуляции | |||
| 1 | 2 | 3 | 4 | |
| im—v—g1—g2 | 9,7 | 1,8 | 10,0 | 3,1 |
| v—g1—g2—g3 | 0 | 1,6 | 4,7 | 4,4 |
| g1—g2—g3—ss | 3,9 | 0,9 | 4,4 | 1,4 |
| g2—g3—ss—s | 2,6 | 0 | 0,4 | 0,6 |
| g3—ss—s—sc | 0 | 0 | 4,9 | 1,2 |
| ss—s—sc | 0 | 0 | 18,2 | 10,8 |
Полученные данные были обработаны с помощью критерия хи-квадрат (χ²). Результаты свидетельствуют о том, что частоты парциальных кустов брусники, развивающихся с последовательной сменой возрастных состояний, однородны для всех изученных ценопопуляций (P>0,1) и составляют 52,9-86,3%.
Следовательно, парциальные кусты брусники во всех изученных ценопопуляциях развиваются преимущественно по пути с пропусками возрастных состояний, и лишь для незначительной доли их характерно последовательное онтогенетическое развитие без пропусков отдельных этапов онтогенеза, причём это утверждение справедливо для всех возрастных групп.
3.2.3. Продолжительность возрастных состояний
В исследованных ценопопуляциях измерялась продолжительность возрастных состояний парциальных кустов брусники и определялось число парциальных кустов брусники, находящихся в данном онтогенетическом состоянии более одного года. Данные представлены в таблицах 7-11.
Полученные данные были проанализированы с помощью критерия хи-квадрат (χ²). Обнаружено, что продолжительность старого генеративного онтогенетического состояния одинакова во всех изученных ценопопуляциях (P>0,1). Однако ценопопуляции различаются по длительности всех остальных рассмотренных онтогенетических состояний (P<0,001). По-видимому, на продолжительность имматурного, виргинильного, молодого генеративного и средневозрастного генеративного состояний влияют какие-то неучтённые эколого-ценотические условия данных местообитаний, которые не сказываются на длительности старого генеративного возрастного состояния.
Таблица 7
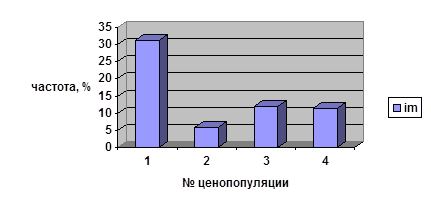
Число имматурных парциальных кустов, находящихся в данном возрастном состоянии более 1 года
№ ценопопуляции |
Общее число ПК |
Частота ПК, находящихся в данном возрастном состоянии более 1 года, % |
| 1 | 100 | 31,0 |
| 2 | 50 | 6,0 |
| 3 | 100 | 12,0 |
| 4 | 62 | 11,3 |
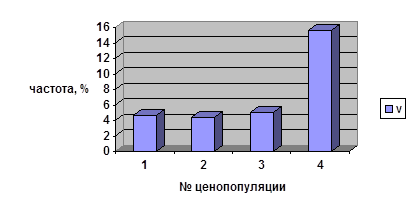
Таблица 8
Число виргинильных парциальных кустов, находящихся в данном
возрастном состоянии более 1 года
№ ценопопуляции |
Общее число ПК |
Частота ПК, находящихся в данном возрастном состоянии более 1 года, % |
| 1 | 64 | 4,7 |
| 2 | 22 | 4,4 |
| 3 | 120 | 5,0 |
| 4 | 128 | 15,6 |
Таблица 9
Число молодых генеративных парциальных кустов, находящихся в данном возрастном состоянии более 1 года
№ ценопопуляции |
Общее число ПК |
Частота ПК, находящихся в данном возрастном состоянии более 1 года, % |
| 1 | 92 | 4,3 |
| 2 | 45 | 15,6 |
| 3 | 143 | 35,7 |
| 4 | 193 | 48,2 |
Таблица 10
Число средневозрастных генеративных парциальных кустов, находящихся в данном возрастном состоянии более 1 года
№ ценопопуляции |
Общее число ПК |
Частота ПК, находящихся в данном возрастном состоянии более 1 года, % |
| 1 | 32 | 68,4 |
| 2 | 26 | 80,8 |
| 3 | 195 | 89,7 |
| 4 | 97 | 61,9 |
Таблица 11
Число старых генеративных парциальных кустов, находящихся в данном возрастном состоянии более 1 года
№ ценопопуляции |
Общее число ПК |
Частота ПК, находящихся в данном возрастном состоянии более 1 года, % |
| 1 | 10 | 90,0 |
| 2 | 7 | 57,1 |
| 3 | 46 | 78,3 |
| 4 | 31 | 87,1 |
Исходя из данных таблиц 7-11, были построены диаграммы частот парциальных кустов брусники, находящихся в данном возрастном состоянии более одного года для каждого этапа онтогенеза (рис. 3).
Частоты, характеризующие длительность всех этапов онтогенеза кроме старого генеративного возрастного состояния, сильно варьируют во всех ценопопуляциях брусники. Так, максимальное количество имматурных парциальных кустов, сохранивших своё онтогенетическое состояние более одного года, отмечено в ЦП 1 (31%), что может быть объяснено недавним пожаром (в 1993 г.) в данном местообитании. Для остальных ценопопуляций наблюдаются близкие значения частот нахождения в данном возрастном состоянии в пределах этой возрастной группы (6-12%). В возрастной группе виргинильных парциальных кустов максимум приходится на ценопопуляцию 4 (15,4%). Эта ценопопуляция сформировалась в данном ценозе очень давно — более 80 лет назад.
а) для парциальных кустов имматурного состояния
б) для парциальных кустов виргинильного состояния
в) для парциальных кустов молодого генеративного состояния
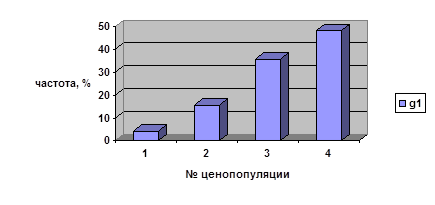
Рис. 3. Частота парциальных кустов брусники, находящихся в данном возрастном состоянии более одного года
г) для парциальных кустов средневозрастного генеративного состояния
![]()
д) для парциальных кустов старого генеративного состояния
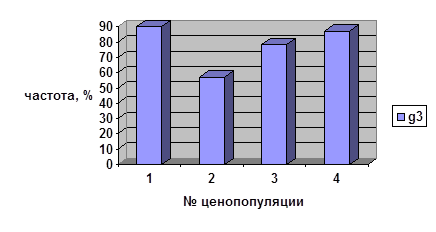
Рис. 3 (продолжение). Частота парциальных кустов брусники, находящихся в данном возрастном состоянии более одного года
Три другие ценопопуляции характеризуются достаточно близкими значениями частот (4,4-5,0%). Что касается молодых генеративных и средневозрастных генеративных парциальных кустов, то в этих случаях отмечаются значительные колебания частот парциальных кустов, находящихся в данном возрастном состоянии более одного года, во всех четырёх ценопопуляциях. Как свидетельствуют данные рисунка 3, продолжительность онтогенетического состояния старых генеративных парциальных кустов характеризуется близкими значениями частот во всех изученных ценопопуляциях — от 57 до 90%.
Длительность возрастных состояний, по-видимому, определяется неучтёнными факторами среды, такими как возраст древостоя, возраст ценопопуляции, давность пожара, плотность парциальных кустов, влияние других видов и т.п., которые не сказываются на продолжительности только старого генеративного состояния.
3.2.4. Итоговые возрастные спектры
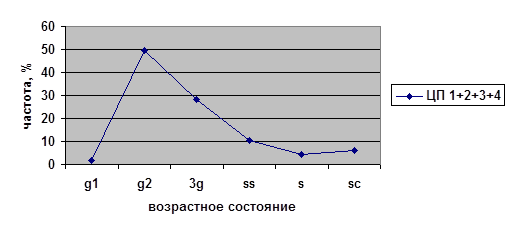
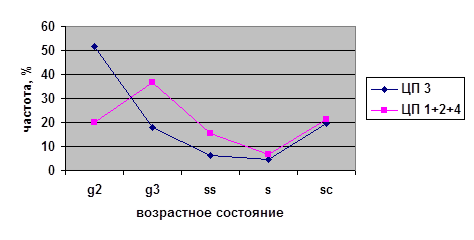
В исследованных ценопопуляциях проводилось изучение возрастных спектров последнего года исследований (2002) для парциальных кустов разных возрастных состояний на старте (рис. 4). Полученные данные были проанализированы с помощью критерия хи-квадрат (χ²). Виргинильные, молодые генеративные и средневозрастные генеративные парциальные кусты из разных ценопопуляций на четвёртый год наблюдений дают разные возрастные спектры. При этом не наблюдается статистически значимых различий между ЦП 1, 3 и 4 в случае виргинильных парциальных кустов; между ЦП 2, 3 и 4 в случае молодых генеративных парциальных кустов и между ЦП 1, 2 и 4 в случае средневозрастных генеративных парциальных кустов. Возрастные спектры имматурных и старых генеративных ПК из разных ценопопуляций не различаются.
а) итоговый возрастной спектр для имматурных парциальных кустов брусники
б) итоговые возрастные спектры для виргинильных парциальных кустов брусники
![]()
в) итоговые возрастные спектры для молодых генеративных парциальных кустов брусники
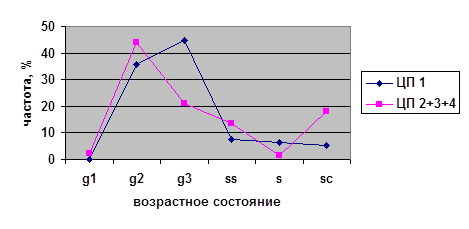
Рис. 4. Итоговые возрастные спектры
г) итоговые возрастные спектры для средневозрастных генеративных парциальных кустов брусники
д) итоговый возрастной спектр для старых генеративных парциальных кустов брусники
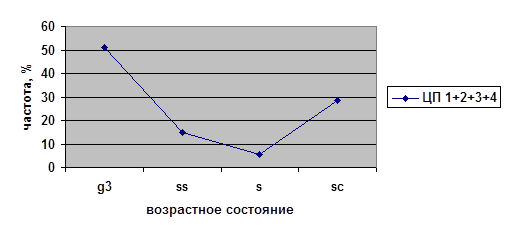
Рис. 4 (продолжение). Итоговые возрастные спектры
Таким образом, темпы развития парциальных кустов брусники являются довольно динамичными в разных ценопопуляциях. Парциальные кусты различных возрастных состояний ведут себя не одинаково в разных условиях. Причём, чаще всего это касается парциальных кустов генеративного периода — молодых генеративных и средневозрастных генераитвных, а также виргинильных парциальных кустов. На начальных этапах (im) и в конце (g3, ss) онтогенеза различий экологических условий не наблюдается.
ВЫВОДЫ
1. По экологическим шкалам Д.Н. Цыганова местообитания брусники находятся в сходных экологических условиях по факторам увлажнения и переменности увлажнения, освещённости, кислотности, солевого режима и богатства почв азотом, но в то же время значительно различаются по флористическому составу (коэффициенты Жаккара не превышают 40%).
2. Изученные ценопопуляции различаются по выживаемости парциальных кустов брусники всех возрастных состояний, причём большая смертность отмечена в ценопопуляциях 2 и 4, характеризующихся высокой плотностью парциальных кустов брусники.
3. Парциальные кусты брусники всех изученных ценопопуляций имеют различный ход онтогенеза. Наиболее часто встречается ход с пропусками одного или нескольких возрастных состояний. Частоты таких парциальных кустов составляют 52-86%.
4. Виргинильные, молодые генеративные и средневозрастные генеративные парциальные кусты брусники из разных ценопопуляций на четвёртый год наблюдений дают разные возрастные спектры; возрастные спектры имматурных и старых генеративных парциальных кустов из разных ценопопуляций не различаются.
5. Продолжительность онтогенетических состояний одинакова во всех изученных ценопопуляциях для старых генеративных парциальных кустов брусники и различается для других возрастных состояний.
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
1. Абрамов Н.В. Сосудистые растения флоры МАССР: учеб. пособие.—Йошкар-Ола: Марийский ун-т, 1989.—147 с.
2. Абрамов Н.В. Конспект флоры республики Марий Эл.—Йошкар-Ола, 1995.—191 с.
3. Абрамов Н.В. Флора Республики Марий Эл.—Йошкар-Ола, 2000.—163 с.
4. Агроклиматические ресурсы Марийской АССР.—Л.: Гидрометеоиздат, 1972.—107 с.
5. Алметева Н.В.
6. Атлас ареалов и ресурсов лекарственных растений СССР.—М.: ГУГК, 1976.—340 с.
7. Баландина Т.П., Вахрамеева М.Г. Брусника обыкновенная // Биол. флора Моск. обл. Вып. 4—М., 1978.—С.167-178.
8. Богданова Г.А., Муратов Ю.М. Брусника в лесах Сибири.—Новосибирск: Наука, 1978.—178 с.
9. Боч М.С., Смагин В.А. Флора и растительность болот Северо-запада России и принципы их охраны.—С-Пб.: Гидрометеоиздат, 1993.—224 с.
10. Брусника: Морфология и анатомия. Фитоценотическая приуроченность. Урожайность. Хранение и переработка. Химический состав ягод/Юдина В.Ф., Белоногова Т.В., Колупаева С.Н. и др.—М., 1986.—73 с.
11. Булгаков Н.К., Козьяков С.Н., Фесюк А.В. Технология заготовки и переработки недревесных ресурсов леса.—М.: Лесн. Пром-ть, 1987.—С. 33-34.
12. Бумар Г.И., Таргонский П.Н. Возрастной состав ценопопуляций Vaccinium vitis - idaea L. в Полесском государственном заповеднике (УССР)//Растительные ресурсы.—1989.—Т.25, вып.3.—с.349-353.
13. Викторов С.В., Ремезова Г.Л. Индикационная геоботаника.—М., 1988.—168 с.
14. Воронцова Л.И. Изменение жизненного состояния эдификаторов растительного покрова южной полупустыни под влиянием экологических условий // Онтогенез и возрастной состав популяций цветковых растений.—М.: Наука, 1967.—155 с.
15. Воронцова Л.И., Заугольнова Л.Б. Мультивариантность развития особей в течение онтогенеза и её значение в регуляции численности и состава ценопопуляций растений // Журн. общ. биологии, №4, 1978.—С. 555-562.
16. Гатцук Л.Е. Морфогенез копеечника кустарникового (Hedysarum fruticosum Pall.) при постоянном уровне песчаного субстрата // Онтогенез и возрастной состав популяций цветковых растений.—М.: Наука, 1967.—155 с.
17.
18. Давидюк Л.К. Динамика численности и возрастного состава ценопопуляций представителей травяного покрова пойменных дубрав // Структура и динамика растительных сообществ Волго-Вятского региона.—Горький: Горьковский ун-т, 1987.—87 с.
19. Егошина Т.Л. Продуктивность листьев брусники в фитоценозах средней тайги // Продуктивность таёжных биогеоценозов. Тезисы докладов краевой научной конференции 15-17 апреля 1986 г., г. Красноярск.—Красноярск: ИЛиД, 1986.—С.56.
20. Елина Г.А. и др. Структурно-функциональная организация и динамика болотных экосистем Карелии/ отв. Ред. Н.И. Пьявченко.—Л.: Наука, 1984.—128 с.
21. Ермакова И.М., Жукова Л.А. Типы функционирования ежи сборной и овсяницы луговой в луговых агрофитоценозах // Динамика ценопопуляций растений.—М.: Наука, 1985.—С.110-126.
22. Жукова Л.А. Популяционная жизнь луговых растений.—Йошкар-Ола, 1995.—224 с.
23. Жукова Л.А., Комаров А.С. Количественный анализ динамической поливариантности в ценопопуляциях подорожника большого при разной плотности посадок // Биол. науки, №8, 1991.—С. 51-66.
24. Жукова Л.А., Шейпак О.А. Влияние гербицидов на возрастной состав ценопопуляций подорожника большого//Воздействие гербицидов на растения на организменном и популяционном уровнях.—М.: МГПИ им. В.И. Ленина, 1985, с. 3-25.
25. Завадский К.М. Вид и видообразование. Л.: Наука, 1968.—404 с.
26. Закамская Е.С., Панова Г.М. Возрастная структура ценопопуляций Rubus saxatilis L. в различных экологических условиях // Жизнь популяций в гетерогенной среде. Ч. 1.—Йошкар-Ола, 1998.—С. 221.
27. Зворыкина К.В. Некоторые особенности структуры травяно-кустарничкового яруса сосняков // Продуктивность и структура растительности молодых сосняков.—М.: Наука, 1973.—С. 87-110.
28. Иванова Р.Р., Ефремова Л.П. Состояние популяций константных видов растений в живом напочвенном покрове различных типов сосновых лесов // Жизнь популяций в гетерогенной среде. Ч. 1.—Йошкар-Ола: Педагогика Марий Эл, 1998.—С. 229-231.
29. Изучение структуры и взаимоотношения ценопопуляций. Метод. Разработки для студентов биол. специальностей/ под ред. Т.И.Серебряковой и др.—М.: Моск. Гос. Пед. ин-т, 1986.—73 с.
30. Ильинская С.А. Изучение синузиальной структуры лесных сообществ // Типы лесов Сибири. Вып. 1.—М.: Изд-во АН СССР, 1963.—С. 5-29.
31. Илюшечкина Н.В. Поливариантность онтогенеза Polemonium coeruleum L. и Valeriana officinalis L. в природных и искусственных ценопопуляциях // Жизнь популяций в гетерогенной среде. Ч. 1.—Йошкар-Ола, 1998.—С. 223.
32. Ипатов В.С., Кирикова Л.А. Фитоценология.—С-Пб.: Изд-во С.-Петербургского ун-та, 1997.—316 с.
33. Кормовые растения сенокосов и пастбищ СССР. Том III. Двудольные (Гераниевые – Сложноцветные). Общие выводы и заключения.—М.-Л.: Гос. Изд-во с/х лит-ры, 1956.—С. 210-217.
34. Крылов А.Г. Жизненные формы лесных фитоценозов.—Л.: Наука, 1984.—189 с.
35. Куркин К.А. Системные исследования динамики лугов.—М.: Наука, 1976.—284 с.
36. Лакин Г.Ф. Биометрия: уч. пособие.—4-е изд., перераб. и доп.—М.: Высшая школа, 1990.—351 с.
37. Летопись природы. Кн.5.—Йошкар-Ола, 1998.—213 с.
38. Мазная Е.А. Влияние промышленных выбросов на состояние и структуру ценопопуляций Vaccinium myrtillus L.иVaccinium vitis - idaea L.(Кольский п-ов)// Растительные ресурсы.—2001.—Т.37, вып.3.
39. Миронов К.А. Восстановление черники после пожаров в лесном Среднем Заволжье // Структура и динамика растительных сообществ Волго-Вятского региона.—Горький: Горьковский ун-т, 1987.—87 с.
40. Онтогенетический атлас лекарственных растений. Т.2.—Йошкар-Ола, 2000.—267 с.
41. Палёнова М.М. Эколого-генетический подход к исследованию искусственной популяции клевера ползучего // Популяции растений: принципы организации и проблемы охраны природы.—Йошкар-Ола, 1991.—с. 90.
42. Популяции растений: принципы организации и проблемы охраны природы/ Мар. Гос. Ун-т.—Йошкар-ола, 1991.—136 с.
43. Пояркова А.И. Ericaceae // Флора СССР. Т. 18—М.-Л., 1952.—С. 22-107.
44. Работнов Т.А. Вопросы изучения состава популяций для целей фитоценологии // Проблемы ботаники. Вып. 1, 1950.—548 с.
45. Работнов Т.А. Фитоценология.—М.: Изд-во МГУ, 1978.—383 с.
46. Работнов Т.А. Луговедение.—М.: Изд-во МГУ, 1984.—319 с.
47. Сабинин Д.А. Физиология развития растений.—М.: Изд-во АН СССР, 1963.—196 с.
48. Смелов С.П. Теоретические основы луговодства.—М.: Колос, 1966.—367 с.
49. Смирнова О.В. Структура травяного покрова широколиственных лесов.—М.: Наука, 1987.—205 с.
50. Сокол А.П. Экологические шкалы болотных растений // Взаимосвязи компонентов лесных и болотных экосистем средней тайги Приуралья.—Л., 1980.—С. 230-240.
51. Соколова И.Г. Пространственно-временная структура ценопопуляций черники резервата «Венский лес» // Популяции растений: принципы организации и проблемы охраны природы.—Йошкар-Ола, 1991.—136 с.
52. Солоневич Н.Г. Материалы и эколого-биологическая характеристика болотных трав и кустарников. Вып. 2.—М.-Л.: Изд-во АН СССР, 1956.—С. 307-497.
53. Структура и динамика растительных сообществ Волго-Вятского региона: межвуз. сборник/ Горький: Горьковский ун-т, 1987.—87 с.
54. Тимошок Е.Е. Внутривидовая изменчивость и демографические особенности брусники обыкновенной в пределах западно-сибирского фрагмента ареала // Популяции и сообщества растений: экология, биоразнообразие, мониторинг. Ч. 2.—Кострома, 1996.—С. 156-166.
55. Тимошок Е.Е., Паршина Н.В. Возрастная структура ценопопуляций Vaccinium vitis - idaea L. в Западной Сибири // Растительные ресурсы.—1992.—Т.28, вып.3.—с.1-14.
56. Тимошок Е.Е. Семейство брусничные (Vaccinaceae ) в Западной Сибири (распространение, экология, популяционная биология и охрана): Диссертация…д-ра биол. наук.—Томск, 1998.—С. 92-231.
57. Толмачёв А.И. К истории возникновения и развития темнохвойной тайги.—М.-Л.: Изд-во АН СССР, 1954.
58. Трубина М.Р. Стресс и возрастная структура популяций растений // Жизнь популяций в гетерогенной среде. Ч. 2.—Йошкар-Ола, 1998.—С. 172.
59. Тюрина Е.В. Изменение ритма роста и развития Peucedanum batealense (Redow.) C. Koch. в зависимости от экологических условий и при интродукции // Ритмы развития и продуктивность полезных растений сибирской флоры.—Новосибирск: Наука, 1975.—182 с.
60. Уранов А.А. Жизненное состояние вида в растительном сообществе.—Бюлл. МОИП. Отд. биол. Т. 65, вып. 3, 1960.
61. Ценопопуляции растений (очерки популяционной биологии). Заугольнова Л.Б., Жукова Л.А., Комаров А.С., Смирнова О.В./ отв. ред. Т.И. Серебрякова, Т.Г. Соколова—М.: Наука, 1988.—181 с.
62. Ценопопуляции растений. Развитие и взаимоотношения.—М.: Наука, 1977.—134 с.
63. Цыганов Д.Н. Фитоиндикация экологических режимов в подзоне хвойно-широколиственных лесов.—М., 1983.—197 с.
64. Черёмушкина В.А. Структура популяций Allium glaucum Schrad. // Жизнь популяций в гетерогенной среде. Ч. 1—Йошкар-Ола, 1998.—С. 239.
65. Чистякова А.А. О жизненной форме и вегетативном разрастании липы сердцевидной // Бюлл. МОИП. Отд. биол. Т.83, вып. 2, 1978.—С. 129-137.
66. Чистякова А.А. Биологические особенности вегетативного возобновления основных пород в широколиственных лесах // Лесоведение. №2, 1982.—С.11-17.
67. Шаврина Е.В. Продуктивность и возрастная структура ценопопуляций черники в различных эколого-фитоценотических условиях // Популяции растений: принципы организации и проблемы охраны природы.—Йошкар-Ола, 1991.—136 с.
68. Шенников А.П. Экология растений.—М.: Советская наука, 1950.—375 с.
69. Шмальгаузен И.И. Кибернетические вопросы биологии.—Новосибирск: Наука, 1968.—223 с.
70. Шорина Н.И. Строение ценопопуляций шафрана Шарояна и шафрана доменного в субальпийском и альпийском поясах западного Закавказья // Исследования состава и изменчивости флоры и растительности.—М., 1979.—134 с.
71. Bozenna C. Ecodiagrams of common species of the forest herb layer in the Roztocze National Park // Acta Societans Botanicorum Poloniae.—1986.—Vol. 55 nr 3: P. 429-466.
72. Ritchie J.C. Vaccinium vitis-idaea L. // J. Ecol.—1955.—Vol. 43, №2.—P. 701-708.
БЛАГОДАРНОСТИ
Выражаю глубокую благодарность за неоценимую помощь при выполнении дипломной работы, внимание и доброе отношение своим научным руководителям: доктору биологических наук, профессору Николаю Васильевичу Глотову и преподавателю кафедры ботаники, экологии и физиологии растений Людмиле Валерьяновне Прокопьевой.
Отдельная благодарность директору Государственного природного заповедника «Большая Кокшага» А.И.Попову и сотрудникам Г.А.Богданову, А.В.Полевщикову.
Благодарю весь педагогический коллектив биолого-химического факультета за труд и терпение, а также декана факультета Михаила Гурьевича Григорьева за неизменную поддержку.
ПРИЛОЖЕНИЕ
Таблица 1
Пути онтогенеза парциальных кустов брусники имматурного состояния
| Год | № ценопопуляции | ||||||
| 1999 | 2000 | 2001 | 2002 | 1 | 2 | 3 | 4 |
| Im | погиб | 5 | 34 | 10 | |||
| im | im | погиб | 2 | ||||
| im | im | v | g1v | 1 | 1 | ||
| im | im | v | g2v | 2 | 2 | ||
| im | im | v | g3v | 1 | |||
| im | im | g1v | g1v | 1 | |||
| im | im | g1v | g2v | 9 | 2 | ||
| im | im | g1v | g3v | 2 | 1 | 2 | |
| im | im | g1v | ss | 1 | 1 | ||
| im | im | g2 | g2v | 1 | |||
| im | im | g2v | g2 | 2 | |||
| im | im | g2v | g2v | 5 | 2 | ||
| im | im | g2v | g3v | 4 | 1 | 1 | 2 |
| im | im | g2v | ss | 1 | |||
| im | im | g3v | g3v | 3 | 1 | ||
| im | im | g3v | ss | 1 | |||
| im | im | ss | ss | 1 | |||
| im | im | ss | s | 1 | 1 | ||
| im | v | v | g1v | 1 | |||
| im | v | v | g2v | 1 | 1 | ||
| im | v | g1v | погиб | 1 | |||
| im | v | g1v | g1v | 1 | |||
| im | v | g1v | g2v | 11 | 2 | 11 | 3 |
| im | v | g1v | g3v | 2 | 1 | 1 | 5 |
| im | v | g1v | ss | 1 | |||
| im | v | g1v | s | 1 | |||
| im | v | g1v | sc | 1 | |||
| im | v | g2 | g2 | 2 | |||
| im | v | g2 | g2v | 1 | |||
| im | v | g2v | погиб | 2 | |||
| im | v | g2v | g2v | 4 | 6 | 4 | 3 |
| im | v | g2v | g3v | 3 | 1 | 1 | 1 |
| im | v | g2v | ss | 2 | |||
| im | v | g2v | sc | 1 | |||
| im | v | g3v | g3v | 1 | |||
| im | v | g3v | ss | 1 | |||
| im | v | ss | ss | 1 | |||
| im | v | ss | sc | 1 | |||
| im | v | sc | погиб | 3 | |||
| im | v | sc | ss | 1 | |||
| im | g1 | g1v | погиб | 1 | |||
| im | g1 | g1v | g2v | 2 | |||
| im | g1 | g2 | g2v | 1 | |||
| im | g1 | g2v | g2v | 1 | |||
| im | g1 | sc | sc | 1 | |||
| im | g1v | погиб | 1 | 1 | |||
| im | g1v | g1v | g2v | 4 | 2 | 8 | 3 |
| im | g1v | g1v | g3v | 3 | 2 | ||
| im | g1v | g1v | ss | 1 | |||
| im | g1v | g1v | sc | 1 | |||
| im | g1v | g1v | погиб | 1 | |||
| im | g1v | g2v | погиб | 3 | 2 | ||
| im | g1v | g2v | g2v | 7 | 4 | 12 | 5 |
| im | g1v | g2v | g3v | 1 | 5 | 4 | 7 |
| im | g1v | g2v | ss | 1 | |||
| im | g1v | g2v | s | 1 | |||
| im | g1v | g2v | sc | 1 | 1 | ||
| im | g1v | g2 | g2v | 2 | |||
| im | g1v | g3v | погиб | 2 | 1 | 1 | |
| im | g1v | g3v | g3 | 1 | |||
| im | g1v | g3v | g3v | 2 | 6 | 2 | |
| im | g1v | g3v | ss | 1 | |||
| im | g1v | g3v | s | 2 | |||
| im | g1v | g3v | sc | 1 | |||
| im | g1v | g3 | g3v | 1 | |||
| im | g1v | ss | погиб | 1 | 1 | ||
| im | g1v | ss | ss | 1 | 2 | ||
| im | g1v | ss | s | 1 | 1 | ||
| im | g1v | ss | sc | 1 | |||
| im | g1v | s | погиб | 1 | 1 | ||
| im | g1v | s | sc | 2 | |||
| im | g1v | sc | погиб | 1 | 1 | 4 | 1 |
| im | g2v | погиб | 2 | ||||
| im | g2v | g2 | g2v | 1 | |||
| im | g2v | g2v | погиб | 2 | |||
| im | g2v | g2v | g2 | 1 | |||
| im | g2v | g2v | g2v | 11 | 4 | 6 | 6 |
| im | g2v | g2v | g3v | 4 | 1 | 3 | 4 |
| im | g2v | g2v | ss | 1 | 1 | ||
| im | g2v | g2v | s | 1 | |||
| im | g2v | g2v | sc | 1 | |||
| im | g2v | g3v | g3v | 3 | 3 | ||
| im | g2v | g3v | ss | 3 | 1 | ||
| im | g2v | g3v | s | 1 | 1 | ||
| im | g2v | g3v | sc | 1 | |||
| im | g2v | ss | погиб | 1 | 1 | 2 | |
| im | g2v | ss | ss | 1 | 2 | 1 | |
| im | g2v | ss | s | 1 | |||
| im | g2v | ss | sc | 1 | 2 | 1 | |
| im | g2v | s | погиб | 2 | |||
| im | g2v | sc | погиб | 2 | 2 | 5 | |
| im | g3v | g3v | g3v | 1 | 1 | ||
| im | g3v | ss | погиб | 1 | |||
| im | g3v | ss | ss | 1 | 3 | 1 | |
| im | ss | ss | s | 1 | |||
| im | ss | s | погиб | 2 | |||
| im | s | погиб | 2 | ||||
| im | s | s | погиб | 1 | |||
| im | s | s | s | 1 | |||
| im | s | s | sc | 2 | |||
| im | s | sc | погиб | 2 | |||
| im | sc | погиб | 1 | 1 | 2 | 6 | |
| Всего | 113 | 109 | 110 | 98 | |||
Таблица 2
Пути онтогенеза парциальных кустов брусники виргинильного состояния
| Год | № ценопопуляции | |||||||
| 1999 | 2000 | 2001 | 2002 | 1 | 2 | 3 | 4 | |
| v | погиб | 1 | 13 | |||||
| v | v | v | v | 1 | ||||
| v | v | g1v | g2v | 1 | 3 | |||
| v | v | g1v | g3v | 1 | 2 | 1 | ||
| v | v | g2v | g2 | 1 | 3 | |||
| v | v | g2v | g2v | 1 | 8 | |||
| v | v | g2v | s | 1 | ||||
| v | v | g3v | g3v | 1 | 4 | |||
| v | v | g3v | ss | 1 | 1 | |||
| v | g1 | g1v | погиб | 1 | 1 | |||
| v | g1 | g1v | g2v | 1 | ||||
| v | g1 | g2 | g2 | 1 | 3 | |||
| v | g1 | g2v | погиб | 1 | ||||
| v | g1 | g2v | g3v | 1 | 1 | |||
| v | g1 | g3v | g3v | 1 | 2 | |||
| v | g1v | g1v | погиб | 3 | ||||
| v | g1v | g1v | g2 | 1 | ||||
| v | g1v | g1v | g2v | 2 | 13 | 7 | ||
| v | g1v | g1v | g3v | 2 | ||||
| v | g1v | g2 | g2 | 2 | 5 | |||
| v | g1v | g2 | g2v | 3 | 3 | |||
| v | g1v | g2 | g3v | 1 | ||||
| v | g1v | g2v | погиб | 1 | 2 | |||
| v | g1v | g2v | g2v | 4 | 1 | 12 | ||
| v | g1v | g2v | g3 | 1 | 2 | |||
| v | g1v | g2v | g3v | 4 | 3 | |||
| v | g1v | g2v | ss | 1 | 2 | |||
| v | g1v | g2v | sc | 1 | ||||
| v | g1v | g3v | погиб | 1 | ||||
| v | g1v | g3v | g3v | 2 | 5 | 5 | ||
| v | g1v | g3v | ss | 1 | 1 | 1 | 3 | |
| v | g1v | ss | погиб | 1 | ||||
| v | g1v | ss | s | 1 | 2 | |||
| v | g1v | ss | sc | 1 | 2 | |||
| v | g1v | s | sc | 1 | 1 | |||
| v | g1v | sc | погиб | 1 | 2 | |||
| v | g2 | g2 | g2 | 1 | ||||
| v | g2 | g2 | g2v | 2 | 1 | |||
| v | g2 | g2v | g2v | 2 | 6 | |||
| v | g2 | g2v | g3v | 1 | 3 | |||
| v | g2 | g2v | ss | 1 | ||||
| v | g2 | g3v | ss | 1 | 1 | 1 | ||
| v | g2 | g3v | s | 1 | 1 | |||
| v | g2 | ss | s | 1 | ||||
| v | g2 | sc | погиб | 1 | 1 | |||
| v | g2v | погиб | 2 | 2 | ||||
| v | g2v | g2 | g2 | 1 | 4 | |||
| v | g2v | g2 | g2v | 1 | 1 | 4 | ||
| v | g2v | g2v | погиб | 2 | 1 | 1 | ||
| v | g2v | g2v | g2 | 2 | 9 | |||
| v | g2v | g2v | g2v | 12 | 1 | 24 | 8 | |
| v | g2v | g2v | g3v | 7 | 2 | 3 | 14 | |
| v | g2v | g2v | ss | 1 | ||||
| v | g2v | g2v | sc | 2 | 1 | 2 | ||
| v | g2v | g3v | g3v | 13 | 1 | 4 | 6 | |
| v | g2v | g3v | ss | 4 | 8 | 1 | ||
| v | g2v | g3v | s | 1 | 2 | 3 | ||
| v | g2v | ss | погиб | 1 | 1 | 1 | ||
| v | g2v | ss | ss | 1 | 1 | 2 | ||
| v | g2v | ss | s | 3 | 1 | 1 | ||
| v | g2v | ss | sc | 1 | 1 | 2 | ||
| v | g2v | s | погиб | 1 | 1 | |||
| v | g2v | s | sc | 1 | 1 | |||
| v | g2v | sc | погиб | 3 | 1 | |||
| v | g3 | sc | погиб | 1 | ||||
| v | g3v | g3v | g3v | 1 | 3 | |||
| v | g3v | g3v | ss | 2 | ||||
| v | g3v | g3v | sc | 1 | ||||
| v | g3v | ss | ss | 2 | 1 | 1 | ||
| v | g3v | ss | s | 1 | ||||
| v | g3v | s | sc | 1 | ||||
| v | g3v | sc | погиб | 1 | ||||
| v | ss | ss | ss | 1 | ||||
| v | ss | ss | s | 1 | ||||
| v | ss | s | погиб | 1 | ||||
| v | ss | s | sc | 1 | 1 | |||
| v | ss | sc | погиб | 2 | 1 | |||
| v | s | погиб | 1 | 1 | ||||
| v | s | s | s | 1 | ||||
| v | s | sc | погиб | 1 | 1 | |||
| v | sc | погиб | 2 | 6 | ||||
| Всего | 76 | 61 | 128 | 135 | ||||
Таблица 3
Пути онтогенеза парциальных кустов брусники молодого генеративного состояния
| Год | № ценопопуляции | |||||||
| 1999 | 2000 | 2001 | 2002 | 1 | 2 | 3 | 4 | |
| g1 | погиб | 4 | ||||||
| g1 | g1 | погиб | 1 | |||||
| g1 | g1 | g2v | g3v | 1 | ||||
| g1 | g1 | g2v | ss | 1 | ||||
| g1 | g1 | g3v | g3v | 2 | 2 | |||
| g1 | g1v | g1v | g2v | 1 | 6 | |||
| g1 | g1v | g1v | ss | 1 | 1 | |||
| g1 | g1v | g2 | g2v | 2 | 4 | |||
| g1 | g1v | g2v | погиб | 1 | ||||
| g1 | g1v | g2v | g2 | 1 | ||||
| g1 | g1v | g2v | g2v | 2 | 7 | 7 | ||
| g1 | g1v | g2v | g3v | 1 | 3 | |||
| g1 | g1v | g2v | ss | 1 | 2 | |||
| g1 | g1v | g2v | sc | 1 | 3 | |||
| g1 | g1v | g3v | погиб | 1 | ||||
| g1 | g1v | g3v | g3v | 4 | 3 | |||
| g1 | g1v | g3v | ss | 1 | ||||
| g1 | g1v | g3v | s | 1 | ||||
| g1 | g1v | g3v | sc | 1 | ||||
| g1 | g1v | ss | ss | 1 | 1 | |||
| g1 | g2 | g2 | g2 | 1 | 1 | 1 | ||
| g1 | g2 | g2 | g2v | 1 | ||||
| g1 | g2 | g2v | g2v | 1 | 1 | 1 | ||
| g1 | g2 | g2v | sc | 1 | ||||
| g1 | g2 | ss | sc | 1 | 1 | |||
| g1 | g2 | sc | погиб | 1 | ||||
| g1 | g2v | g2 | g2 | 2 | ||||
| g1 | g2v | g2v | погиб | 2 | ||||
| g1 | g2v | g2v | g2v | 1 | 7 | 6 | ||
| g1 | g2v | g2v | g3v | 2 | 1 | 11 | ||
| g1 | g2v | g2v | ss | 2 | 1 | 1 | ||
| g1 | g2v | g2v | sc | 1 | 1 | 4 | ||
| g1 | g2v | g3 | g3v | 1 | 2 | |||
| g1 | g2v | g3v | погиб | 1 | ||||
| g1 | g2v | g3v | g3v | 1 | 4 | 3 | ||
| g1 | g2v | g3v | ss | 3 | ||||
| g1 | g2v | g3v | sc | 1 | 1 | 3 | ||
| g1 | g2v | ss | погиб | 1 | 1 | |||
| g1 | g2v | ss | ss | 1 | ||||
| g1 | g2v | ss | s | 1 | 1 | 1 | ||
| g1 | g2v | ss | sc | 1 | 2 | |||
| g1 | g2v | s | sc | 1 | ||||
| g1 | g2v | sc | погиб | 2 | 3 | 1 | ||
| g1 | g3v | погиб | 1 | |||||
| g1 | g3v | g3v | ss | 1 | ||||
| g1 | g3v | ss | погиб | 1 | ||||
| g1 | ss | ss | ss | 1 | 1 | |||
| g1 | ss | sc | погиб | 1 | 1 | |||
| g1 | s | погиб | 1 | |||||
| g1 | s | sc | погиб | 1 | ||||
| g1 | sc | погиб | 1 | 2 | ||||
| g1v | погиб | 1 | 11 | |||||
| g1v | g1 | g1v | g2v | 1 | 4 | |||
| g1v | g1 | g2 | g2v | 1 | ||||
| g1v | g1 | g2v | погиб | 1 | ||||
| g1v | g1 | g2v | g2 | 3 | ||||
| g1v | g1 | g2v | g2v | 2 | 8 | |||
| g1v | g1 | g3v | sc | 1 | 1 | |||
| g1v | g1 | ss | ss | 1 | ||||
| g1v | g1v | g1v | g1v | 1 | 7 | |||
| g1v | g1v | g1v | g2v | 1 | 9 | |||
| g1v | g1v | g2 | g2 | 1 | 1 | |||
| g1v | g1v | g2v | g2 | 2 | ||||
| g1v | g1v | g2v | g2v | 2 | 9 | 12 | ||
| g1v | g1v | g2v | g3 | 1 | ||||
| g1v | g1v | g2v | g3v | 2 | 2 | 4 | ||
| g1v | g1v | g2v | ss | 2 | 2 | |||
| g1v | g1v | g3v | ss | 1 | 4 | |||
| g1v | g1v | g3v | sc | 1 | 3 | |||
| g1v | g1v | ss | ss | 1 | ||||
| g1v | g1v | ss | s | 1 | ||||
| g1v | g1v | ss | sc | 2 | 1 | |||
| g1v | g1v | sc | погиб | 1 | 1 | 1 | ||
| g1v | g2 | g2v | погиб | 1 | ||||
| g1v | g2 | g2v | g2v | 2 | 4 | 3 | ||
| g1v | g2 | g3v | g3v | 2 | 2 | |||
| g1v | g2 | g3v | sc | 2 | ||||
| g1v | g2 | ss | sc | 1 | ||||
| g1v | g2 | s | sc | 1 | 3 | |||
| g1v | g2 | sc | погиб | 1 | ||||
| g1v | g2v | погиб | 1 | |||||
| g1v | g2v | g2 | g2v | 1 | 1 | 5 | ||
| g1v | g2v | g2v | g2 | 1 | ||||
| g1v | g2v | g2v | g2v | 29 | 5 | 24 | 15 | |
| g1v | g2v | g2v | g3v | 18 | 2 | 7 | 5 | |
| g1v | g2v | g2v | ss | 1 | 1 | |||
| g1v | g2v | g3 | g3v | 1 | ||||
| g1v | g2v | g3v | погиб | 3 | ||||
| g1v | g2v | g3v | g3v | 18 | 5 | 4 | ||
| g1v | g2v | g3v | ss | 4 | 1 | 4 | 3 | |
| g1v | g2v | g3v | s | 1 | ||||
| g1v | g2v | g3v | sc | 1 | 3 | 1 | 1 | |
| g1v | g2v | ss | погиб | 1 | 1 | |||
| g1v | g2v | ss | ss | 2 | 1 | 2 | ||
| g1v | g2v | ss | s | 3 | ||||
| g1v | g2v | ss | sc | 1 | 1 | 2 | 4 | |
| g1v | g2v | s | погиб | 1 | ||||
| g1v | g2v | s | s | 1 | ||||
| g1v | g2v | s | sc | 1 | 1 | 1 | 1 | |
| g1v | g2v | sc | погиб | 3 | 3 | |||
| g1v | g3 | g3v | ss | 1 | 1 | |||
| g1v | g3 | g3v | sc | 1 | ||||
| g1v | g3 | s | погиб | 1 | ||||
| g1v | g3 | sc | погиб | 2 | ||||
| g1v | g3v | g3v | g3v | 5 | 1 | 3 | ||
| g1v | g3v | g3v | ss | 1 | ||||
| g1v | g3v | g3v | s | 1 | ||||
| g1v | g3v | g3v | sc | 1 | ||||
| g1v | g3v | ss | погиб | 1 | ||||
| g1v | g3v | ss | ss | 3 | ||||
| g1v | g3v | ss | sc | 2 | 1 | |||
| g1v | g3v | s | погиб | 1 | 1 | 2 | ||
| g1v | g3v | s | sc | 1 | 1 | 1 | 2 | |
| g1v | g3v | sc | погиб | 3 | 1 | 2 | ||
| g1v | ss | ss | ss | 1 | 1 | |||
| g1v | ss | ss | sc | 2 | 2 | |||
| g1v | ss | s | погиб | 1 | ||||
| g1v | ss | s | sc | 1 | 1 | |||
| g1v | ss | sc | погиб | 3 | 2 | 1 | 1 | |
| g1v | s | погиб | 6 | |||||
| g1v | s | s | s | 1 | ||||
| g1v | s | sc | погиб | 9 | 1 | 1 | ||
| g1v | sc | погиб | 1 | 4 | 4 | 5 | ||
| g1v | sc | ss | погиб | 1 | ||||
| g1v | sc | ss | sc | 1 | ||||
| Всего | 102 | 118 | 159 | 208 | ||||
Таблица 4
Пути онтогенеза парциальных кустов брусники средневозрастного генеративного состояния
| Год | № ценопопуляции | |||||||
| 1999 | 2000 | 2001 | 2002 | 1 | 2 | 3 | 4 | |
| g2 | погиб | 3 | 1 | |||||
| g2 | g2 | погиб | 1 | |||||
| g2 | g2 | g2 | g2v | 1 | ||||
| g2 | g2 | g2 | sc | 1 | ||||
| g2 | g2 | g2v | g2v | 1 | ||||
| g2 | g2 | g2v | sc | 1 | ||||
| g2 | g2 | g3 | g3v | 1 | ||||
| g2 | g2 | g3v | погиб | 1 | ||||
| g2 | g2 | g3v | g3v | 2 | 1 | |||
| g2 | g2 | g3v | sc | 2 | ||||
| g2 | g2 | ss | sc | 1 | ||||
| g2 | g2 | s | sc | 1 | ||||
| g2 | g2v | погиб | 6 | |||||
| g2 | g2v | g2 | g2 | 1 | ||||
| g2 | g2v | g2 | g2v | 1 | 4 | |||
| g2 | g2v | g2 | g3v | 1 | ||||
| g2 | g2v | g2v | погиб | 1 | ||||
| g2 | g2v | g2v | g2 | 1 | 1 | |||
| g2 | g2v | g2v | g2v | 1 | 26 | 7 | ||
| g2 | g2v | g2v | g3v | 4 | 5 | 1 | ||
| g2 | g2v | g2v | ss | 1 | ||||
| g2 | g2v | g3v | погиб | 3 | 4 | 1 | ||
| g2 | g2v | g3v | g3v | 4 | 3 | |||
| g2 | g2v | g3v | ss | 3 | ||||
| g2 | g2v | g3v | s | 1 | ||||
| g2 | g2v | g3v | sc | 2 | 1 | 1 | ||
| g2 | g2v | ss | погиб | 2 | ||||
| g2 | g2v | ss | s | 1 | 1 | 1 | ||
| g2 | g2v | ss | sc | 1 | 1 | |||
| g2 | g2v | s | погиб | 1 | 1 | |||
| g2 | g2v | s | sc | 3 | 1 | |||
| g2 | g2v | sc | погиб | 4 | 2 | |||
| g2 | g2v | sc | ss | 1 | ||||
| g2 | g3 | g3v | g3v | 1 | 1 | |||
| g2 | g3 | g3v | sc | 1 | 1 | |||
| g2 | g3v | погиб | 3 | |||||
| g2 | g3v | g3v | погиб | 1 | 1 | |||
| g2 | g3v | g3v | g3v | 1 | 1 | 4 | ||
| g2 | g3v | g3v | g3 | 3 | ||||
| g2 | g3v | g3v | s | 1 | ||||
| g2 | g3v | g3v | sc | 1 | ||||
| g2 | g3v | ss | погиб | 1 | ||||
| g2 | g3v | ss | ss | 1 | ||||
| g2 | g3v | ss | s | 1 | ||||
| g2 | g3v | ss | sc | 1 | 2 | |||
| g2 | g3v | s | погиб | 1 | ||||
| g2 | g3v | s | sc | 1 | ||||
| g2 | g3v | sc | погиб | 1 | 2 | 1 | ||
| g2 | ss | ss | ss | 1 | ||||
| g2 | ss | s | погиб | 1 | ||||
| g2 | ss | s | s | 1 | ||||
| g2 | ss | sc | погиб | 1 | 1 | 4 | ||
| g2 | s | s | sc | 1 | 1 | |||
| g2 | s | sc | погиб | 1 | 3 | |||
| g2 | sc | погиб | 2 | 6 | ||||
| g2v | погиб | 1 | 7 | 3 | ||||
| g2v | g2 | g2 | g2 | 3 | ||||
| g2v | g2 | g2 | g2v | 2 | ||||
| g2v | g2 | g2v | g2v | 1 | 7 | |||
| g2v | g2 | g2v | g3v | 1 | ||||
| g2v | g2 | g2v | sc | 2 | 2 | |||
| g2v | g2 | g3v | погиб | 1 | ||||
| g2v | g2 | g3v | g3v | 2 | ||||
| g2v | g2 | g3v | ss | 1 | ||||
| g2v | g2 | ss | погиб | 1 | ||||
| g2v | g2 | ss | s | 1 | ||||
| g2v | g2 | ss | sc | 1 | 1 | |||
| g2v | g2 | sc | погиб | 4 | ||||
| g2v | g2v | погиб | 2 | 1 | 1 | |||
| g2v | g2v | g2 | g2 | 1 | ||||
| g2v | g2v | g2 | g2v | 9 | ||||
| g2v | g2v | g2v | погиб | 2 | 1 | |||
| g2v | g2v | g2v | g2 | 2 | ||||
| g2v | g2v | g2v | g2v | 7 | 44 | 12 | ||
| g2v | g2v | g2v | g3 | 1 | ||||
| g2v | g2v | g2v | g3v | 5 | 1 | 3 | 11 | |
| g2v | g2v | g2v | ss | 1 | ||||
| g2v | g2v | g2v | s | 1 | ||||
| g2v | g2v | g2v | sc | 2 | 1 | |||
| g2v | g2v | g3v | погиб | 2 | 4 | |||
| g2v | g2v | g3v | g3v | 3 | 12 | 5 | ||
| g2v | g2v | g3v | ss | 3 | 4 | 3 | ||
| g2v | g2v | g3v | s | 1 | 1 | |||
| g2v | g2v | g3v | sc | 1 | 7 | 1 | ||
| g2v | g2v | ss | погиб | 1 | 1 | 2 | 1 | |
| g2v | g2v | ss | ss | 2 | 2 | |||
| g2v | g2v | ss | s | 4 | ||||
| g2v | g2v | ss | sc | 1 | 6 | 2 | ||
| g2v | g2v | s | sc | 1 | 2 | |||
| g2v | g2v | sc | погиб | 3 | 6 | 3 | ||
| g2v | g3 | погиб | 3 | |||||
| g2v | g3 | g3v | погиб | 1 | 1 | |||
| g2v | g3 | g3v | g3v | 1 | ||||
| g2v | g3 | g3v | sc | 1 | ||||
| g2v | g3 | ss | s | 1 | ||||
| g2v | g3 | ss | sc | 1 | ||||
| g2v | g3 | sc | погиб | 1 | 1 | |||
| g2v | g3v | погиб | 1 | 1 | ||||
| g2v | g3v | g3v | погиб | 1 | ||||
| g2v | g3v | g3v | g3 | 1 | ||||
| g2v | g3v | g3v | g3v | 2 | 1 | 3 | 7 | |
| g2v | g3v | g3v | ss | 3 | 1 | 4 | ||
| g2v | g3v | g3v | sc | 2 | ||||
| g2v | g3v | ss | погиб | 1 | 1 | 3 | ||
| g2v | g3v | ss | ss | 1 | 1 | 1 | 1 | |
| g2v | g3v | ss | s | 1 | ||||
| g2v | g3v | ss | sc | 2 | 1 | |||
| g2v | g3v | s | погиб | 1 | 1 | |||
| g2v | g3v | s | sc | 2 | 2 | |||
| g2v | g3v | sc | погиб | 3 | 4 | |||
| g2v | ss | погиб | 2 | 7 | ||||
| g2v | ss | ss | погиб | 1 | 1 | |||
| g2v | ss | ss | s | 1 | ||||
| g2v | ss | s | погиб | 1 | 3 | |||
| g2v | ss | s | s | 1 | 1 | |||
| g2v | ss | s | sc | 1 | 2 | |||
| g2v | ss | sc | погиб | 2 | 3 | |||
| g2v | s | погиб | 1 | 3 | ||||
| g2v | s | s | sc | 1 | ||||
| g2v | s | sc | погиб | 2 | 2 | 2 | ||
| g2v | sc | погиб | 10 | 2 | 16 | |||
| Всего | 38 | 82 | 247 | 172 | ||||
Таблица 5
Пути онтогенеза парциальных кустов брусники старого генеративного состояния
| Год | № ценопопуляции | |||||||
| 1999 | 2000 | 2001 | 2002 | 1 | 2 | 3 | 4 | |
| g3 | g3 | погиб | 1 | 2 | ||||
| g3 | g3v | погиб | 1 | 1 | ||||
| g3 | g3v | g3v | g3 | 1 | ||||
| g3 | g3v | g3v | g3v | 2 | 4 | |||
| g3 | g3v | g3v | ss | 1 | ||||
| g3 | g3v | ss | погиб | 2 | ||||
| g3 | g3v | ss | sc | 2 | 1 | |||
| g3 | g3v | sc | погиб | 1 | ||||
| g3 | ss | погиб | 1 | |||||
| g3 | ss | ss | погиб | 1 | ||||
| g3 | ss | ss | ss | 1 | 2 | |||
| g3 | ss | ss | sc | 2 | ||||
| g3 | s | погиб | 1 | 1 | ||||
| g3 | s | s | погиб | 1 | ||||
| g3 | s | sc | погиб | 1 | ||||
| g3 | sc | погиб | 6 | |||||
| g3v | погиб | 1 | 13 | 3 | ||||
| g3v | g3 | g3 | g3v | 1 | ||||
| g3v | g3 | g3v | погиб | 1 | ||||
| g3v | g3 | g3v | g3v | 7 | 1 | |||
| g3v | g3 | g3v | s | 1 | ||||
| g3v | g3 | g3v | sc | 1 | ||||
| g3v | g3 | ss | ss | 1 | ||||
| g3v | g3 | ss | s | 1 | ||||
| g3v | g3 | sc | погиб | 1 | 1 | 1 | ||
| g3v | g3v | погиб | 2 | 2 | ||||
| g3v | g3v | g3 | ss | 1 | ||||
| g3v | g3v | g3v | погиб | 2 | 1 | |||
| g3v | g3v | g3v | g3v | 6 | 1 | 9 | 12 | |
| g3v | g3v | g3v | g3 | 1 | 3 | |||
| g3v | g3v | g3v | sc | 1 | 3 | |||
| g3v | g3v | ss | погиб | 2 | 1 | 2 | ||
| g3v | g3v | ss | ss | 1 | 2 | 1 | ||
| g3v | g3v | ss | s | 1 | 1 | |||
| g3v | g3v | ss | sc | 1 | 2 | 2 | 1 | |
| g3v | g3v | s | погиб | 1 | 1 | 1 | ||
| g3v | g3v | s | sc | 1 | 1 | 1 | ||
| g3v | g3v | sc | погиб | 2 | 2 | 3 | 7 | |
| g3v | ss | ss | ss | 1 | 1 | 1 | 1 | |
| g3v | ss | ss | s | 1 | ||||
| g3v | ss | ss | sc | 1 | 1 | |||
| g3v | ss | s | погиб | 2 | 1 | |||
| g3v | ss | s | sc | 3 | 1 | |||
| g3v | ss | sc | погиб | 2 | 3 | |||
| g3v | s | погиб | 1 | 3 | ||||
| g3v | s | ss | sc | 1 | ||||
| g3v | s | s | sc | 1 | ||||
| g3v | s | sc | погиб | 5 | 3 | 3 | ||
| g3v | sc | погиб | 1 | 6 | 17 | |||
| g3v | sc | ss | погиб | 1 | ||||
| g3v | sc | sc | погиб | 1 | ||||
| Всего | 19 | 46 | 61 | 87 | ||||
Таблица 6
Пути онтогенеза парциальных кустов брусники субсенильного состояния
| Год | № ценопопуляции | |||||||
| 1999 | 2000 | 2001 | 2002 | 1 | 2 | 3 | 4 | |
| ss | погиб | 2 | 5 | 5 | ||||
| ss | ss | ss | погиб | 1 | ||||
| ss | ss | ss | ss | 1 | 1 | 2 | ||
| ss | ss | ss | sc | 1 | 1 | |||
| ss | ss | s | погиб | 1 | 2 | 3 | ||
| ss | ss | s | s | 1 | ||||
| ss | ss | s | sc | 2 | 1 | |||
| ss | ss | sc | погиб | 1 | 3 | 5 | ||
| ss | s | погиб | 2 | |||||
| ss | s | sc | погиб | 2 | 2 | 4 | ||
| ss | sc | погиб | 1 | 5 | 12 | |||
| Всего | 5 | 13 | 11 | 37 | ||||
Таблица 7
Пути онтогенеза парциальных кустов брусники сенильного состояния
| Год | № ценопопуляции | |||||||
| 1999 | 2000 | 2001 | 2002 | 1 | 2 | 3 | 4 | |
| s | погиб | 2 | 1 | |||||
| s | s | погиб | 2 | |||||
| s | s | sc | погиб | 1 | ||||
| s | s | sc | 2 | |||||
| s | sc | погиб | 2 | 16 | ||||
| Всего | 0 | 4 | 1 | 21 | ||||
Таблица 8
Пути онтогенеза парциальных кустов брусники отмирающего состояния
| Год | № ценопопуляции | ||||||
| 1999 | 2000 | 2001 | 2002 | 1 | 2 | 3 | 4 |
| sc | погиб | 3 | 16 | 4 | |||
| sc | sc | погиб | 12 | ||||
| sc | ss | sc | погиб | 2 | |||
| Всего | 3 | 18 | 0 | 16 | |||